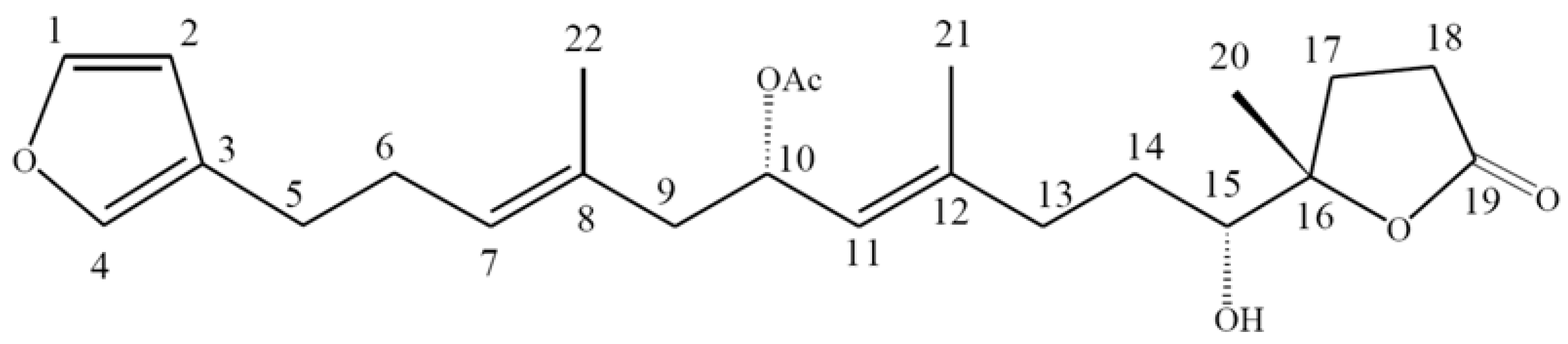
10-Acetylirciformonin B, A Sponge Furanoterpenoid, Induces DNA Damage and Apoptosis in Leukemia Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Results and Discussion
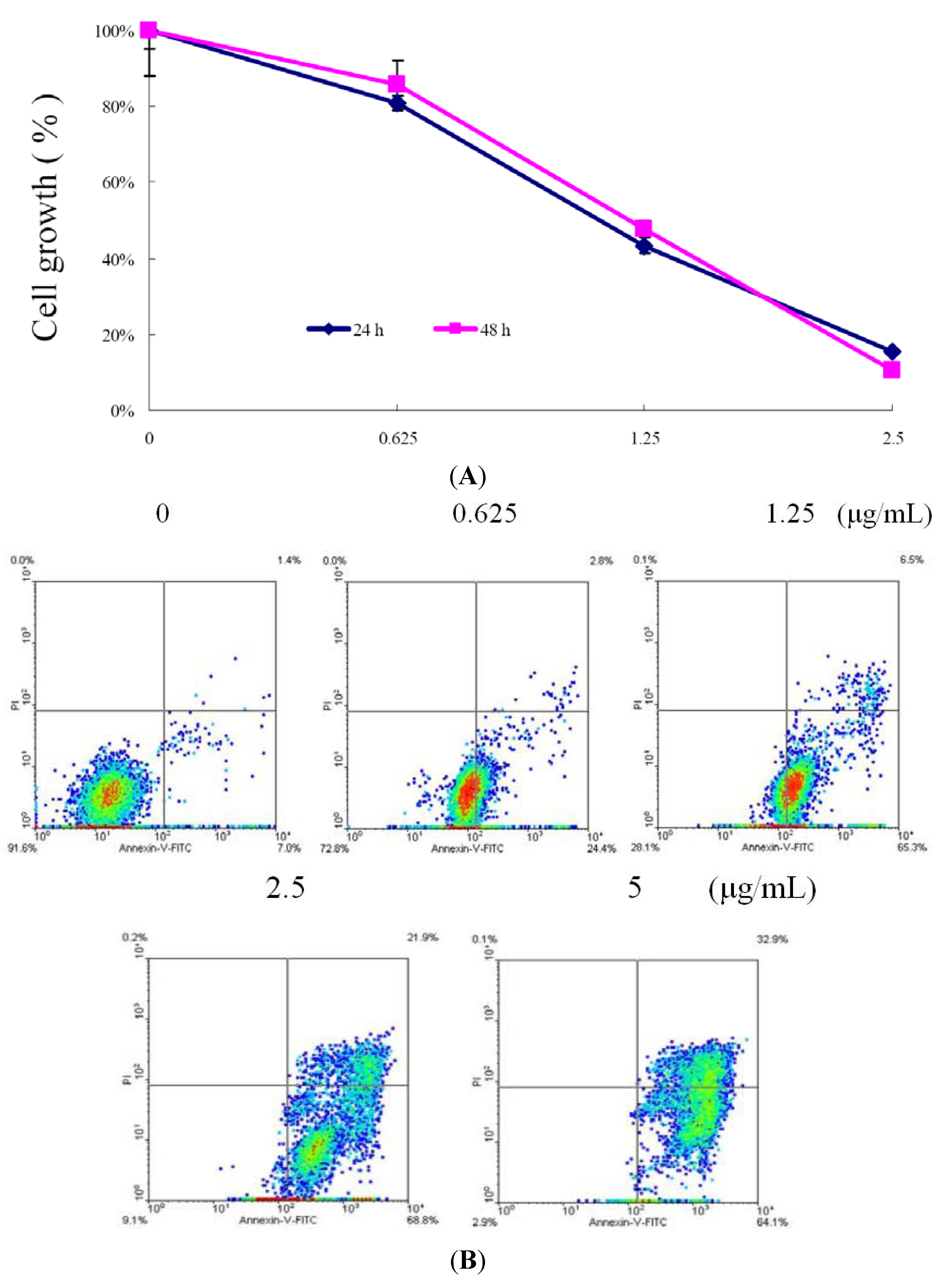
2.1. 10-Acetylirciformonin B is A Potential Inhibitor of Cell Growth and Inducer of Apoptosis in Leukemia HL 60 Cells

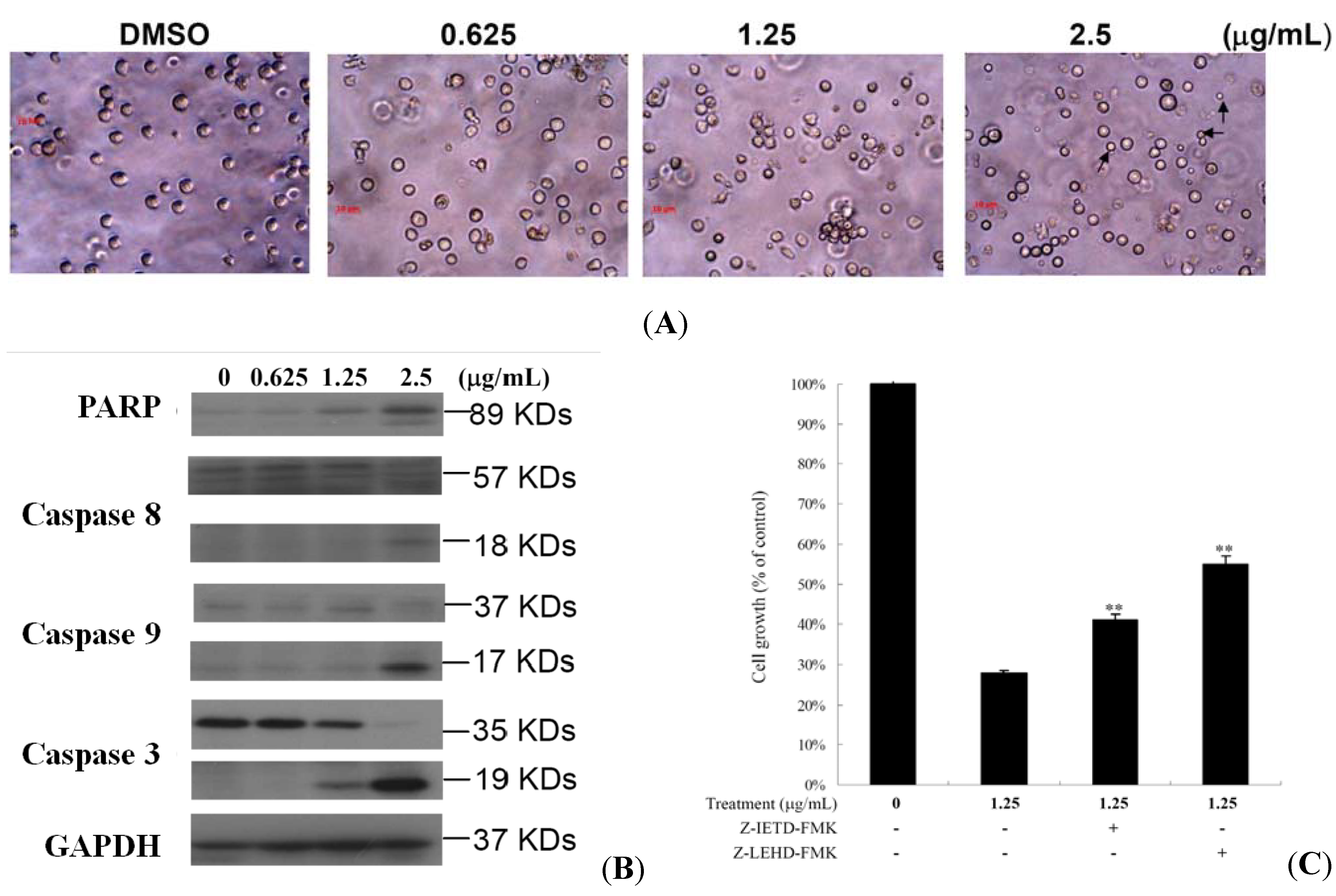
2.2. 10-Acetylirciformonin B Treatment Induced HL 60 Cells DNA Double-Strand Breaks

2.3. 10-Acetylirciformonin B Induced HL 60 Cells Apoptosis through Caspase-Dependent Pathway

3. Experimental
3.1. Bioassays Materials
3.2. Preparation of 10-Acetylirciformonin B
3.3. MTT Antiproliferative Assay
3.4. Annexin V/PI Apoptosis Assay
3.5. Neutral Comet Assay for Detection of DNA Double-strand Breaks (DSBs)
3.6. Western Blotting Analysis
3.7. Statistics
4. Conclusions
Acknowledgements
- Samples Availability: Sample of 10-acetylirciformonin B is available from the authors.
References
- D’Autréaux, B.; Toledano, M.B. ROS as signalling molecules: mechanisms that generate specificity in ROS homeostasis. Nat. Rev. Mol. Cell Biol. 2007, 8, 813–824. [Google Scholar] [CrossRef]
- Fruehauf, J.P.; Meyskens, F.L., Jr. Reactive oxygen species: A breath of life or death? Clin. Cancer Res. 2007, 13, 789–794. [Google Scholar] [CrossRef]
- Chen, H.M.; Wu, Y.C.; Chia, Y.C.; Chang, F.R.; Hsu, H.K.; Hsieh, Y.C.; Chen, C.C.; Yuan, S.S. Gallic acid, A major component of Toona sinensis leaf extracts, Contains a ROS-mediated anti-cancer activity in human prostate cancer cells. Cancer Lett. 2009, 286, 161–171. [Google Scholar] [CrossRef]
- Havsteen, B. Flavonoids, A class of natural products of high pharmacological potency. Biochem. Pharmacol. 1983, 32, 1141–1148. [Google Scholar]
- Mroz, R.M.; Schins, R.P.F.; Li, H.; Jimenez, L.A.; Drost, E.M.; Holownia, A.; MacNee, W.; Donaldson, K. Nanoparticle-driven DNA damage mimics irradiation-related carcinogenesis pathways. Eur. Respir. J. 2008, 31, 241–251. [Google Scholar] [CrossRef]
- Brem, R.; Montaner, B.; Reelfs, O.; Karran, P. DNA breakage and cell cycle checkpoint abrogation induced by a therapeutic thiopurine and UVA radiation. Oncogene 2010, 29, 3953–3963. [Google Scholar] [CrossRef]
- Ha, L.; Ceryak, S.; Patierno, S.R. Generation of S phase-dependent DNA double-strand breaks by Cr(VI) exposure: involvement of ATM in Cr(VI) induction of gamma-H2AX. Carcinogenesis 2004, 25, 2265–2274. [Google Scholar] [CrossRef]
- Powell, S.N.; Bindra, R.S. Targeting the DNA damage response for cancer therapy. DNA Repair 2009, 8, 1153–1165. [Google Scholar] [CrossRef]
- Chen, H.M.; Chang, F.R.; Hsieh, Y.C.; Cheng, Y.J.; Hseih, K.C.; Tsai, L.M.; Lin, A.S.; Wu, Y.C.; Yuan, S.S. A novel synthetic protoapigenone analogue, WYC02–9, Induces DNA damage and apoptosis in DU145 prostate cancer cells through generation of reactive oxygen species. Free Radic. Biol. Med. 2011, 50, 1151–1162. [Google Scholar] [CrossRef]
- Kastan, M.B.; Lim, D.S. The many substrates and functions of ATM. Nat. Rev. Mol. Cell Biol. 2000, 1, 179–186. [Google Scholar] [CrossRef]
- Bakkenist, C.J.; Kastan, M.B. DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature 2003, 421, 499–506. [Google Scholar] [CrossRef]
- Abraham, R.T.; Tibbetts, R.S. Guiding ATM to broken DNA. Science 2005, 308, 510–511. [Google Scholar]
- Bulavin, D.V.; Higashimoto, Y.; Demidenko, Z.N.; Meek, S.; Graves, P.; Phillips, C.; Zhao, H.; Moody, S.A.; Apella, E.; Piwnica-Worms, H.; et al. Dual phosphorylation controls Cdc25 phosphatases and mitotic entry. Nat. Cell Biol. 2003, 5, 545–551. [Google Scholar] [CrossRef]
- Jang, E.R.; Choi, J.D.; Park, M.A.; Jeong, G.; Cho, H.; Lee, J.S. ATM modulates transcription in response to histone deacetylase inhibition as part of its DNA damage response. Exp. Mol. Med. 2010, 42, 195–204. [Google Scholar] [CrossRef]
- Skropeta, D.; Pastro, N.; Zivanovic, A. Kinase inhibitors from marine sponges. Mar. Drugs 2011, 9, 2131–2154. [Google Scholar] [CrossRef]
- Koopmans, M.; Martens, D.; Wijffels, R.H. Towards commercial production of sponge medicines. Mar. Drugs 2009, 7, 787–802. [Google Scholar] [CrossRef]
- Mohapatra, B.R.; Bapuji, M.; Sree, A. Antifungal efficacy of bacteria isolated from marine sedentary organisms. Folia Microbiol. 2002, 47, 51–55. [Google Scholar] [CrossRef]
- Li, D.; Xu, Y.; Shao, C.L.; Yang, R.Y.; Zheng, C.J.; Chen, Y.Y.; Fu, X.M.; Qian, P.Y.; She, Z.G.; de Voogd, N.J.; et al. Antibacterial Bisabolane-Type Sesquiterpenoids from the Sponge-Derived Fungus Aspergillus sp. Mar. Drugs 2012, 10, 234–241. [Google Scholar] [CrossRef]
- Ohno, O.; Suenaga, K.; Uemura, D. Secondary metabolites with new medicinal functions from marine organisms. Adv. Food Nutr. Res. 2012, 65, 185–193. [Google Scholar] [CrossRef]
- Su, J.H.; Tseng, S.W.; Lu, M.C.; Liu, L.L.; Chou, Y.; Sung, P.J. Cytotoxic C21 and C22 terpenoid-derived metabolites from the sponge Ircinia sp. J. Nat. Prod. 2011, 74, 2005–2009. [Google Scholar] [CrossRef]
- Yang, H.L.; Hseu, Y.C.; Chen, J.Y.; Yech, Y.J.; Lu, F.J.; Wang, H.H.; Lin, P.S.; Wamg, B.C. Antrodia camphorata in submerged culture protects low density lipoproteins against oxidative modification. Am. J. Chin. Med. 2006, 34, 217–231. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Su, J.-H.; Chang, W.-B.; Chen, H.-M.; El-Shazly, M.; Du, Y.-C.; Kung, T.-H.; Chen, Y.-C.; Sung, P.-J.; Ho, Y.-S.; Kuo, F.-W.; et al. 10-Acetylirciformonin B, A Sponge Furanoterpenoid, Induces DNA Damage and Apoptosis in Leukemia Cells. Molecules 2012, 17, 11839-11848. https://doi.org/10.3390/molecules171011839
Su J-H, Chang W-B, Chen H-M, El-Shazly M, Du Y-C, Kung T-H, Chen Y-C, Sung P-J, Ho Y-S, Kuo F-W, et al. 10-Acetylirciformonin B, A Sponge Furanoterpenoid, Induces DNA Damage and Apoptosis in Leukemia Cells. Molecules. 2012; 17(10):11839-11848. https://doi.org/10.3390/molecules171011839
Chicago/Turabian StyleSu, Jui-Hsin, Wen-Been Chang, Huei-Mei Chen, Mohamed El-Shazly, Ying-Chi Du, Ting-Hsuan Kung, Yu-Cheng Chen, Ping-Jyun Sung, Yuan-Shing Ho, Fu-Wen Kuo, and et al. 2012. "10-Acetylirciformonin B, A Sponge Furanoterpenoid, Induces DNA Damage and Apoptosis in Leukemia Cells" Molecules 17, no. 10: 11839-11848. https://doi.org/10.3390/molecules171011839