Therapeutic Applications of Nucleic Acids and Their Analogues in Toll-like Receptor Signaling

Abstract
:1. Introduction
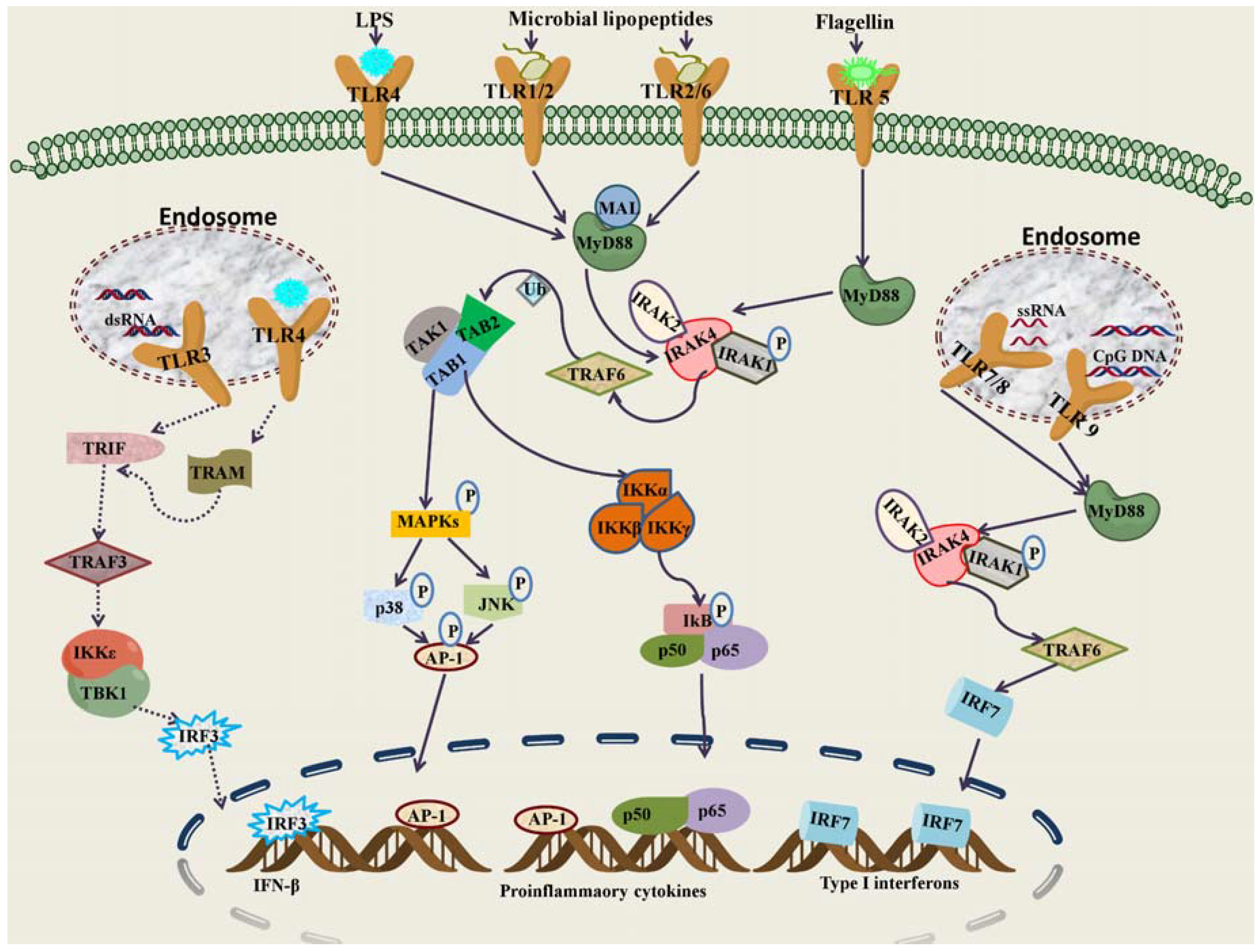
2. Nucleic Acid Sensing Toll-Like Receptors and Their Signaling Pathways

3. Cellular Localization and Regulation of Nucleic Acid Sensing TLRs
4. Nucleic Acid Sensing TLR Agonists and Their Therapeutic Applications
4.1. TLR3

{kind=link}
{kind=link}
| Compd. | Phase | Status | Indications | Notes | ClinicalTrials.gov [83] |
|---|---|---|---|---|---|
| Ampligen® | I | Completed | HIV infections | Single agent | NCT00000735 |
| I | Completed | HIV infections | Single agent | NCT00000713 | |
| III | Recruiting | Chronic fatigue syndrome | Single agent | NCT00215813 | |
| Not yet recruiting | Colorectal cancer | Combined with IFN-α and celecoxib | NCT01545141 | ||
| I-II | Active but not yet recruiting | Ovarian, fallopian tube or primary peritoneal cancer | Combined with tumor cell lysate and multiple adjuvants | NCT01312389 | |
| I-II | Recruiting | Breast cancer | GM-CSF as a combined adjuvant strategy with HER2 vaccination | NCT01355393 | |
| Hiltonol® | I | Completed | Respiratory infections | Single agent | NCT00646152 |
| II | Recruiting | Low grade glioma | Single agent | NCT01188096 | |
| II | Recruiting | Colorectal cancer | Combined with MUC1peptide vaccine | NCT00773097 | |
| I-II | Recruiting | Melanoma | Combined with NY-ESO-1based vaccine | NCT01079741 | |
| I | Active but not yet recruiting | Prostate cancer | Combined with MUC1 | NCT00374049 | |
| 0 | Recruiting | Breast cancer | Combined with MUC1 peptide vaccine | NCT00986609 | |
| 0 | Recruiting | Adenocarcinoma | Vaccination with DCs | NCT01677962 | |
| I | Not yet recruiting | Melanoma | Combined with multipeptide vaccine | NCT01585350 | |
| II | Recruiting | Advanced myeloma | Combined with multipeptide vaccine and lenalidomide | NCT01245673 |
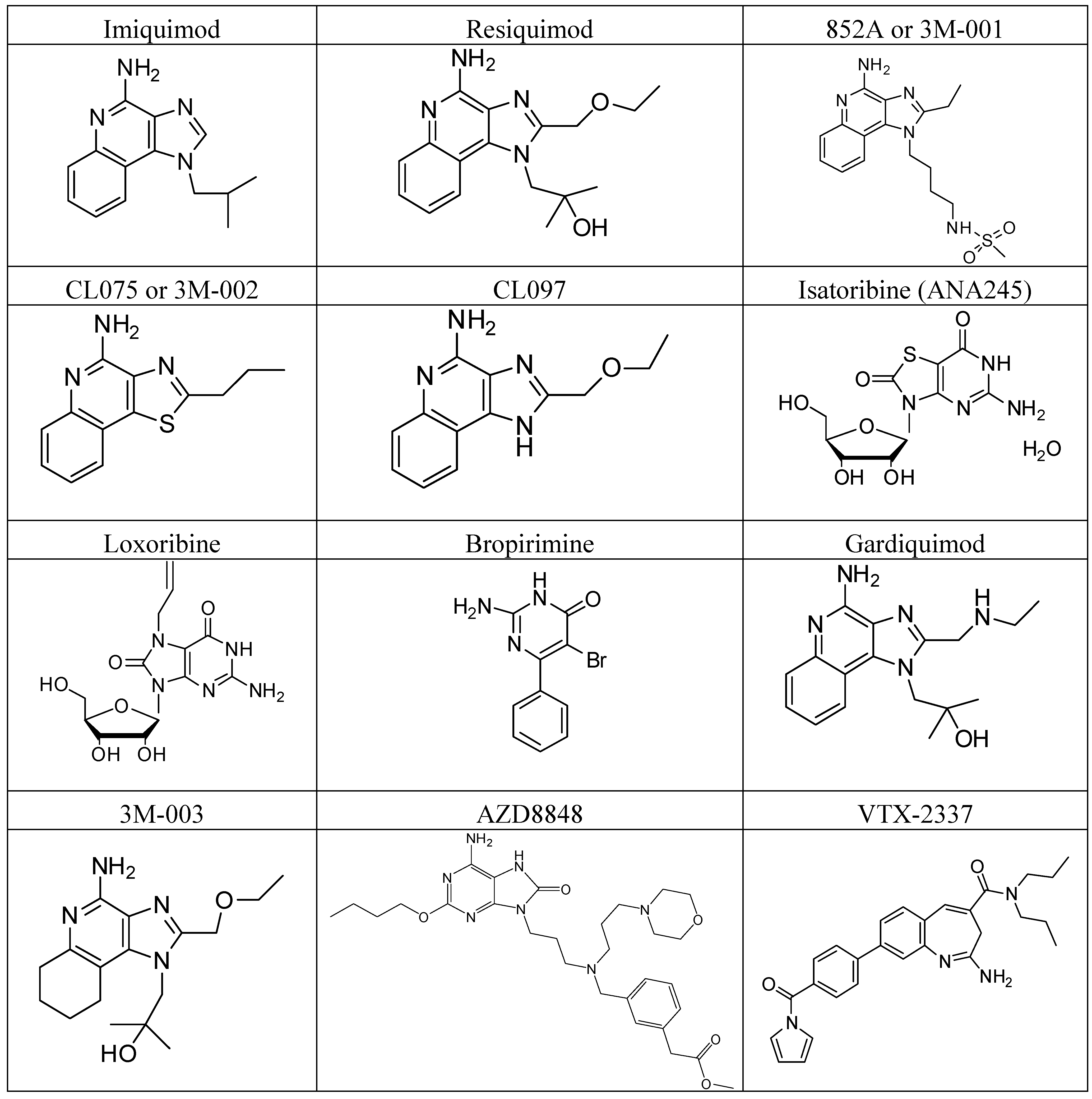
4.2. TLR7/TLR8

| Compd. | Phase | Status | Indications | Notes | ClinicalTrials.gov [83] | |||
|---|---|---|---|---|---|---|---|---|
| Imiquimod | IV | Recruiting | Basal cell carcinoma | Single agent | NCT00803907 | |||
| Not available | Recruiting | Melanoma | Combined with multipeptide vaccine | NCT01264731 | ||||
| II | Recruiting | Breast cancer | Together with Abraxane | NCT00821964 | ||||
| I | Recruiting | Glioma | Combined with lysate based vaccine | NCT01400672 | ||||
| I-II | Recruiting | Metastatic breast cancer | Together with radiotherapy | NCT01421017 | ||||
| IV | Recruiting | Plantar warts | NCT01059110 | |||||
| Not available | Recruiting | Psoriasis | Together with UV radiation | NCT00470392 | ||||
| Resiquimod | II | Recruiting | Actinic keratosis | Single agent | NCT01583816 | |||
| I-II | Recruiting | T cell lymphoma | Single agent | NCT01676831 | ||||
| I-II | Recruiting | T cell lymphoma | Single agent | NCT01497795 | ||||
| II | Recruiting | Melanoma | Combined with gp 100 and MAGE-3 based vaccine | NCT00960752 | ||||
| I-II | Active but not recruiting | Advanced malignancies | Combined with CDX-1401 | NCT00948961 | ||||
| VTX-2337 | I | Recruiting | HNSCC | Combined with cetuximab | NCT01334177 | |||
| I | Recruiting | Reproductive tract cancer | Combined with liposomal doxorubicin | NCT01294293 | ||||
| II | Not yet recruiting | Reproductive tract cancer | Combined with liposomal doxorubicin | NCT01666444 | ||||
| I | Completed | Advanced solid tumor | Single agent | NCT00688415 | ||||
| 852A | II | Completed | Melanoma | Single agent | NCT00189332 | |||
| I | Completed | Neoplasms | Single agent | NCT00095160 | ||||
| Not available | Terminated | Barrett esophagus | Single agent | NCT00386594 | ||||
| II | Terminated | Hematologic malignancies | Single agent | NCT00276159 | ||||
| ANA773 | I | Completed | HCV infection | Single agent | NCT01211626 | |||
| AZD8848/DS P-3025 | II | Completed | Allergic asthma | Single agent | NCT00999466 | |||
| I | Completed | Allergic rhinitis | Single agent | NCT00688779 | ||||
| I | Recruiting | Healthy | Single agent | NCT01560234 | ||||
| I | Completed | Allergic rhinitis | Single agent | NCT00925678 | ||||
4.3. TLR9
| Compd. | Phase | Status | Indications | Notes | ClinicalTrials.gov [83] |
|---|---|---|---|---|---|
| IMO-2055 | I | Terminated | Colorectal cancer | Combined with Erbitux and FOLFIRI | NCT00719199 |
| I | Terminated | Head and neck carcinoma | Combined with Erbitux, 5-FU, and cisplatin | NCT01360827 | |
| II | Active but not recruiting | Head and neck carcinoma | Combined with Erbitux | NCT01040832 | |
| II | Completed | Renal cell carcinoma | Single agent | NCT00729053 | |
| I | Completed | Non-small cell lung cancer | Single agent | NCT00633529 | |
| IMO-2125 | I | Completed | Hepatitis c | Combined with ribavirin | NCT00990938 |
| I | Completed | Hepatitis c | Single agent | NCT00728936 | |
| MGN-1703 | II | Active but not recruiting | Colorectal carcinoma | Single agent | NCT01208194 |
| HEPLISAVTM | III | Active but not recruiting | Chronic kidney disease | Single agent | NCT00985426 |
| III | Recruiting | Renal disease | Single agent | NCT01195246 | |
| SD-101 | I | Completed | Hepatitis c | Combined with ribavirin | NCT00823862 |
| CpG 7909 | II | Recruiting | Breast cancer | Combined with trastuzumab | NCT00824733 |
| I-II | Completed | Metastatic breast carcinoma | Combined with Herceptin | NCT00031278 | |
| I-II | Terminated | Malignant melanoma | Combined with MAGE-3.A1 peptide | NCT00145145 | |
| I | Suspended | Septicemia | Combined with J5-OMP vaccine | NCT01164514 | |
| GNKG168 | I | Recruiting | Leukemia | Single agent | NCT01035216 |
5. Nucleic Acid Sensing TLR Antagonists and Their Therapeutic Applications
TLR7, TLR8, and TLR9
6. Nucleic Acid Sensing Toll-Like Receptors in Autoimmunity
7. Conclusions
Acknowledgements
References
- Janeway, C.A., Jr.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef]
- Yoneyama, M.; Fujita, T. RNA recognition and signal transduction by RIG-I-like receptors. Immunol. Rev. 2009, 227, 54–65. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. The roles of TLRs, RLRs and NLRs in pathogen recognition. Int. Immunol. 2009, 21, 317–337. [Google Scholar] [CrossRef]
- Ronald, P.C.; Beutler, B. Plant and animal sensors of conserved microbial signatures. Science 2010, 330, 1061–1064. [Google Scholar] [CrossRef]
- Iwasaki, A.; Medzhitov, R. Regulation of adaptive immunity by the innate immune system. Science 2010, 327, 291–295. [Google Scholar] [CrossRef]
- Basith, S.; Manavalan, B.; Yoo, T.H.; Kim, S.G.; Choi, S. Roles of Toll-like receptors in Cancer: A double-edged sword for defense and offense. Arch. Pharm. Res. 2012, 35, 1297–1316. [Google Scholar] [CrossRef]
- Manavalan, B.; Basith, S.; Choi, S. Similar Structures but Different Roles—An Updated Perspective on TLR Structures. Front. Physiol. 2011, 2011, 41. [Google Scholar]
- Basith, S.; Manavalan, B.; Govindaraj, R.G.; Choi, S. In silico approach to inhibition of signaling pathways of Toll-like receptors 2 and 4 by ST2L. PLoS One 2011, 6, e23989. [Google Scholar]
- Krishnan, J.; Lee, G.; Choi, S. Drugs targeting Toll-like receptors. Arch. Pharm. Res. 2009, 32, 1485–1502. [Google Scholar] [CrossRef]
- Krishnan, J.; Selvarajoo, K.; Tsuchiya, M.; Lee, G.; Choi, S. Toll-like receptor signal transduction. Exp. Mol. Med. 2007, 39, 421–438. [Google Scholar]
- Govindaraj, R.G.; Manavalan, B.; Lee, G.; Choi, S. Molecular modeling-based evaluation of hTLR10 and identification of potential ligands in Toll-like receptor signaling. PLoS One 2010, 5, e12713. [Google Scholar]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef]
- Hayashi, F.; Smith, K.D.; Ozinsky, A.; Hawn, T.R.; Yi, E.C.; Goodlett, D.R.; Eng, J.K.; Akira, S.; Underhill, D.M.; Aderem, A. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 2001, 410, 1099–1103. [Google Scholar] [CrossRef]
- Schwandner, R.; Dziarski, R.; Wesche, H.; Rothe, M.; Kirschning, C.J. Peptidoglycan- and lipoteichoic acid-induced cell activation is mediated by Toll-like receptor 2. J. Biol. Chem. 1999, 274, 17406–17409. [Google Scholar]
- Aliprantis, A.O.; Yang, R.B.; Mark, M.R.; Suggett, S.; Devaux, B.; Radolf, J.D.; Klimpel, G.R.; Godowski, P.; Zychlinsky, A. Cell activation and apoptosis by bacterial lipoproteins through Toll-like receptor-2. Science 1999, 285, 736–739. [Google Scholar] [CrossRef]
- Takeuchi, O.; Hoshino, K.; Kawai, T.; Sanjo, H.; Takada, H.; Ogawa, T.; Takeda, K.; Akira, S. Differential roles of TLR2 and TLR4 in recognition of gram-negative and gram-positive bacterial cell wall components. Immunity 1999, 11, 443–451. [Google Scholar] [CrossRef]
- Picker, L.J.; Butcher, E.C. Physiological and molecular mechanisms of lymphocyte homing. Annu. Rev. Immunol. 1992, 10, 561–591. [Google Scholar] [CrossRef]
- Blander, J.M.; Medzhitov, R. Regulation of phagosome maturation by signals from Toll-like receptors. Science 2004, 304, 1014–1018. [Google Scholar] [CrossRef]
- Oldenburg, M.; Kruger, A.; Ferstl, R.; Kaufmann, A.; Nees, G.; Sigmund, A.; Bathke, B.; Lauterbach, H.; Suter, M.; Dreher, S.; et al. TLR13 recognizes bacterial 23S rRNA devoid of erythromycin resistance-forming modification. Science 2012, 337, 1111–1115. [Google Scholar]
- Leadbetter, E.A.; Rifkin, I.R.; Hohlbaum, A.M.; Beaudette, B.C.; Shlomchik, M.J.; Marshak-Rothstein, A. Chromatin-IgG complexes activate B cells by dual engagement of IgM and Toll-like receptors. Nature 2002, 416, 603–607. [Google Scholar] [CrossRef]
- Viglianti, G.A.; Lau, C.M.; Hanley, T.M.; Miko, B.A.; Shlomchik, M.J.; Marshak-Rothstein, A. Activation of autoreactive B cells by CpG dsDNA. Immunity 2003, 19, 837–847. [Google Scholar] [CrossRef]
- Krieg, A.M. A role for Toll in autoimmunity. Nat. Immunol. 2002, 3, 423–424. [Google Scholar] [CrossRef]
- Baccala, R.; Hoebe, K.; Kono, D.H.; Beutler, B.; Theofilopoulos, A.N. TLR-dependent and TLR-independent pathways of type I interferon induction in systemic autoimmunity. Nat. Med. 2007, 13, 543–51. [Google Scholar] [CrossRef]
- Miyake, K. Innate immune sensing of pathogens and danger signals by cell surface Toll-like receptors. Semin. Immunol. 2007, 19, 3–10. [Google Scholar] [CrossRef]
- Zhu, J.; Brownlie, R.; Liu, Q.; Babiuk, L.A.; Potter, A.; Mutwiri, G.K. Characterization of bovine Toll-like receptor 8: Ligand specificity, signaling essential sites and dimerization. Mol. Immunol. 2009, 46, 978–990. [Google Scholar] [CrossRef]
- Medzhitov, R.; Preston-Hurlburt, P.; Kopp, E.; Stadlen, A.; Chen, C.; Ghosh, S.; Janeway, C.A., Jr. MyD88 is an adaptor protein in the hToll/IL-1 receptor family signaling pathways. Mol. Cell 1998, 2, 253–258. [Google Scholar] [CrossRef]
- Takeda, K.; Akira, S. TLR signaling pathways. Semin. Immunol. 2004, 16, 3–9. [Google Scholar] [CrossRef]
- Suzuki, N.; Suzuki, S.; Duncan, G.S.; Millar, D.G.; Wada, T.; Mirtsos, C.; Takada, H.; Wakeham, A.; Itie, A.; Li, S.; et al. Severe impairment of interleukin-1 and Toll-like receptor signalling in mice lacking IRAK-4. Nature 2002, 416, 750–756. [Google Scholar] [CrossRef]
- Kawagoe, T.; Sato, S.; Matsushita, K.; Kato, H.; Matsui, K.; Kumagai, Y.; Saitoh, T.; Kawai, T.; Takeuchi, O.; Akira, S. Sequential control of Toll-like receptor-dependent responses by IRAK1 and IRAK2. Nat. Immunol. 2008, 9, 684–691. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. Pathogen recognition with Toll-like receptors. Curr. Opin. Immunol. 2005, 17, 338–344. [Google Scholar] [CrossRef]
- O’Neill, L.A. How Toll-like receptors signal: What we know and what we don’t know. Curr. Opin. Immunol. 2006, 18, 3–9. [Google Scholar]
- Hoebe, K.; Du, X.; Georgel, P.; Janssen, E.; Tabeta, K.; Kim, S.O.; Goode, J.; Lin, P.; Mann, N.; Mudd, S.; et al. Identification of Lps2 as a key transducer of MyD88-independent TIR signalling. Nature 2003, 424, 743–748. [Google Scholar] [CrossRef]
- Fitzgerald, K.A.; McWhirter, S.M.; Faia, K.L.; Rowe, D.C.; Latz, E.; Golenbock, D.T.; Coyle, A.J.; Liao, S.M.; Maniatis, T. IKKepsilon and TBK1 are essential components of the IRF3 signaling pathway. Nat. Immunol. 2003, 4, 491–496. [Google Scholar]
- Pandey, S.; Agrawal, D.K. Immunobiology of Toll-like receptors: Emerging trends. Immunol. Cell Biol. 2006, 84, 333–341. [Google Scholar] [CrossRef]
- Lee, M.S.; Kim, Y.J. Signaling pathways downstream of pattern-recognition receptors and their cross talk. Annu. Rev. Biochem. 2007, 76, 447–480. [Google Scholar] [CrossRef]
- Matsumoto, M.; Funami, K.; Tanabe, M.; Oshiumi, H.; Shingai, M.; Seto, Y.; Yamamoto, A.; Seya, T. Subcellular localization of Toll-like receptor 3 in human dendritic cells. J. Immunol. 2003, 171, 3154–3162. [Google Scholar]
- Latz, E.; Schoenemeyer, A.; Visintin, A.; Fitzgerald, K.A.; Monks, B.G.; Knetter, C.F.; Lien, E.; Nilsen, N.J.; Espevik, T.; Golenbock, D.T. TLR9 signals after translocating from the ER to CpG DNA in the lysosome. Nat. Immunol. 2004, 5, 190–198. [Google Scholar]
- Hacker, H.; Mischak, H.; Miethke, T.; Liptay, S.; Schmid, R.; Sparwasser, T.; Heeg, K.; Lipford, G.B.; Wagner, H. CpG-DNA-specific activation of antigen-presenting cells requires stress kinase activity and is preceded by non-specific endocytosis and endosomal maturation. EMBO J. 1998, 17, 6230–6240. [Google Scholar] [CrossRef]
- Nishiya, T.; Kajita, E.; Miwa, S.; Defranco, A.L. TLR3 and TLR7 are targeted to the same intracellular compartments by distinct regulatory elements. J. Biol. Chem. 2005, 280, 37107–37117. [Google Scholar] [CrossRef]
- Kim, Y.M.; Brinkmann, M.M.; Paquet, M.E.; Ploegh, H.L. UNC93B1 delivers nucleotide-sensing Toll-like receptors to endolysosomes. Nature 2008, 452, 234–238. [Google Scholar] [CrossRef]
- Brinkmann, M.M.; Spooner, E.; Hoebe, K.; Beutler, B.; Ploegh, H.L.; Kim, Y.M. The interaction between the ER membrane protein UNC93B and TLR3, 7, and 9 is crucial for TLR signaling. J. Cell Biol. 2007, 177, 265–275. [Google Scholar] [CrossRef]
- Wang, J.; Shao, Y.; Bennett, T.A.; Shankar, R.A.; Wightman, P.D.; Reddy, L.G. The functional effects of physical interactions among Toll-like receptors 7, 8, and 9. J. Biol. Chem. 2006, 281, 37427–37434. [Google Scholar]
- Tabeta, K.; Hoebe, K.; Janssen, E.M.; Du, X.; Georgel, P.; Crozat, K.; Mudd, S.; Mann, N.; Sovath, S.; Goode, J.; et al. The Unc93b1 mutation 3d disrupts exogenous antigen presentation and signaling via Toll-like receptors 3, 7 and 9. Nat. Immunol. 2006, 7, 156–164. [Google Scholar]
- Casrouge, A.; Zhang, S.Y.; Eidenschenk, C.; Jouanguy, E.; Puel, A.; Yang, K.; Alcais, A.; Picard, C.; Mahfoufi, N.; Nicolas, N.; et al. Herpes simplex virus encephalitis in human UNC-93B deficiency. Science 2006, 314, 308–312. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, B.; Dai, J.; Srivastava, P.K.; Zammit, D.J.; Lefrancois, L.; Li, Z. Heat shock protein gp96 is a master chaperone for Toll-like receptors and is important in the innate function of macrophages. Immunity 2007, 26, 215–226. [Google Scholar] [CrossRef]
- Takahashi, K.; Shibata, T.; Akashi-Takamura, S.; Kiyokawa, T.; Wakabayashi, Y.; Tanimura, N.; Kobayashi, T.; Matsumoto, F.; Fukui, R.; Kouro, T.; et al. A protein associated with Toll-like receptor (TLR) 4 (PRAT4A) is required for TLR-dependent immune responses. J. Exp. Med. 2007, 204, 2963–2976. [Google Scholar] [CrossRef]
- Ewald, S.E.; Lee, B.L.; Lau, L.; Wickliffe, K.E.; Shi, G.P.; Chapman, H.A.; Barton, G.M. The ectodomain of Toll-like receptor 9 is cleaved to generate a functional receptor. Nature 2008, 456, 658–662. [Google Scholar] [CrossRef]
- Asagiri, M.; Hirai, T.; Kunigami, T.; Kamano, S.; Gober, H.J.; Okamoto, K.; Nishikawa, K.; Latz, E.; Golenbock, D.T.; Aoki, K.; et al. Cathepsin K-dependent Toll-like receptor 9 signaling revealed in experimental arthritis. Science 2008, 319, 624–627. [Google Scholar] [CrossRef]
- Matsumoto, F.; Saitoh, S.; Fukui, R.; Kobayashi, T.; Tanimura, N.; Konno, K.; Kusumoto, Y.; Akashi-Takamura, S.; Miyake, K. Cathepsins are required for Toll-like receptor 9 responses. Biochem. Biophys. Res. Commun. 2008, 367, 693–699. [Google Scholar] [CrossRef]
- Sepulveda, F.E.; Maschalidi, S.; Colisson, R.; Heslop, L.; Ghirelli, C.; Sakka, E.; Lennon-Dumenil, A.M.; Amigorena, S.; Cabanie, L.; Manoury, B. Critical role for asparagine endopeptidase in endocytic Toll-like receptor signaling in dendritic cells. Immunity 2009, 31, 737–748. [Google Scholar] [CrossRef]
- Park, B.; Brinkmann, M.M.; Spooner, E.; Lee, C.C.; Kim, Y.M.; Ploegh, H.L. Proteolytic cleavage in an endolysosomal compartment is required for activation of Toll-like receptor 9. Nat. Immunol. 2008, 9, 1407–1414. [Google Scholar] [CrossRef]
- Peter, M.E.; Kubarenko, A.V.; Weber, A.N.; Dalpke, A.H. Identification of an N-terminal recognition site in TLR9 that contributes to CpG-DNA-mediated receptor activation. J. Immunol. 2009, 182, 7690–7697. [Google Scholar]
- Garcia-Cattaneo, A.; Gobert, F.X.; Muller, M.; Toscano, F.; Flores, M.; Lescure, A.; del Nery, E.; Benaroch, P. Cleavage of Toll-like receptor 3 by cathepsins B and H is essential for signaling. Proc. Natl. Acad. Sci. USA 2012, 109, 9053–9058. [Google Scholar]
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to Virchow? Lancet 2001, 357, 539–545. [Google Scholar] [CrossRef]
- Marshall-Clarke, S.; Downes, J.E.; Haga, I.R.; Bowie, A.G.; Borrow, P.; Pennock, J.L.; Grencis, R.K.; Rothwell, P. Polyinosinic acid is a ligand for Toll-like receptor 3. J. Biol. Chem. 2007, 282, 24759–24766. [Google Scholar]
- Daffis, S.; Samuel, M.A.; Suthar, M.S.; Gale, M., Jr.; Diamond, M.S. Toll-like receptor 3 has a protective role against West Nile virus infection. J. Virol. 2008, 82, 10349–10358. [Google Scholar]
- Zhang, S.Y.; Jouanguy, E.; Ugolini, S.; Smahi, A.; Elain, G.; Romero, P.; Segal, D.; Sancho-Shimizu, V.; Lorenzo, L.; Puel, A.; et al. TLR3 deficiency in patients with herpes simplex encephalitis. Science 2007, 317, 1522–7152. [Google Scholar]
- Hardarson, H.S.; Baker, J.S.; Yang, Z.; Purevjav, E.; Huang, C.H.; Alexopoulou, L.; Li, N.; Flavell, R.A.; Bowles, N.E.; Vallejo, J.G. Toll-like receptor 3 is an essential component of the innate stress response in virus-induced cardiac injury. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H251–H258. [Google Scholar]
- Tabeta, K.; Georgel, P.; Janssen, E.; Du, X.; Hoebe, K.; Crozat, K.; Mudd, S.; Shamel, L.; Sovath, S.; Goode, J.; et al. Toll-like receptors 9 and 3 as essential components of innate immune defense against mouse cytomegalovirus infection. Proc. Natl. Acad. Sci. USA 2004, 101, 3516–3521. [Google Scholar]
- Salaun, B.; Coste, I.; Rissoan, M.C.; Lebecque, S.J.; Renno, T. TLR3 can directly trigger apoptosis in human cancer cells. J. Immunol. 2006, 176, 4894–4901. [Google Scholar]
- Salem, M.L.; El-Naggar, S.A.; Kadima, A.; Gillanders, W.E.; Cole, D.J. The adjuvant effects of the Toll-like receptor 3 ligand polyinosinic-cytidylic acid poly (I:C) on antigen-specific CD8+ T cell responses are partially dependent on NK cells with the induction of a beneficial cytokine milieu. Vaccine 2006, 24, 5119–5132. [Google Scholar] [CrossRef]
- Tewari, K.; Flynn, B.J.; Boscardin, S.B.; Kastenmueller, K.; Salazar, A.M.; Anderson, C.A.; Soundarapandian, V.; Ahumada, A.; Keler, T.; Hoffman, S.L.; et al. Poly(I:C) is an effective adjuvant for antibody and multi-functional CD4+ T cell responses to Plasmodium falciparum circumsporozoite protein (CSP) and alphaDEC-CSP in non human primates. Vaccine 2010, 28, 7256–7266. [Google Scholar]
- Zheng, R.; Cohen, P.A.; Paustian, C.A.; Johnson, T.D.; Lee, W.T.; Shu, S.; Koski, G.K. Paired Toll-like receptor agonists enhance vaccine therapy through induction of interleukin-12. Cancer Res. 2008, 68, 4045–4049. [Google Scholar]
- Longhi, M.P.; Trumpfheller, C.; Idoyaga, J.; Caskey, M.; Matos, I.; Kluger, C.; Salazar, A.M.; Colonna, M.; Steinman, R.M. Dendritic cells require a systemic type I interferon response to mature and induce CD4+ Th1 immunity with poly IC as adjuvant. J. Exp. Med. 2009, 206, 1589–1602. [Google Scholar] [CrossRef]
- Carter, W.A.; Hubbell, H.R.; Krueger, L.J.; Strayer, D.R. Comparative studies of ampligen (mismatched double-stranded RNA) and interferons. J. Biol. Response Mod. 1985, 4, 613–620. [Google Scholar]
- Hubbell, H.R.; Pequignot, E.C.; Todd, J.; Raymond, L.C.; Mayberry, S.D.; Carter, W.A.; Strayer, D.R. Augmented antitumor effect of combined human natural interferon-alpha and mismatched double-stranded RNA treatment against a human malignant melanoma xenograft. J. Biol. Response Mod. 1987, 6, 525–536. [Google Scholar]
- Shime, H.; Matsumoto, M.; Oshiumi, H.; Tanaka, S.; Nakane, A.; Iwakura, Y.; Tahara, H.; Inoue, N.; Seya, T. Toll-like receptor 3 signaling converts tumor-supporting myeloid cells to tumoricidal effectors. Proc. Natl. Acad. Sci. USA 2012, 109, 2066–2071. [Google Scholar]
- Akazawa, T.; Ebihara, T.; Okuno, M.; Okuda, Y.; Shingai, M.; Tsujimura, K.; Takahashi, T.; Ikawa, M.; Okabe, M.; Inoue, N.; et al. Antitumor NK activation induced by the Toll-like receptor 3-TICAM-1 (TRIF) pathway in myeloid dendritic cells. Proc. Natl. Acad. Sci. USA 2007, 104, 252–257. [Google Scholar]
- Navabi, H.; Jasani, B.; Reece, A.; Clayton, A.; Tabi, Z.; Donninger, C.; Mason, M.; Adams, M. A clinical grade poly I:C-analogue (Ampligen) promotes optimal DC maturation and Th1-type T cell responses of healthy donors and cancer patients in vitro. Vaccine 2009, 27, 107–115. [Google Scholar] [CrossRef]
- Strayer, D.R.; Carter, W.A.; Stouch, B.C.; Stevens, S.R.; Bateman, L.; Cimoch, P.J.; Lapp, C.W.; Peterson, D.L.; Mitchell, W.M. A double-blind, placebo-controlled, randomized, clinical trial of the TLR-3 agonist rintatolimod in severe cases of chronic fatigue syndrome. PLoS One 2012, 7, e31334. [Google Scholar]
- Shir, A.; Ogris, M.; Roedl, W.; Wagner, E.; Levitzki, A. EGFR-homing dsRNA activates cancer-targeted immune response and eliminates disseminated EGFR-overexpressing tumors in mice. Clin. Cancer Res. 2011, 17, 1033–1043. [Google Scholar] [CrossRef]
- Wang, L.; Smith, D.; Bot, S.; Dellamary, L.; Bloom, A.; Bot, A. Noncoding RNA danger motifs bridge innate and adaptive immunity and are potent adjuvants for vaccination. J. Clin. Invest. 2002, 110, 1175–1184. [Google Scholar]
- Sugiyama, T.; Hoshino, K.; Saito, M.; Yano, T.; Sasaki, I.; Yamazaki, C.; Akira, S.; Kaisho, T. Immunoadjuvant effects of polyadenylic:polyuridylic acids through TLR3 and TLR7. Int. Immunol. 2008, 20, 1–9. [Google Scholar]
- Conforti, R.; Ma, Y.; Morel, Y.; Paturel, C.; Terme, M.; Viaud, S.; Ryffel, B.; Ferrantini, M.; Uppaluri, R.; Schreiber, R.; et al. Opposing effects of Toll-like receptor (TLR3) signaling in tumors can be therapeutically uncoupled to optimize the anticancer efficacy of TLR3 ligands. Cancer Res. 2010, 70, 490–500. [Google Scholar] [CrossRef]
- Laplanche, A.; Alzieu, L.; Delozier, T.; Berlie, J.; Veyret, C.; Fargeot, P.; Luboinski, M.; Lacour, J. Polyadenylic-polyuridylic acid plus locoregional radiotherapy versus chemotherapy with CMF in operable breast cancer: A 14 year follow-up analysis of a randomized trial of the Federation Nationale des Centres de Lutte contre le Cancer (FNCLCC). Breast Cancer Res. Treat. 2000, 64, 189–191. [Google Scholar] [CrossRef]
- Jeung, H.C.; Moon, Y.W.; Rha, S.Y.; Yoo, N.C.; Roh, J.K.; Noh, S.H.; Min, J.S.; Kim, B.S.; Chung, H.C. Phase III trial of adjuvant 5-fluorouracil and adriamycin versus 5-fluorouracil, adriamycin, and polyadenylic-polyuridylic acid (poly A:U) for locally advanced gastric cancer after curative surgery: Final results of 15-year follow-up. Ann. Oncol. 2008, 19, 520–526. [Google Scholar]
- Lacour, J.; Laplanche, A.; Malafosse, M.; Gallot, D.; Julien, M.; Rotman, N.; Guivarc’h, M.; Roullet-Audy, J.C.; Lasser, P.; Hautefeuille, P.; et al. Polyadenylic-polyuridylic acid as an adjuvant in resectable colorectal carcinoma: A 6 1/2 year follow-up analysis of a multicentric double blind randomized trial. Eur. J. Surg. Oncol. 1992, 18, 599–604. [Google Scholar]
- Levy, H.B.; Baer, G.; Baron, S.; Buckler, C.E.; Gibbs, C.J.; Iadarola, M.J.; London, W.T.; Rice, J. A modified polyriboinosinic-polyribocytidylic acid complex that induces interferon in primates. J. Infect. Dis. 1975, 132, 434–439. [Google Scholar] [CrossRef]
- Hawkins, M.J.; Levin, M.; Borden, E.C. An Eastern Cooperative Oncology Group phase I-II pilot study of polyriboinosinic-polyribocytidylic acid poly-L-lysine complex in patients with metastatic malignant melanoma. J. Biol. Response Mod. 1985, 4, 664–668. [Google Scholar]
- Hennessy, E.J.; Parker, A.E.; O’Neill, L.A. Targeting Toll-like receptors: Emerging therapeutics? Nat. Rev. Drug Discov. 2010, 9, 293–307. [Google Scholar] [CrossRef]
- Panter, G.; Kuznik, A.; Jerala, R. Therapeutic applications of nucleic acids as ligands for Toll-like receptors. Curr. Opin. Mol. Ther. 2009, 11, 133–145. [Google Scholar]
- ClinicalTrials.gov. Available online: http://clinicaltrials.gov/ (accessed on 15 August 2012).
- Diebold, S.S.; Kaisho, T.; Hemmi, H.; Akira, S.; Reis e Sousa, C. Innate antiviral responses by means of TLR7-mediated recognition of single-stranded RNA. Science 2004, 303, 1529–1531. [Google Scholar]
- Heil, F.; Hemmi, H.; Hochrein, H.; Ampenberger, F.; Kirschning, C.; Akira, S.; Lipford, G.; Wagner, H.; Bauer, S. Species-specific recognition of single-stranded RNA via Toll-like receptor 7 and 8. Science 2004, 303, 1526–1529. [Google Scholar] [CrossRef]
- Forsbach, A.; Samulowitz, U.; Volp, K.; Hofmann, H.P.; Noll, B.; Tluk, S.; Schmitz, C.; Wader, T.; Muller, C.; Podszuweit, A.; et al. Dual or triple activation of TLR7, TLR8, and/or TLR9 by single-stranded oligoribonucleotides. Nucleic Acid Ther. 2011, 21, 423–436. [Google Scholar]
- Lund, J.M.; Alexopoulou, L.; Sato, A.; Karow, M.; Adams, N.C.; Gale, N.W.; Iwasaki, A.; Flavell, R.A. Recognition of single-stranded RNA viruses by Toll-like receptor 7. Proc. Natl. Acad. Sci. USA 2004, 101, 5598–5603. [Google Scholar]
- Forsbach, A.; Nemorin, J.G.; Montino, C.; Muller, C.; Samulowitz, U.; Vicari, A.P.; Jurk, M.; Mutwiri, G.K.; Krieg, A.M.; Lipford, G.B.; et al. Identification of RNA sequence motifs stimulating sequence-specific TLR8-dependent immune responses. J. Immunol. 2008, 180, 3729–2738. [Google Scholar]
- Jurk, M.; Heil, F.; Vollmer, J.; Schetter, C.; Krieg, A.M.; Wagner, H.; Lipford, G.; Bauer, S. Human TLR7 or TLR8 independently confer responsiveness to the antiviral compound R-848. Nat. Immunol. 2002, 3, 499. [Google Scholar]
- Demaria, O.; Pagni, P.P.; Traub, S.; de Gassart, A.; Branzk, N.; Murphy, A.J.; Valenzuela, D.M.; Yancopoulos, G.D.; Flavell, R.A.; Alexopoulou, L. TLR8 deficiency leads to autoimmunity in mice. J. Clin. Invest. 2010, 120, 3651–3662. [Google Scholar]
- Imbertson, L.M.; Beaurline, J.M.; Couture, A.M.; Gibson, S.J.; Smith, R.M.; Miller, R.L.; Reiter, M.J.; Wagner, T.L.; Tomai, M.A. Cytokine induction in hairless mouse and rat skin after topical application of the immune response modifiers imiquimod and S-28463. J. Invest. Dermatol. 1998, 110, 734–739. [Google Scholar] [CrossRef]
- Reiter, M.J.; Testerman, T.L.; Miller, R.L.; Weeks, C.E.; Tomai, M.A. Cytokine induction in mice by the immunomodulator imiquimod. J. Leukoc. Biol. 1994, 55, 234–240. [Google Scholar]
- Testerman, T.L.; Gerster, J.F.; Imbertson, L.M.; Reiter, M.J.; Miller, R.L.; Gibson, S.J.; Wagner, T.L.; Tomai, M.A. Cytokine induction by the immunomodulators imiquimod and S-27609. J. Leukoc. Biol. 1995, 58, 365–372. [Google Scholar]
- Miller, R.L.; Gerster, J.F.; Owens, M.L.; Slade, H.B.; Tomai, M.A. Review Article Imiquimod applied topically: A novel immune response modifier and new class of drug. Int. J. Immunopharmacol. 1999, 21, 1–14. [Google Scholar] [CrossRef]
- Miller, R.L.; Meng, T.C.; Tomai, M.A. The antiviral activity of Toll-like receptor 7 and 7/8 agonists. Drug News Perspect. 2008, 21, 69–87. [Google Scholar] [CrossRef]
- Wagner, T.L.; Ahonen, C.L.; Couture, A.M.; Gibson, S.J.; Miller, R.L.; Smith, R.M.; Reiter, M.J.; Vasilakos, J.P.; Tomai, M.A. Modulation of TH1 and TH2 cytokine production with the immune response modifiers, R-848 and imiquimod. Cell. Immunol. 1999, 191, 10–19. [Google Scholar] [CrossRef]
- Bernstein, D.I.; Harrison, C.J.; Tomai, M.A.; Miller, R.L. Daily or weekly therapy with resiquimod (R-848) reduces genital recurrences in herpes simplex virus-infected guinea pigs during and after treatment. J. Infect. Dis. 2001, 183, 844–849. [Google Scholar] [CrossRef]
- Schon, M.P.; Schon, M. TLR7 and TLR8 as targets in cancer therapy. Oncogene 2008, 27, 190–199. [Google Scholar] [CrossRef]
- Pockros, P.J.; Guyader, D.; Patton, H.; Tong, M.J.; Wright, T.; McHutchison, J.G.; Meng, T.C. Oral resiquimod in chronic HCV infection: Safety and efficacy in 2 placebo-controlled, double-blind phase IIa studies. J. Hepatol. 2007, 47, 174–182. [Google Scholar] [CrossRef]
- Butchi, N.B.; Pourciau, S.; Du, M.; Morgan, T.W.; Peterson, K.E. Analysis of the neuroinflammatory response to TLR7 stimulation in the brain: Comparison of multiple TLR7 and/or TLR8 agonists. J. Immunol. 2008, 180, 7604–7612. [Google Scholar]
- Salio, M.; Speak, A.O.; Shepherd, D.; Polzella, P.; Illarionov, P.A.; Veerapen, N.; Besra, G.S.; Platt, F.M.; Cerundolo, V. Modulation of human natural killer T cell ligands on TLR-mediated antigen-presenting cell activation. Proc. Natl. Acad. Sci. USA 2007, 104, 20490–20495. [Google Scholar]
- Dummer, R.; Hauschild, A.; Becker, J.C.; Grob, J.J.; Schadendorf, D.; Tebbs, V.; Skalsky, J.; Kaehler, K.C.; Moosbauer, S.; Clark, R.; et al. An exploratory study of systemic administration of the Toll-like receptor-7 agonist 852A in patients with refractory metastatic melanoma. Clin. Cancer Res. 2008, 14, 856–864. [Google Scholar] [CrossRef]
- Gorden, K.B.; Gorski, K.S.; Gibson, S.J.; Kedl, R.M.; Kieper, W.C.; Qiu, X.; Tomai, M.A.; Alkan, S.S.; Vasilakos, J.P. Synthetic TLR agonists reveal functional differences between human TLR7 and TLR8. J. Immunol. 2005, 174, 1259–1268. [Google Scholar]
- Velasquez, L.S.; Hjelm, B.E.; Arntzen, C.J.; Herbst-Kralovetz, M.M. An intranasally delivered Toll-like receptor 7 agonist elicits robust systemic and mucosal responses to Norwalk virus-like particles. Clin. Vaccine Immunol. 2010, 17, 1850–1858. [Google Scholar] [CrossRef]
- Pope, B.L.; Sigindere, J.; Chourmouzis, E.; MacIntyre, P.; Goodman, M.G. 7-Allyl-8-oxoguanosine (loxoribine) inhibits the metastasis of B16 melanoma cells and has adjuvant activity in mice immunized with a B16 tumor vaccine. Cancer Immunol. Immunother. 1994, 38, 83–91. [Google Scholar] [CrossRef]
- Heil, F.; Ahmad-Nejad, P.; Hemmi, H.; Hochrein, H.; Ampenberger, F.; Gellert, T.; Dietrich, H.; Lipford, G.; Takeda, K.; Akira, S.; et al. The Toll-like receptor 7 (TLR7)-specific stimulus loxoribine uncovers a strong relationship within the TLR7, 8 and 9 subfamily. Eur. J. Immunol. 2003, 33, 2987–2997. [Google Scholar] [CrossRef]
- Lee, J.; Chuang, T.H.; Redecke, V.; She, L.; Pitha, P.M.; Carson, D.A.; Raz, E.; Cottam, H.B. Molecular basis for the immunostimulatory activity of guanine nucleoside analogs: Activation of Toll-like receptor 7. Proc. Natl. Acad. Sci. USA 2003, 100, 6646–6651. [Google Scholar]
- Kang, S.S.; Kauls, L.S.; Gaspari, A.A. Toll-like receptors: Applications to dermatologic disease. J. Am. Acad. Dermatol. 2006, 54, 951–983. [Google Scholar] [CrossRef]
- Sarosdy, M.F. A review of clinical studies of bropirimine immunotherapy of carcinoma in situ of the bladder and upper urinary tract. Eur. Urol. 1997, 31 (Suppl. 1), 20–26. [Google Scholar]
- Tei, Y.; Matsuyama, H.; Wada, T.; Kurisu, H.; Tahara, M.; Naito, K. In vitro antitumor activity of bropirimine against urinary bladder tumor cells. Anticancer Res. 2002, 22, 1667–1671. [Google Scholar]
- Lee, J.; Wu, C.C.; Lee, K.J.; Chuang, T.H.; Katakura, K.; Liu, Y.T.; Chan, M.; Tawatao, R.; Chung, M.; Shen, C.; et al. Activation of anti-hepatitis C virus responses via Toll-like receptor 7. Proc. Natl. Acad. Sci. USA 2006, 103, 1828–1833. [Google Scholar]
- Horsmans, Y.; Berg, T.; Desager, J.P.; Mueller, T.; Schott, E.; Fletcher, S.P.; Steffy, K.R.; Bauman, L.A.; Kerr, B.M.; Averett, D.R. Isatoribine, an agonist of TLR7, reduces plasma virus concentration in chronic hepatitis C infection. Hepatology 2005, 42, 724–731. [Google Scholar] [CrossRef]
- Fletcher, S.; Steffy, K.; Averett, D. Masked oral prodrugs of Toll-like receptor 7 agonists: A new approach for the treatment of infectious disease. Curr. Opin. Investig. Drugs 2006, 7, 702–708. [Google Scholar]
- Kronenberger, B.; Zeuzem, S. Current and future treatment options for HCV. Ann. Hepatol. 2009, 8, 103–112. [Google Scholar]
- Kaisho, T.; Akira, S. Toll-like receptor function and signaling. J. Allergy Clin. Immunol. 2006, 117, 979–987. [Google Scholar]
- Hemmi, H.; Takeuchi, O.; Kawai, T.; Kaisho, T.; Sato, S.; Sanjo, H.; Matsumoto, M.; Hoshino, K.; Wagner, H.; Takeda, K.; et al. A Toll-like receptor recognizes bacterial DNA. Nature 2000, 408, 740–745. [Google Scholar]
- Krieg, A.M. Therapeutic potential of Toll-like receptor 9 activation. Nat. Rev. Drug Discov. 2006, 5, 471–484. [Google Scholar] [CrossRef]
- Barton, G.M.; Kagan, J.C.; Medzhitov, R. Intracellular localization of Toll-like receptor 9 prevents recognition of self DNA but facilitates access to viral DNA. Nat. Immunol. 2006, 7, 49–56. [Google Scholar] [CrossRef]
- Carpentier, A.; Laigle-Donadey, F.; Zohar, S.; Capelle, L.; Behin, A.; Tibi, A.; Martin-Duverneuil, N.; Sanson, M.; Lacomblez, L.; Taillibert, S.; et al. Phase 1 trial of a CpG oligodeoxynucleotide for patients with recurrent glioblastoma. Neuro Oncol. 2006, 8, 60–66. [Google Scholar] [CrossRef]
- Vollmer, J.; Krieg, A.M. Immunotherapeutic applications of CpG oligodeoxynucleotide TLR9 agonists. Adv. Drug Deliv. Rev. 2009, 61, 195–204. [Google Scholar] [CrossRef]
- Agrawal, S.; Kandimalla, E.R. Synthetic agonists of Toll-like receptors 7, 8 and 9. Biochem. Soc. Trans. 2007, 35, 1461–1467. [Google Scholar] [CrossRef]
- Wang, D.; Li, Y.; Yu, D.; Song, S.S.; Kandimalla, E.R.; Agrawal, S. Immunopharmacological and antitumor effects of second-generation immunomodulatory oligonucleotides containing synthetic CpR motifs. Int. J. Oncol. 2004, 24, 901–908. [Google Scholar]
- Kline, J.N.; Krieg, A.M. Toll-like receptor 9 activation with CpG oligodeoxynucleotides for asthma therapy. Drug News Perspect. 2008, 21, 434–439. [Google Scholar]
- Kochling, J.; Prada, J.; Bahrami, M.; Stripecke, R.; Seeger, K.; Henze, G.; Wittig, B.; Schmidt, M. Anti-tumor effect of DNA-based vaccination and dSLIM immunomodulatory molecules in mice with Ph+ acute lymphoblastic leukaemia. Vaccine 2008, 26, 4669–4675. [Google Scholar] [CrossRef]
- Horner, A.A.; Widhopf, G.F.; Burger, J.A.; Takabayashi, K.; Cinman, N.; Ronaghy, A.; Spiegelberg, H.L.; Raz, E. Immunostimulatory DNA inhibits IL-4-dependent IgE synthesis by human B cells. J. Allergy Clin. Immunol. 2001, 108, 417–423. [Google Scholar] [CrossRef]
- Rakoff-Nahoum, S.; Medzhitov, R. Toll-like receptors and cancer. Nat. Rev. Cancer 2009, 9, 57–63. [Google Scholar] [CrossRef]
- Barry, M.; Cooper, C. Review of hepatitis B surface antigen-1018 ISS adjuvant-containing vaccine safety and efficacy. Expert Opin. Biol. Ther. 2007, 7, 1731–1737. [Google Scholar] [CrossRef]
- Data From Two Phase 3 Studies Demonstrate HEPLISAV’s Superiority in Immunizing Persons With Diabetes From Hepatitis B. Available online: http://www.evaluatepharma.com/Universal/View.aspx?type=Story&id=260991 (accessed on 28 August 2012).
- Harandi, A.M.; Davies, G.; Olesen, O.F. Vaccine adjuvants: Scientific challenges and strategic initiatives. Expert Rev. Vaccines 2009, 8, 293–298. [Google Scholar] [CrossRef]
- Hepatitis C. Available online: http://www.dynavax.com/hepatitis_ct.html (accessed on 28 August 2012).
- Krieg, A.M.; Yi, A.K.; Matson, S.; Waldschmidt, T.J.; Bishop, G.A.; Teasdale, R.; Koretzky, G.A.; Klinman, D.M. CpG motifs in bacterial DNA trigger direct B-cell activation. Nature 1995, 374, 546–549. [Google Scholar]
- Manegold, C.; Gravenor, D.; Woytowitz, D.; Mezger, J.; Hirsh, V.; Albert, G.; Al-Adhami, M.; Readett, D.; Krieg, A.M.; Leichman, C.G. Randomized phase II trial of a Toll-like receptor 9 agonist oligodeoxynucleotide, PF-3512676, in combination with first-line taxane plus platinum chemotherapy for advanced-stage non-small-cell lung cancer. J. Clin. Oncol. 2008, 26, 3979–3986. [Google Scholar] [CrossRef]
- Zent, C.S.; Smith, B.J.; Ballas, Z.K.; Wooldridge, J.E.; Link, B.K.; Call, T.G.; Shanafelt, T.D.; Bowen, D.A.; Kay, N.E.; Witzig, T.E.; et al. Phase I clinical trial of CpG oligonucleotide 7909 (PF-03512676) in patients with previously treated chronic lymphocytic leukemia. Leuk. Lymphoma 2012, 53, 211–217. [Google Scholar] [CrossRef]
- Dorn, A.; Kippenberger, S. Clinical application of CpG-, non-CpG-, and antisense oligodeoxynucleotides as immunomodulators. Curr. Opin. Mol. Ther. 2008, 10, 10–20. [Google Scholar]
- Heijink, I.H.; van Oosterhout, A.J. Strategies for targeting T-cells in allergic diseases and asthma. Pharmacol. Ther. 2006, 112, 489–500. [Google Scholar] [CrossRef]
- Pastorelli, L.; Pizarro, T.T.; Cominelli, F.; Vecchi, M. Emerging drugs for the treatment of ulcerative colitis. Expert Opin. Emerg. Drugs 2009, 14, 505–521. [Google Scholar] [CrossRef]
- Vollmer, J.; Tluk, S.; Schmitz, C.; Hamm, S.; Jurk, M.; Forsbach, A.; Akira, S.; Kelly, K.M.; Reeves, W.H.; Bauer, S.; et al. Immune stimulation mediated by autoantigen binding sites within small nuclear RNAs involves Toll-like receptors 7 and 8. J. Exp. Med. 2005, 202, 1575–1585. [Google Scholar] [CrossRef]
- Christensen, S.R.; Shupe, J.; Nickerson, K.; Kashgarian, M.; Flavell, R.A.; Shlomchik, M.J. Toll-like receptor 7 and TLR9 dictate autoantibody specificity and have opposing inflammatory and regulatory roles in a murine model of lupus. Immunity 2006, 25, 417–428. [Google Scholar] [CrossRef]
- Sun, S.; Rao, N.L.; Venable, J.; Thurmond, R.; Karlsson, L. TLR7/9 antagonists as therapeutics for immune-mediated inflammatory disorders. Inflamm. Allergy Drug Targets 2007, 6, 223–235. [Google Scholar] [CrossRef]
- Parkinson, T. The future of Toll-like receptor therapeutics. Curr. Opin. Mol. Ther. 2008, 10, 21–31. [Google Scholar]
- Autoimmune. Available online: http://www.iderapharma.com/clinical/autoimmune.html (accessed on 12 September 2012).
- Barrat, F.J.; Meeker, T.; Chan, J.H.; Guiducci, C.; Coffman, R.L. Treatment of lupus-prone mice with a dual inhibitor of TLR7 and TLR9 leads to reduction of autoantibody production and amelioration of disease symptoms. Eur. J. Immunol. 2007, 37, 3582–3586. [Google Scholar] [CrossRef]
- Pawar, R.D.; Ramanjaneyulu, A.; Kulkarni, O.P.; Lech, M.; Segerer, S.; Anders, H.J. Inhibition of Toll-like receptor-7 (TLR-7) or TLR-7 plus TLR-9 attenuates glomerulonephritis and lung injury in experimental lupus. J. Am. Soc. Nephrol. 2007, 18, 1721–1731. [Google Scholar] [CrossRef]
- Napirei, M.; Karsunky, H.; Zevnik, B.; Stephan, H.; Mannherz, H.G.; Moroy, T. Features of systemic lupus erythematosus in Dnase1-deficient mice. Nat. Genet. 2000, 25, 177–181. [Google Scholar] [CrossRef]
- Yasutomo, K.; Horiuchi, T.; Kagami, S.; Tsukamoto, H.; Hashimura, C.; Urushihara, M.; Kuroda, Y. Mutation of DNASE1 in people with systemic lupus erythematosus. Nat. Genet. 2001, 28, 313–314. [Google Scholar] [CrossRef]
- Lau, C.M.; Broughton, C.; Tabor, A.S.; Akira, S.; Flavell, R.A.; Mamula, M.J.; Christensen, S.R.; Shlomchik, M.J.; Viglianti, G.A.; Rifkin, I.R.; et al. RNA-associated autoantigens activate B cells by combined B cell antigen receptor/Toll-like receptor 7 engagement. J. Exp. Med. 2005, 202, 1171–1177. [Google Scholar] [CrossRef]
- Hooks, J.J.; Moutsopoulos, H.M.; Geis, S.A.; Stahl, N.I.; Decker, J.L.; Notkins, A.L. Immune interferon in the circulation of patients with autoimmune disease. N. Engl. J. Med. 1979, 301, 5–8. [Google Scholar] [CrossRef]
- Ytterberg, S.R.; Schnitzer, T.J. Serum interferon levels in patients with systemic lupus erythematosus. Arthritis Rheum. 1982, 25, 401–406. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef]
- Lovgren, T.; Eloranta, M.L.; Bave, U.; Alm, G.V.; Ronnblom, L. Induction of interferon-alpha production in plasmacytoid dendritic cells by immune complexes containing nucleic acid released by necrotic or late apoptotic cells and lupus IgG. Arthritis Rheum. 2004, 50, 1861–1872. [Google Scholar] [CrossRef]
- Stott, K.; Tang, G.S.; Lee, K.B.; Thomas, J.O. Structure of a complex of tandem HMG boxes and DNA. J. Mol. Biol. 2006, 360, 90–104. [Google Scholar] [CrossRef]
- Jego, G.; Palucka, A.K.; Blanck, J.P.; Chalouni, C.; Pascual, V.; Banchereau, J. Plasmacytoid dendritic cells induce plasma cell differentiation through type I interferon and interleukin 6. Immunity 2003, 19, 225–234. [Google Scholar] [CrossRef]
- Izui, S.; Iwamoto, M.; Fossati, L.; Merino, R.; Takahashi, S.; Ibnou-Zekri, N. The Yaa gene model of systemic lupus erythematosus. Immunol. Rev. 1995, 144, 137–156. [Google Scholar] [CrossRef]
- Deng, G.M.; Nilsson, I.M.; Verdrengh, M.; Collins, L.V.; Tarkowski, A. Intra-articularly localized bacterial DNA containing CpG motifs induces arthritis. Nat. Med. 1999, 5, 702–705. [Google Scholar] [CrossRef]
- Prinz, M.; Garbe, F.; Schmidt, H.; Mildner, A.; Gutcher, I.; Wolter, K.; Piesche, M.; Schroers, R.; Weiss, E.; Kirschning, C.J.; et al. Innate immunity mediated by TLR9 modulates pathogenicity in an animal model of multiple sclerosis. J. Clin. Invest. 2006, 116, 456–464. [Google Scholar] [CrossRef]
- Wu, H.J.; Sawaya, H.; Binstadt, B.; Brickelmaier, M.; Blasius, A.; Gorelik, L.; Mahmood, U.; Weissleder, R.; Carulli, J.; Benoist, C.; et al. Inflammatory arthritis can be reined in by CpG-induced DC-NK cell cross talk. J. Exp. Med. 2007, 204, 1911–1922. [Google Scholar]
- Cherfils-Vicini, J.; Platonova, S.; Gillard, M.; Laurans, L.; Validire, P.; Caliandro, R.; Magdeleinat, P.; Mami-Chouaib, F.; Dieu-Nosjean, M.C.; Fridman, W.H.; et al. Triggering of TLR7 and TLR8 expressed by human lung cancer cells induces cell survival and chemoresistance. J. Clin. Invest. 2010, 120, 1285–1297. [Google Scholar] [CrossRef]
- Chiron, D.; Pellat-Deceunynck, C.; Amiot, M.; Bataille, R.; Jego, G. TLR3 ligand induces NF-{kappa}B activation and various fates of multiple myeloma cells depending on IFN-{alpha} production. J. Immunol. 2009, 182, 4471–4478. [Google Scholar] [CrossRef]
- Gorden, K.K.; Qiu, X.X.; Binsfeld, C.C.; Vasilakos, J.P.; Alkan, S.S. Cutting edge: Activation of murine TLR8 by a combination of imidazoquinoline immune response modifiers and polyT oligodeoxynucleotides. J. Immunol. 2006, 177, 6584–6587. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gosu, V.; Basith, S.; Kwon, O.-P.; Choi, S. Therapeutic Applications of Nucleic Acids and Their Analogues in Toll-like Receptor Signaling. Molecules 2012, 17, 13503-13529. https://doi.org/10.3390/molecules171113503
Gosu V, Basith S, Kwon O-P, Choi S. Therapeutic Applications of Nucleic Acids and Their Analogues in Toll-like Receptor Signaling. Molecules. 2012; 17(11):13503-13529. https://doi.org/10.3390/molecules171113503
Chicago/Turabian StyleGosu, Vijayakumar, Shaherin Basith, O-Pil Kwon, and Sangdun Choi. 2012. "Therapeutic Applications of Nucleic Acids and Their Analogues in Toll-like Receptor Signaling" Molecules 17, no. 11: 13503-13529. https://doi.org/10.3390/molecules171113503