Oligonucleotide-Templated Reactions for Sensing Nucleic Acids

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Templated Photochemical Reaction


3. The Templated Ester-Hydrolysis Reaction

4. The Templated Nucleophilic Substitution Reaction


5. The Templated Staudinger Reaction

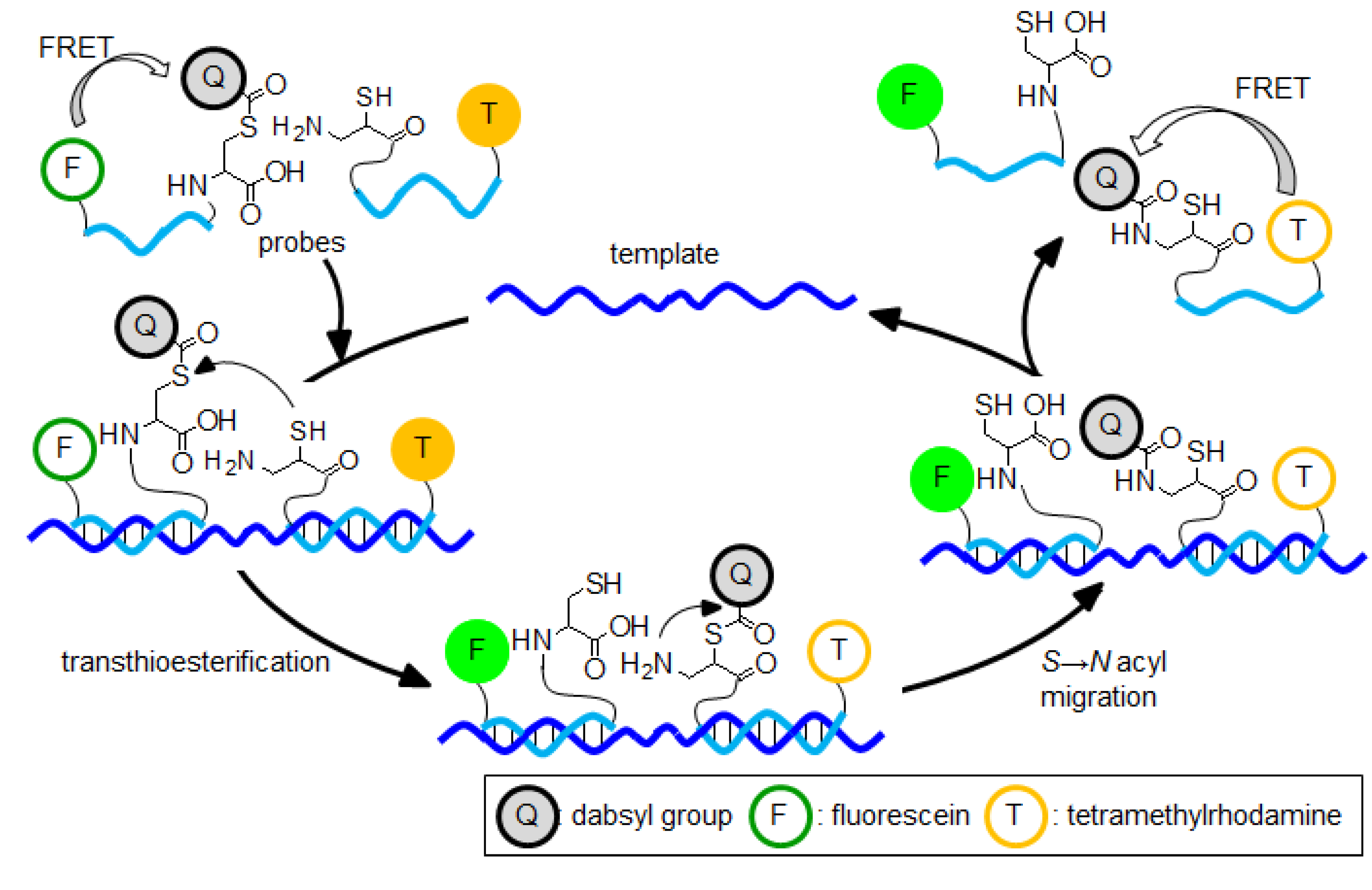
6. Templated Peptide Chemical Reaction

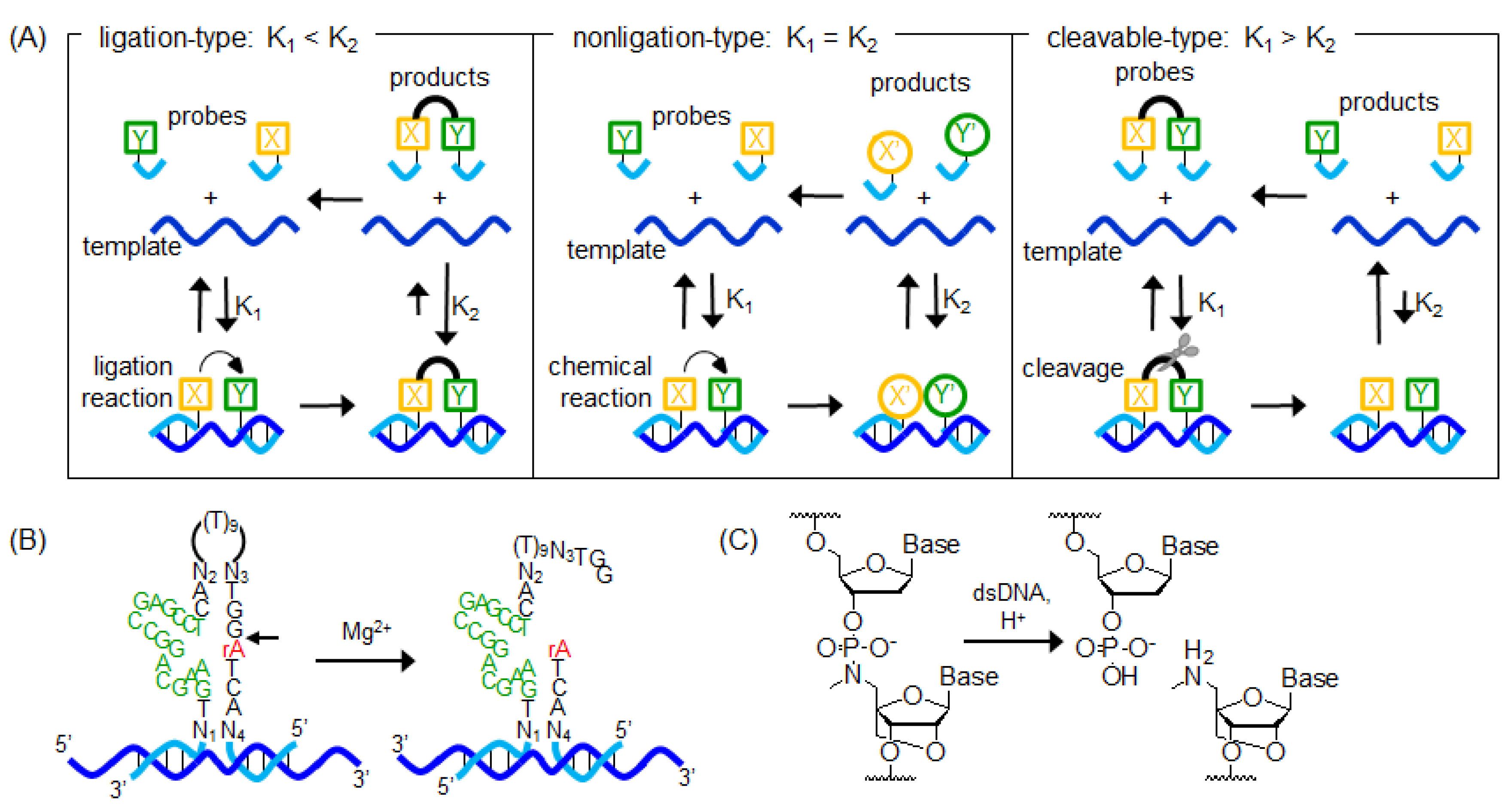
7. Other Templated Reactions


8. Conclusions
Acknowledgments
References
- Silverman, A.P.; Kool, E.T. Detecting RNA and DNA with templated chemical reactions. Chem. Rev. 2006, 106, 3775–3789. [Google Scholar] [CrossRef]
- Grossmann, T.N.; Strohbach, A.; Seitz, O. Achieving turnover in DNA-templated reactions. ChemBioChem 2008, 9, 2185–2192. [Google Scholar] [CrossRef]
- Lewis, R.J.; Hanawalt, P.C. Ligation of oligonucleotides by pyrimidine dimers—A missing “link” in the origin of life? Nature 1982, 298, 393–396. [Google Scholar]
- Churchill, M.E.; Peak, J.G.; Peak, M.J. Correlation between cell survival and DNA single-strand break repair proficiency in the Chinese hamster ovary cell lines AA8 and EM9 irradiated with 365-nm ultraviolet-A radiation. Photochem. Photobiol. 1991, 53, 229–236. [Google Scholar] [CrossRef]
- Albagli, D.; van Atta, R.; Cheng, P.; Huan, B.F.; Wood, M.L. Chemical amplification (CHAMP) by a continuous, self-replicating oligonucleotide-based system. J. Am. Chem. Soc. 1999, 121, 6954–6955. [Google Scholar] [CrossRef]
- Woo, J.; Hopkins, P.B. Template directed modification of single stranded DNA by psoralen-tethered oligonucleotides: Sites of photoadduct formation analyzed by sequence-specific and sequence-random cleavage. J. Am. Chem. Soc. 1991, 113, 5457–5459. [Google Scholar] [CrossRef]
- Lewis, F.D.; Wu, T.F.; Burch, E.L.; Bassani, D.M.; Yang, J.S.; Schneider, S.; Jager, W.; Letsinger, R.L. Hybrid oligonucleotides containing stilbene units. Excimer fluorescence and photodimerization. J. Am. Chem. Soc. 1995, 117, 8785–8792. [Google Scholar]
- Letsinger, R.L.; Wu, T.F. Control of excimer emission and photochemistry of stilbene units by oligonucleotide hybridization. J. Am. Chem. Soc. 1994, 116, 811–812. [Google Scholar] [CrossRef]
- Liu, J.Q.; Taylor, J.S. Remarkable photoreversal of a thio analog of the dewar valence isomer of the (6-4) photoproduct of DNA to the parent nucleotides. J. Am. Chem. Soc. 1996, 118, 3287–3288. [Google Scholar] [CrossRef]
- Zhao, X.D.; Liu, J.Q.; Hsu, D.S.; Zhao, S.Y.; Taylor, J.S.; Sancar, A. Reaction mechanism of (6-4) photolyase. J. Biol. Chem. 1997, 272, 32580–32590. [Google Scholar]
- Liu, J.Q.; Taylor, J.S. Template-directed photoligation of oligodeoxyribonucleotides via 4-thiothymidine. Nucleic Acids Res. 1998, 26, 3300–3304. [Google Scholar] [CrossRef]
- Fujimoto, K.; Matsuda, S.; Takahashi, N.; Saito, I. Template-directed photoreversible ligation of deoxyoligonucleotides via 5-vinyldeoxyuridine. J. Am. Chem. Soc. 2000, 122, 5646–5647. [Google Scholar] [CrossRef]
- Fujimoto, K.; Matsuda, S.; Ogawa, N.; Hayashi, M.; Saito, I. Template-directed reversible photocircularization of DNA via 5-vinyldeoxycytidine. Tetrahedron Lett. 2000, 41, 6451–6454. [Google Scholar]
- Fujimoto, K.; Matsuda, S.; Hayashi, M.; Saito, I. Reversible DNA photocircularization on triple helix: Effect of vinyl substituent on base stacking. Tetrahedron Lett. 2000, 41, 7897–7900. [Google Scholar] [CrossRef]
- Fujimoto, K.; Ogawa, N.; Hayashi, M.; Matsuda, S.; Saito, I. Template directed photochemical synthesis of branched oligodeoxynucleotides via 5-carboxyvinyldeoxyuridine. Tetrahedron Lett. 2000, 41, 9437–9440. [Google Scholar] [CrossRef]
- Fujimoto, K.; Matsuda, S.; Yoshimura, Y.; Matsumura, T.; Hayashi, M.; Saito, I. Site-specific transition of cytosine to uracil via reversible DNA photoligation. Chem. Commun. 2006, 3223–3225. [Google Scholar]
- Fujimoto, K.; Matsuda, S.; Yoshimura, Y.; Ami, T.; Saito, I. Reversible photopadlocking on double-stranded DNA. Chem. Commun. 2007, 2968–2970. [Google Scholar]
- Ogasawara, S.; Fujimoto, K. Solution of a SAT problem on a photochemical DNA computer. Chem. Lett. 2005, 34, 378–379. [Google Scholar] [CrossRef]
- Yoshimura, Y.; Noguchi, Y.; Sato, H.; Fujimoto, K. Template-directed DNA photoligation in rapid and selective detection of RNA point mutations. ChemBioChem 2006, 7, 598–601. [Google Scholar] [CrossRef]
- Fujimoto, K.; Yoshino, H.; Ami, T.; Yoshimura, Y.; Saito, I. A light-controlled reversible DNA photoligation via carbazole-tethered 5-carboxyvinyluracil. Org. Lett. 2008, 10, 397–400. [Google Scholar] [CrossRef]
- Nakamura, S.; Ogasawara, S.; Matuda, S.; Saito, I.; Fujimoto, K. Template directed reversible photochemical ligation of oligodeoxynucleotides. Molecules 2011, 17, 163–178. [Google Scholar] [CrossRef]
- Saito, I.; Miyauchi, Y.; Saito, Y.; Fujimoto, K. Template-directed photoreversible ligation of DNA via 7-carboxyvinyl-7-deaza-2'-deoxyadenosine. Tetrahedron Lett. 2005, 46, 97–99. [Google Scholar] [CrossRef]
- Yoshimura, Y.; Okamura, D.; Ogino, M.; Fujimoto, K. Highly selective and sensitive template-directed photoligation of DNA via 5-carbamoylvinyl-2'-deoxycytidine. Org. Lett. 2006, 8, 5049–5051. [Google Scholar] [CrossRef]
- Ami, T.; Fujimoto, K. Click chemistry as an efficient method for preparing a sensitive DNA probe for photochemical ligation. ChemBioChem 2008, 9, 2071–2074. [Google Scholar] [CrossRef]
- Ami, T.; Yoshimura, Y.; Matsuzaki, T.; Fujimoto, K. Photochemical ligation of DNA probe prepared in click chemistry. J. Photopolym. Sci. Technol. 2009, 22, 267–272. [Google Scholar] [CrossRef]
- Ogino, M.; Yoshimura, Y.; Nakazawa, A.; Saito, I.; Fujimoto, K. Template-directed DNA photoligation via alpha-5-cyanovinyldeoxyuridine. Org. Lett. 2005, 7, 2853–2856. [Google Scholar] [CrossRef]
- Ami, T.; Fujimoto, K. Fluorescence labeling of DNA based on photochemical ligation. Sci. Technol. Adv. Mater. 2006, 7, 249–254. [Google Scholar] [CrossRef]
- Fujimoto, K.; Konishi-Hiratsuka, K.; Sakamoto, T.; Yoshimura, Y. Site-specific photochemical RNA editing. Chem. Commun. 2010, 46, 7545–7547. [Google Scholar]
- Fujimoto, K.; Konishi-Hiratsuka, K.; Sakamoto, T.; Yoshimura, Y. Site-specific cytosine to uracil transition by using reversible DNA photo-crosslinking. ChemBioChem 2010, 11, 1661–1664. [Google Scholar] [CrossRef]
- Ihara, T.; Fujii, T.; Mukae, M.; Kitamura, Y.; Jyo, A. Photochemical ligation of DNA conjugates through anthracene cyclodimer formation and its fidelity to the template sequences. J. Am. Chem. Soc. 2004, 126, 8880–8881. [Google Scholar] [CrossRef]
- Arslan, P.; Ihara, T.; Mukae, M.; Jyo, A. The effect of local structural disruption on the yield of photochemical ligation between anthracene-oligonucleotide conjugates. Anal. Sci. 2008, 24, 173–176. [Google Scholar] [CrossRef]
- Mukae, M.; Ihara, T.; Tabara, M.; Jyo, A. Anthracene-DNA conjugates as building blocks of designed DNA structures constructed by photochemical reactions. Org. Biomol. Chem. 2009, 7, 1349–1354. [Google Scholar] [CrossRef]
- Arslan, P.; Jyo, A.; Ihara, T. Reversible circularization of an anthracene-modified DNA conjugate through bimolecular triplex formation and its analytical application. Org. Biomol. Chem. 2010, 8, 4843–4848. [Google Scholar] [CrossRef]
- Becker, H.D. Unimolecular photochemistry of anthracenes. Chem. Rev. 1993, 93, 145–172. [Google Scholar] [CrossRef]
- Bouas-Laurent, H.; Castellan, A.; Desvergne, J.P.; Lapouyade, R. Photodimerization of anthracenes in fluid solution: Structural aspects. Chem. Soc. Rev. 2000, 29, 43–55. [Google Scholar]
- Pasternak, K.; Pasternak, A.; Gupta, P.; Veedu, R.N.; Wengel, J. Photoligation of self-assembled DNA constructs containing anthracene-functionalized 2'-amino-LNA monomers. Bioorg. Med. Chem. 2011, 19, 7407–7415. [Google Scholar] [CrossRef]
- Lee, H.M.; Larson, D.R.; Lawrence, D.S. Illuminating the chemistry of life: Design, synthesis, and applications of “caged” and related photoresponsive compounds. ACS Chem. Biol. 2009, 4, 409–427. [Google Scholar] [CrossRef]
- Okamoto, A.; Tanabe, K.; Inasaki, T.; Saito, I. Phototriggered drug release from functionalized oligonucleotides by a molecular beacon strategy. Angew. Chem. Int. Edit. 2003, 42, 2502–2504. [Google Scholar] [CrossRef]
- Tanabe, K.; Nakata, H.; Mukai, S.; Nishimoto, S. Modulated drug release from the stem-and-loop structured oligodeoxynucleotide upon UV-A irradiation in the presence of target DNA. Org. Biomol. Chem. 2005, 3, 3893–3897. [Google Scholar] [CrossRef]
- Rothlingshofer, M.; Gorska, K.; Winssinger, N. Nucleic acid-templated energy transfer leading to a photorelease reaction and its application to a system displaying a nonlinear response. J. Am. Chem. Soc. 2011, 133, 18110–18113. [Google Scholar] [CrossRef]
- Clo, E.; Snyder, J.W.; Voigt, N.V.; Ogilby, P.R.; Gothelf, K.V. DNA-programmed control of photosensitized singlet oxygen production. J. Am. Chem. Soc. 2006, 128, 4200–4201. [Google Scholar]
- Clo, E.; Snyder, J.W.; Ogilby, P.R.; Gothelf, K.V. Control and selectivity of photosensitized singlet oxygen production: Challenges in complex biological systems. ChemBioChem 2007, 8, 475–481. [Google Scholar] [CrossRef]
- Jacobsen, M.F.; Clo, E.; Mokhir, A.; Gothelf, K.V. Model systems for activation of nucleic acid encoded prodrugs. ChemMedChem 2007, 2, 793–799. [Google Scholar] [CrossRef]
- Arian, D.; Clo, E.; Gothelf, K.V.; Mokhir, A. A nucleic acid dependent chemical photocatalysis in live human cells. Chem. Eur. J. 2010, 16, 288–295. [Google Scholar] [CrossRef]
- Ma, Z.C.; Taylor, J.S. Nucleic acid-triggered catalytic drug release. Proc. Natl. Acad. Sci. USA 2000, 97, 11159–11163. [Google Scholar]
- Ma, Z.C.; Taylor, J.S. Nucleic acid triggered catalytic drug and probe release: A new concept for the design of chemotherapeutic and diagnostic agents. Bioorg. Med. Chem. 2001, 9, 2501–2510. [Google Scholar] [CrossRef]
- Ma, Z.C.; Taylor, J.S. PNA-based RNA-triggered drug-releasing system. Bioconjug. Chem. 2003, 14, 679–683. [Google Scholar] [CrossRef]
- Cai, J.F.; Li, X.X.; Taylor, J.S. Improved nucleic acid triggered probe activation through the use of a 5-thiomethyluracil peptide nucleic acid building block. Org. Lett. 2005, 7, 751–754. [Google Scholar] [CrossRef]
- Brunner, J.; Mokhir, A.; Kraemer, R. DNA-templated metal catalysis. J. Am. Chem. Soc. 2003, 125, 12410–12411. [Google Scholar] [CrossRef]
- Zelder, F.H.; Brunner, J.; Kramer, R. DNA-templated catalysis using a metal-cleavable linker. Chem. Commun. 2004, 902–903. [Google Scholar]
- Boll, I.; Kramer, R.; Brunner, J.; Mokhir, A. Templated metal catalysis for single nucleotide specific DNA sequence detection. J. Am. Chem. Soc. 2005, 127, 7849–7856. [Google Scholar]
- Gryaznov, S.M.; Letsinger, R.L. Chemical ligation of oligonucleotides in the presence and absence of a template. J. Am. Chem. Soc. 1993, 115, 3808–3809. [Google Scholar] [CrossRef]
- Gryaznov, S.M.; Schultz, R.; Chaturvedi, S.K.; Letsinger, R.L. Enhancement of selectivity in recognition of nucleic acids via chemical autoligation. Nucleic Acids Res. 1994, 22, 2366–2369. [Google Scholar] [CrossRef]
- Herrlein, M.K.; Letsinger, R.L. Selective chemical autoligation on a double-stranded DNA template. Nucleic Acids Res. 1994, 22, 5076–5078. [Google Scholar] [CrossRef]
- Herrlein, M.K.; Nelson, J.S.; Letsinger, R.L. A covalent lock for self-assembled oligonucleotide conjugates. J. Am. Chem. Soc. 1995, 117, 10151–10152. [Google Scholar]
- Xu, Y.Z.; Kool, E.T. High sequence fidelity in a non-enzymatic DNA autoligation reaction. Nucleic Acids Res. 1999, 27, 875–881. [Google Scholar] [CrossRef]
- Xu, Y.Z.; Kool, E.T. Rapid and selective selenium-mediated autoligation of DNA strands. J. Am. Chem. Soc. 2000, 122, 9040–9041. [Google Scholar] [CrossRef]
- Xu, Y.Z.; Karalkar, N.B.; Kool, E.T. Nonenzymatic autoligation in direct three-color detection of RNA and DNA point mutations. Nat. Biotechnol. 2001, 19, 148–152. [Google Scholar] [CrossRef]
- Gartner, Z.J.; Liu, D.R. The generality of DNA-templated synthesis as a basis for evolving non-natural small molecules. J. Am. Chem. Soc. 2001, 123, 6961–6963. [Google Scholar] [CrossRef]
- Abe, H.; Kondo, Y.; Jinmei, H.; Abe, N.; Furukawa, K.; Uchiyama, A.; Tsuneda, S.; Aikawa, K.; Matsumoto, I.; Ito, Y. Rapid DNA chemical ligation for amplification of RNA and DNA signal. Bioconjug. Chem. 2008, 19, 327–333. [Google Scholar] [CrossRef]
- Sando, S.; Kool, E.T. Quencher as leaving group: Efficient detection of DNA-joining reactions. J. Am. Chem. Soc. 2002, 124, 2096–2097. [Google Scholar] [CrossRef]
- Sando, S.; Kool, E.T. Imaging of RNA in bacteria with self-ligating quenched probes. J. Am. Chem. Soc. 2002, 124, 9686–9687. [Google Scholar] [CrossRef]
- Sando, S.; Abe, H.; Kool, E.T. Quenched auto-ligating DNAs: Multicolor identification of nucleic acids at single nucleotide resolution. J. Am. Chem. Soc. 2004, 126, 1081–1087. [Google Scholar] [CrossRef]
- Silverman, A.P.; Kool, E.T. Quenched probes for highly specific detection of cellular RNAs. Trends Biotechnol. 2005, 23, 225–230. [Google Scholar] [CrossRef]
- Abe, H.; Kool, E.T. Destabilizing universal linkers for signal amplification in self-ligating probes for RNA. J. Am. Chem. Soc. 2004, 126, 13980–13986. [Google Scholar]
- Abe, H.; Kool, E.T. Flow cytometric detection of specific RNAs in native human cells with quenched autoligating FRET probes. Proc. Natl. Acad. Sci. USA 2006, 103, 263–268. [Google Scholar] [CrossRef]
- Silverman, A.P.; Baron, E.J.; Kool, E.T. RNA-templated chemistry in cells: Discrimination of Escherichia, Shigella and Salmonella bacterial strains with a new two-color FRET strategy. ChemBioChem 2006, 7, 1890–1894. [Google Scholar] [CrossRef]
- Silverman, A.P.; Kool, E.T. Quenched autoligation probes allow discrimination of live bacterial species by single nucleotide differences in rRNA. Nucleic Acids Res. 2005, 33, 4978–4986. [Google Scholar] [CrossRef]
- Miller, G.P.; Silverman, A.P.; Kool, E.T. New, stronger nucleophiles for nucleic acid-templated chemistry: Synthesis and application in fluorescence detection of cellular RNA. Bioorg. Med. Chem. 2008, 16, 56–64. [Google Scholar] [CrossRef]
- Shibata, A.; Abe, H.; Ito, M.; Kondo, Y.; Shimizu, S.; Aikawa, K.; Ito, Y. DNA templated nucleophilic aromatic substitution reactions for fluorogenic sensing of oligonucleotides. Chem. Commun. 2009, 2009, 6586–6588. [Google Scholar]
- Lemieux, G.A.; de Graffenried, C.L.; Bertozzi, C.R. A fluorogenic dye activated by the Staudinger ligation. J. Am. Chem. Soc. 2003, 125, 4708–4709. [Google Scholar]
- Prescher, J.A.; Bertozzi, C.R. Chemistry in living systems. Nat. Chem. Biol. 2005, 1, 13–21. [Google Scholar]
- El-Sagheer, A.H.; Brown, T. Click chemistry with DNA. Chem. Soc. Rev. 2010, 39, 1388–1405. [Google Scholar]
- Le Droumaguet, C.; Wang, C.; Wang, Q. Fluorogenic click reaction. Chem. Soc. Rev. 2010, 39, 1233–1239. [Google Scholar]
- Jewett, J.C.; Bertozzi, C.R. Cu-free click cycloaddition reactions in chemical biology. Chem. Soc. Rev. 2010, 39, 1272–1279. [Google Scholar]
- Cai, J.F.; Li, X.X.; Yue, X.; Taylor, J.S. Nucleic acid-triggered fluorescent probe activation by the staudinger reaction. J. Am. Chem. Soc. 2004, 126, 16324–16325. [Google Scholar] [CrossRef]
- Pianowski, Z.L.; Winssinger, N. Fluorescence-based detection of single nucleotide permutation in DNA via catalytically templated reaction. Chem. Commun. 2007, 3820–3822. [Google Scholar] [CrossRef]
- Pianowski, Z.; Gorska, K.; Oswald, L.; Merten, C.A.; Winssinger, N. Imaging of mRNA in live cells using nucleic acid-templated reduction of azidorhodamine probes. J. Am. Chem. Soc. 2009, 131, 6492–6497. [Google Scholar]
- Gorska, K.; Keklikoglou, I.; Tschulena, U.; Winssinger, N. Rapid fluorescence imaging of miRNAs in human cells using templated Staudinger reaction. Chem. Sci. 2011, 2, 1969–1975. [Google Scholar] [CrossRef]
- Abe, H.; Wang, J.; Furukawa, K.; Oki, K.; Uda, M.; Tsuneda, S.; Ito, Y. A reduction-triggered fluorescence probe for sensing nucleic acids. Bioconjug. Chem. 2008, 19, 1219–1226. [Google Scholar] [CrossRef]
- Furukawa, K.; Abe, H.; Wang, J.; Uda, M.; Koshino, H.; Tsuneda, S.; Ito, Y. Reduction-triggered red fluorescent probes for dual-color detection of oligonucleotide sequences. Org. Biomol. Chem. 2009, 7, 671–677. [Google Scholar]
- Franzini, R.M.; Kool, E.T. 7-Azidomethoxy-coumarins as profluorophores for templated nucleic acid detection. ChemBioChem 2008, 9, 2981–2988. [Google Scholar] [CrossRef]
- Furukawa, K.; Abe, H.; Hibino, K.; Sako, Y.; Tsuneda, S.; Ito, Y. Reduction-triggered fluorescent amplification probe for the detection of endogenous rnas in living human cells. Bioconjug. Chem. 2009, 20, 1026–1036. [Google Scholar] [CrossRef]
- Shibata, A.; Abe, H.; Furukawa, K.; Tsuneda, S.; Ito, Y. Reduction-triggered fluorescence probe for peptide-templated reactions. Chem. Pharm. Bull. 2009, 57, 1223–1226. [Google Scholar] [CrossRef]
- Furukawa, K.; Abe, H.; Tamura, Y.; Yoshimoto, R.; Yoshida, M.; Tsuneda, S.; Ito, Y. Fluorescence detection of intron lariat RNA with reduction-triggered fluorescent probes. Angew. Chem. Int. Edit 2011, 50, 12020–12023. [Google Scholar] [CrossRef]
- Marras, S.A.E.; Kramer, F.R.; Tyagi, S. Efficiencies of fluorescence resonance energy transfer and contact-mediated quenching in oligonucleotide probes. Nucleic Acids Res. 2002, 30, e122. [Google Scholar] [CrossRef]
- Rothlingshofer, M.; Gorska, K.; Winssinger, N. Nucleic acid templated uncaging of fluorophores using ru-catalyzed photoreduction with visible light. Org. Lett. 2012, 14, 482–485. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Kamlet, A.S.; Steinman, J.B.; Liu, D.R. A biomolecule-compatible visible-light-induced azide reduction from a DNA-encoded reaction-discovery system. Nat. Chem. 2011, 3, 146–153. [Google Scholar]
- Franzini, R.M.; Kool, E.T. Efficient nucleic acid detection by templated reductive quencher release. J. Am. Chem. Soc. 2009, 131, 16021–16023. [Google Scholar] [CrossRef]
- Li, H.; Franzini, R.M.; Bruner, C.; Kool, E.T. Templated chemistry for sequence-specific fluorogenic detection of duplex DNA. ChemBioChem 2010, 11, 2132–2137. [Google Scholar] [CrossRef]
- Franzini, R.M.; Kool, E.T. Two successive reactions on a DNA template: A strategy for improving background fluorescence and specificity in nucleic acid detection. Chem. Eur. J. 2011, 17, 2168–2175. [Google Scholar] [CrossRef]
- Franzini, R.M.; Kool, E.T. Improved templated fluorogenic probes enhance the analysis of closely related pathogenic bacteria by microscopy and flow cytometry. Bioconjugate Chem. 2011, 22, 1869–1877. [Google Scholar] [CrossRef]
- Gorska, K.; Manicardi, A.; Barluenga, S.; Winssinger, N. DNA-templated release of functional molecules with an azide-reduction-triggered immolative linker. Chem. Commun. 2011, 47, 4364–4366. [Google Scholar]
- Ficht, S.; Mattes, A.; Seitz, O. Single-nucleotide-specific PNA-peptide ligation on synthetic and PCR DNA templates. J. Am. Chem. Soc. 2004, 126, 9970–9981. [Google Scholar] [CrossRef]
- Ficht, S.; Dose, C.; Seitz, O. As fast and selective as enzymatic ligations: Unpaired nucleobases increase the selectivity of DNA-controlled native chemical PNA ligation. ChemBioChem 2005, 6, 2098–2103. [Google Scholar] [CrossRef]
- Dose, C.; Ficht, S.; Seitz, O. Reducing product inhibition in DNA-template-controlled ligation reactions. Angew. Chem. Int. Ed. 2006, 45, 5369–5373. [Google Scholar] [CrossRef]
- Dose, C.; Seitz, O. Single nucleotide specific detection of DNA by native chemical ligation of fluorescence labeled PNA-probes. Bioorg. Med. Chem. 2008, 16, 65–77. [Google Scholar] [CrossRef]
- Grossmann, T.N.; Seitz, O. DNA-catalyzed transfer of a reporter group. J. Am. Chem. Soc. 2006, 128, 15596–15597. [Google Scholar] [CrossRef]
- Grossmann, T.N.; Seitz, O. Nucleic acid templated reactions: Consequences of probe reactivity and readout strategy for amplified signaling and sequence selectivity. Chem. Eur. J. 2009, 15, 6723–6730. [Google Scholar] [CrossRef]
- Grossmann, T.N.; Roglin, L.; Seitz, O. Target-catalyzed transfer reactions for the amplified detection of RNA. Angew. Chem. Int. Edit. 2008, 47, 7119–7122. [Google Scholar] [CrossRef]
- Erben, A.; Grossmann, T.N.; Seitz, O. DNA-triggered synthesis and bioactivity of proapoptotic peptides. Angew. Chem. Int. Edit. 2011, 50, 2828–2832. [Google Scholar] [CrossRef]
- Erben, A.; Grossmann, T.N.; Seitz, O. DNA-instructed acyl transfer reactions for the synthesis of bioactive peptides. Bioorg. Med. Chem. Lett. 2011, 21, 4993–4997. [Google Scholar]
- Franzini, R.M.; Kool, E.T. Organometallic activation of a fluorogen for templated nucleic acid detection. Org. Lett. 2008, 10, 2935–2938. [Google Scholar] [CrossRef]
- Meguellati, K.; Koripelly, G.; Ladame, S. DNA-templated synthesis of trimethine cyanine dyes: A versatile fluorogenic reaction for sensing G-quadruplex formation. Angew. Chem. Int. Edit. 2010, 49, 2738–2742. [Google Scholar] [CrossRef]
- Koripelly, G.; Meguellati, K.; Ladame, S. Dual sensing of hairpin and quadruplex DNA structures using multicolored peptide nucleic acid fluorescent probes. Bioconjug. Chem. 2010, 21, 2103–2109. [Google Scholar] [CrossRef]
- Gartner, Z.J.; Grubina, R.; Calderone, C.T.; Liu, D.R. Two enabling architectures for DNA-templated organic synthesis. Angew. Chem. Int. Edit. 2003, 42, 1370–1375. [Google Scholar] [CrossRef]
- Calderone, C.T.; Liu, D.R. Nucleic-acid-templated synthesis as a model system for ancient translation. Curr. Opin. Chem. Biol. 2004, 8, 645–653. [Google Scholar] [CrossRef]
- Li, X.Y.; Liu, D.R. DNA-Templated organic synthesis: Nature’s strategy for controlling chemical reactivity applied to synthetic molecules. Angew. Chem. Int. Edit. 2004, 43, 4848–4870. [Google Scholar] [CrossRef]
- Sando, S.; Sasaki, T.; Kanatani, K.; Aoyama, Y. Amplified nucleic acid sensing using programmed self-cleaving DNAzyme. J. Am. Chem. Soc. 2003, 125, 15720–15721. [Google Scholar]
- Sando, S.; Narita, A.; Sasaki, T.; Aoyama, Y. . Locked TASC probes for homogeneous sensing of nucleic acids and imaging of fixed E-coli cells. Org. Biomol. Chem. 2005, 3, 1002–1007. [Google Scholar] [CrossRef]
- Obika, S.; Tomizu, M.; Negoro, Y.; Orita, A.; Nakagawa, O.; Imanishi, T. Double-stranded DNA-templated oligonucleotide digestion triggered by triplex formation. ChemBioChem 2007, 8, 1924–1928. [Google Scholar] [CrossRef]
- Ito, K.R.; Kodama, T.; Tomizu, M.; Negoro, Y.; Orita, A.; Osaki, T.; Hosoki, N.; Tanaka, T.; Imanishi, T.; Obika, S. Double-stranded DNA-templated cleavage of oligonucleotides containing a P3'→N5' linkage triggered by triplex formation: The effects of chemical modifications and remarkable enhancement in reactivity. Nucleic Acids Res. 2010, 38, 7332–7342. [Google Scholar]
- Ito, K.R.; Kodama, T.; Makimura, F.; Hosoki, N.; Osaki, T.; Orita, A.; Imanishi, T.; Obika, S. Cleavage of oligonucleotides containing a P3'→N5' phosphoramidate linkage mediated by single-stranded oligonucleotide templates. Molecules 2011, 16, 10695–10708. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shibata, A.; Abe, H.; Ito, Y. Oligonucleotide-Templated Reactions for Sensing Nucleic Acids. Molecules 2012, 17, 2446-2463. https://doi.org/10.3390/molecules17032446
Shibata A, Abe H, Ito Y. Oligonucleotide-Templated Reactions for Sensing Nucleic Acids. Molecules. 2012; 17(3):2446-2463. https://doi.org/10.3390/molecules17032446
Chicago/Turabian StyleShibata, Aya, Hiroshi Abe, and Yoshihiro Ito. 2012. "Oligonucleotide-Templated Reactions for Sensing Nucleic Acids" Molecules 17, no. 3: 2446-2463. https://doi.org/10.3390/molecules17032446