The Coumarin Psoralidin Enhances Anticancer Effect of Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand (TRAIL)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Results and Discussion
2.1. Cytotoxic and Apoptotic Activities of Psoralidin in HeLa Cancer Cells

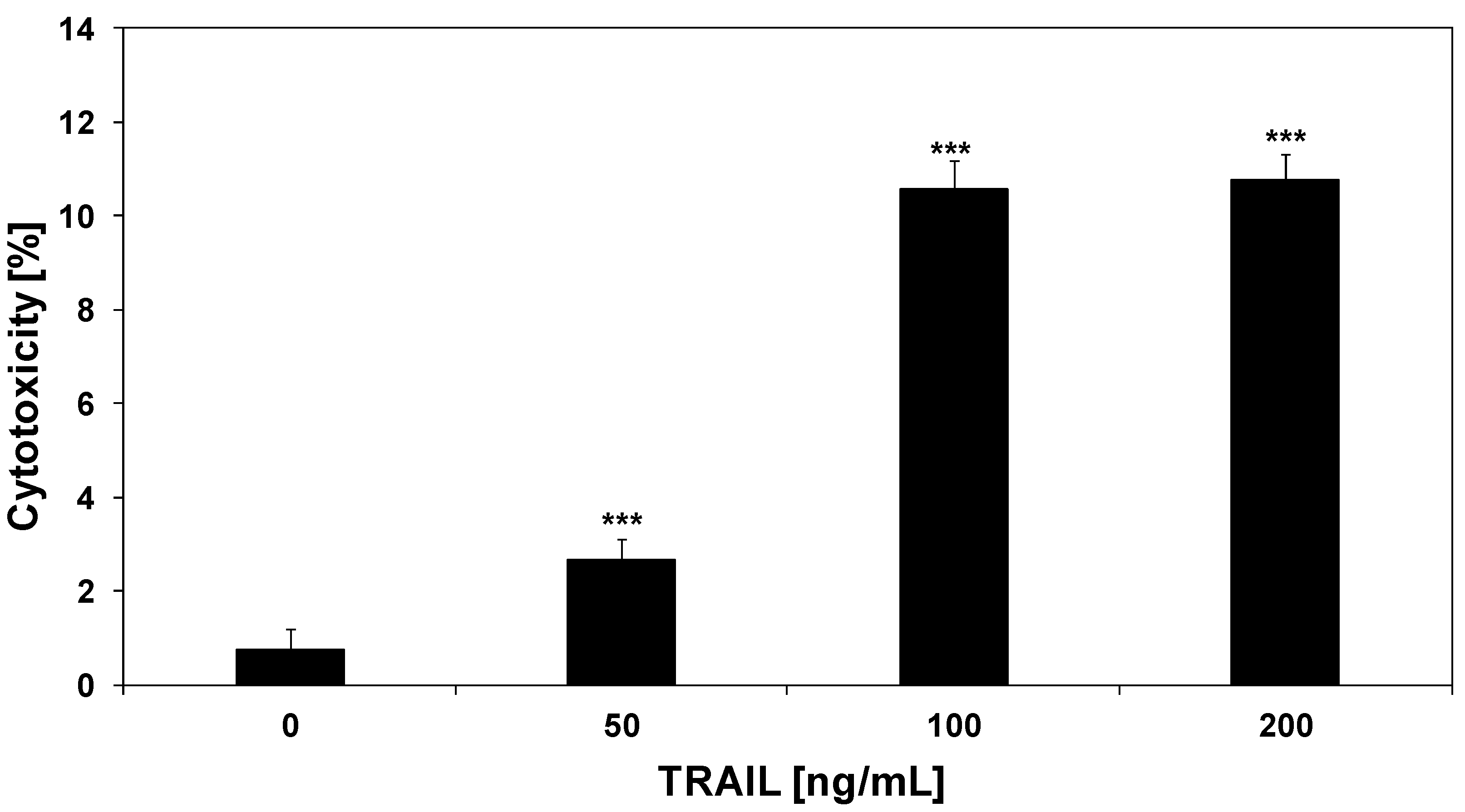
2.2. Cytotoxic and Apoptotic Activities of TRAIL in HeLa Cancer Cells

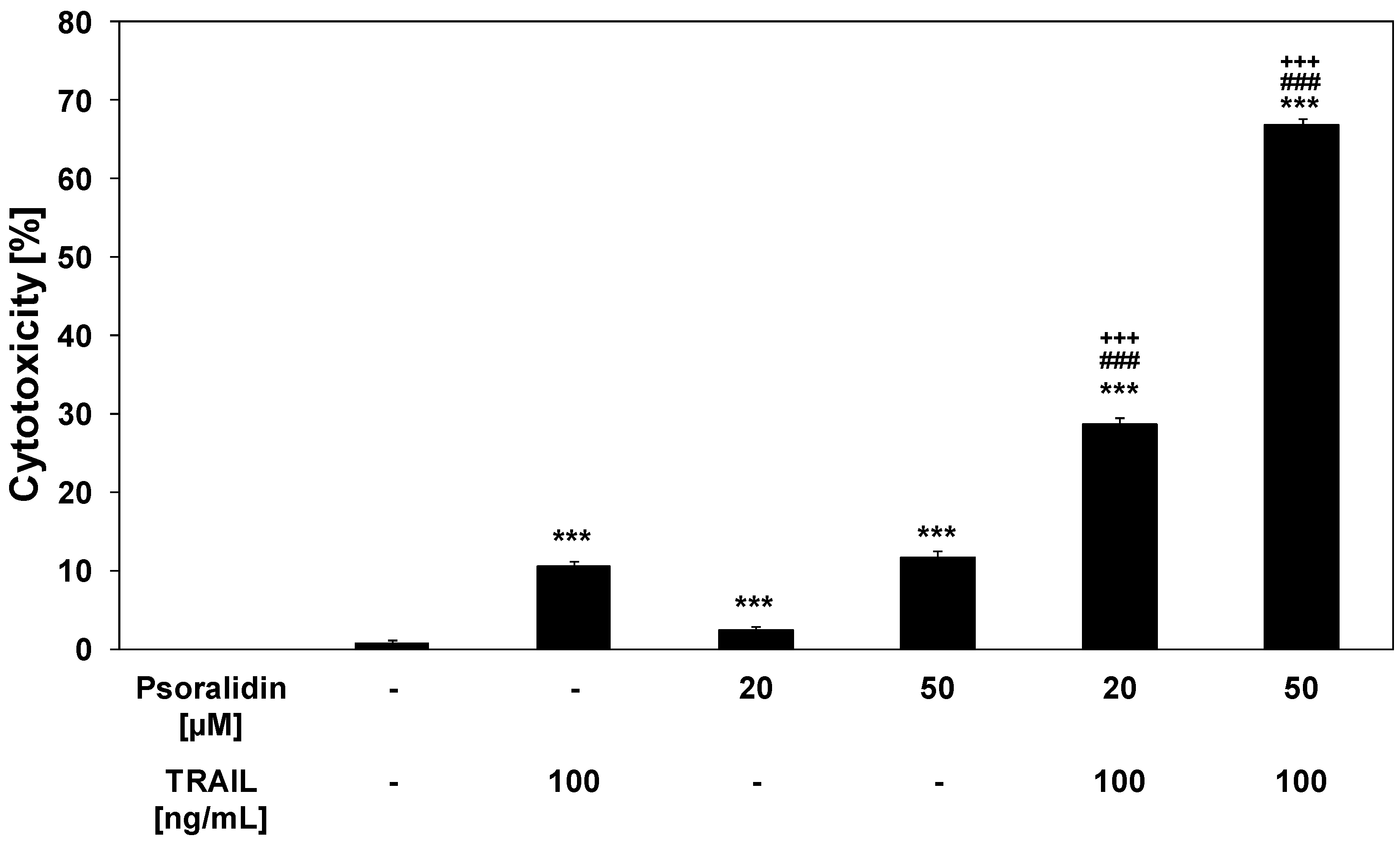
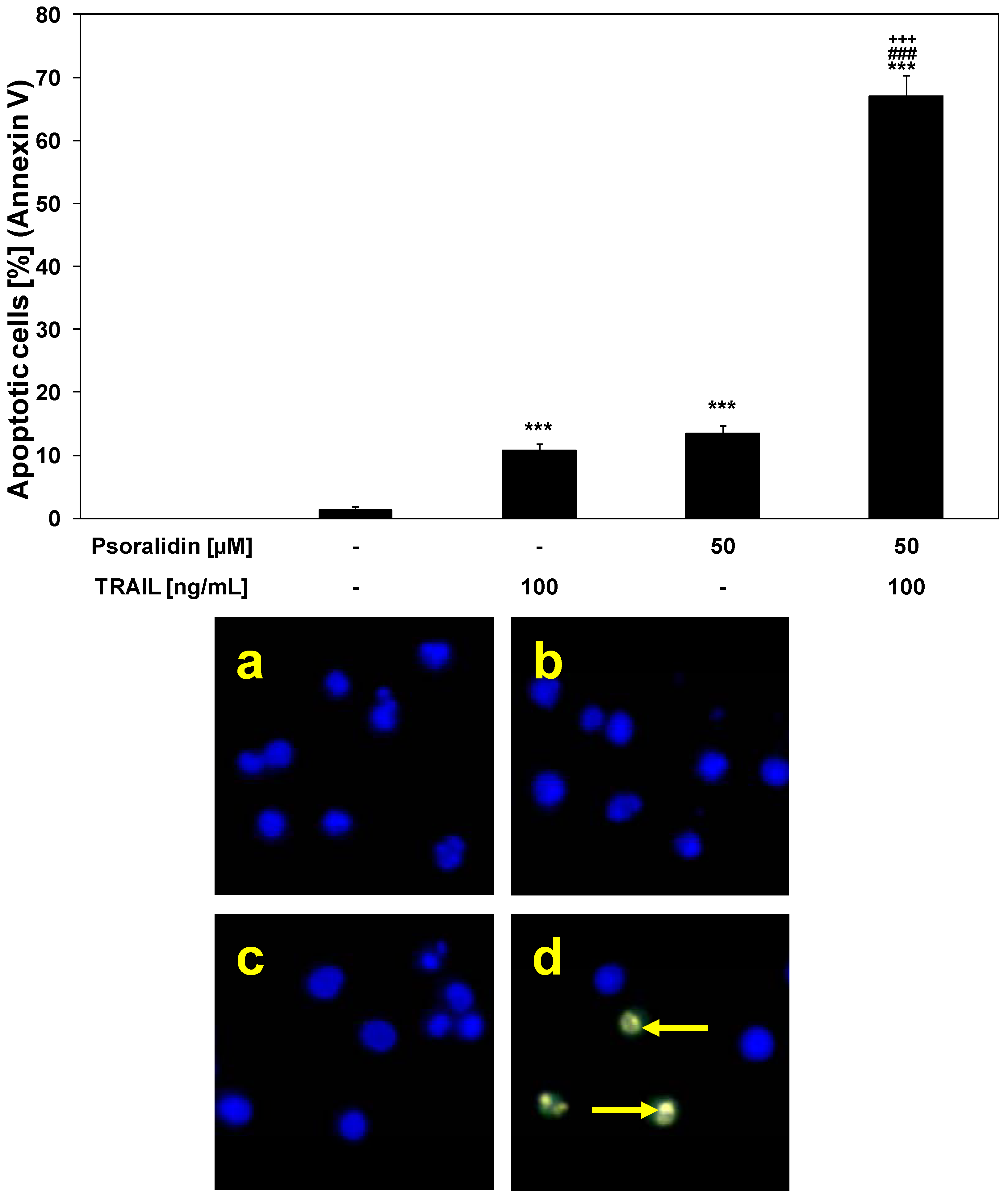
2.3. Cytotoxic and Apoptotic Activities of TRAIL in Combination with Psoralidin in HeLa Cancer Cells


2.4. The Mechanism by Which Psoralidin Sensitize HeLa Cancer Cells to TRAIL-Induced Apoptosis
2.4.1. Effects of Psoralidin on Death Receptor Expression in HeLa Cancer Cells

2.4.2. Effects of TRAIL and Psoralidin on Mitochondrial Membrane Potential (ΔΨm) in HeLa Cancer Cells

3. Experimental
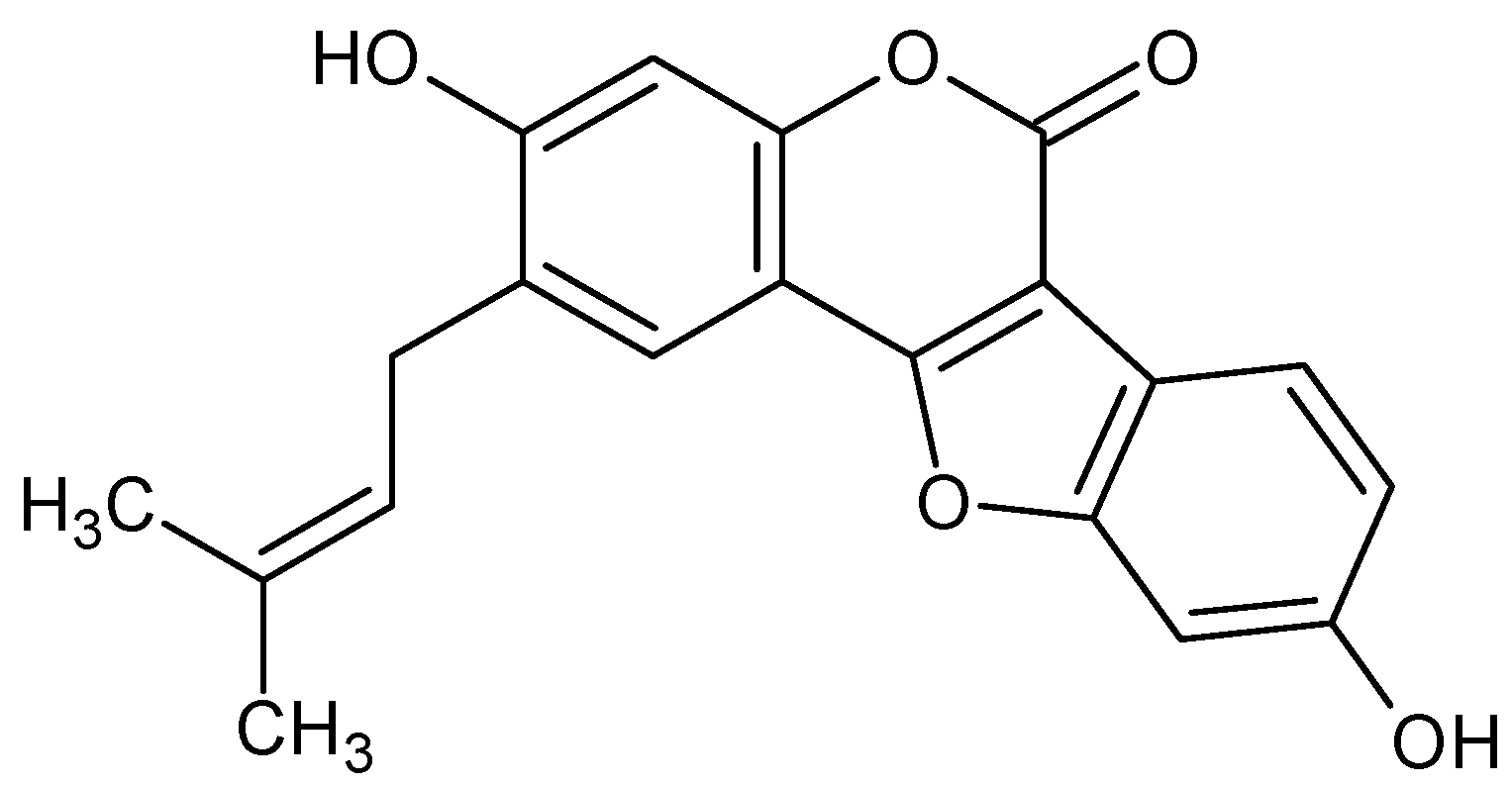
3.1. Coumarin
3.2. TRAIL
3.3. Cancer Cell Culture
3.4. Cytotoxicity Assay
3.5. Lactate Dehydrogenase Release Assay
3.6. Determination of Apoptotic Cell Death by Fluorescence Microscopy with Annexin V-FITC Staining
3.7. Flow Cytometric Analysis of Death Receptor Expression on the Cancer Cell Surface
3.8. Evaluation of Mitochondrial Membrane Potential by DePsipher
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
- Sample Availability: Samples of the compound psoralidin are available from the Alexis Biochemicals (San Diego, CA, USA).
References and Notes
- Almeida, I.M.; Barreira, J.C.; Oliveira, M.B.; Ferreira, I.C. Dietary antioxidant supplements: Benefits of their combined use. Food Chem. Toxicol. 2011, 49, 3232–3237. [Google Scholar] [CrossRef]
- Barros, L.; Duenas, M.; Carvalho, A.M.; Ferreira, I.C.; Santos-Buelga, C. Characterization of phenolic compounds in flowers of wild medicinal plants from Northeastern Portugal. Food Chem. Toxicol. 2012, 50, 1576–1582. [Google Scholar] [CrossRef]
- Pinela, J.; Barros, L.; Carvalho, A.M.; Ferreira, I.C. Nutritional composition and antioxidant activity of four tomato (Lycopersicon esculentum L.) farmer’ varieties in Northeastern Portugal home gardens. Food Chem. Toxicol. 2012, 50, 829–834. [Google Scholar]
- Wu, L.; Wang, X.; Xu, W.; Farzaneh, F.; Xu, R. The structure and pharmacological functions of coumarins and their derivatives. Curr. Med. Chem. 2009, 16, 4236–4260. [Google Scholar] [CrossRef]
- Riveiro, M.E.; De Kimpe, N.; Moglioni, A.; Vázquez, R.; Monczor, F.; Shayo, C.; Davio, C. Coumarins: Old compounds with novel promising therapeutic perspectives. Curr. Med. Chem. 2010, 17, 1325–1338. [Google Scholar] [CrossRef]
- Zhao, L.H.; Huang, C.Y.; Shan, Z.; Xiang, B.G.; Mei, L.H. Fingerprint analysis of Psoralea corylifolia by HLPC and LC-MS. J. Chromatogr. B 2005, 821, 67–74. [Google Scholar] [CrossRef]
- Xiao, D.; Li, G.; Chen, L.; Zhang, Z.; Yin, J.; Wu, T.; Cheng, Z.; Wei, X.; Wang, Z. Isolation of antioxidants from Psoraleacorylifolia fruits using high-speed counter-current chromatography guided by thin layer chromatography-antioxidant autographic assay. J. Chromatogr. A 2010, 1217, 5470–5476. [Google Scholar] [CrossRef]
- Yang, Y.M.; Hyun, J.W.; Sung, M.S.; Chung, H.S., Kim; Paik, W.H.; Kang, S.S.; Park, J.G. The cytotoxicity of psoralidin from Psoralea corylifolia. Planta Med. 1996, 62, 353–354. [Google Scholar] [CrossRef]
- Mar, W.; Je, K.H.; Seo, E.K. Cytotoxic constituents of Psoralea corylifolia. Arch. Pharm. Res. 2001, 24, 211–213. [Google Scholar] [CrossRef]
- Pahari, P.; Rohr, J. Total synthesis of psoralidin, an anticancer natural product. J. Org. Chem. 2009, 74, 2750–2754. [Google Scholar] [CrossRef]
- Srinivasan, S.; Kumar, R.; Koduru, S.; Chandramouli, A.; Damodaran, C. Inhibiting TNF-mediated signaling: A novel therapeutic paradigm for androgen independent prostate cancer. Apoptosis 2010, 15, 153–161. [Google Scholar] [CrossRef]
- Kumar, R.; Srinivasan, S.; Pahari, P.; Rohr, J.; Damodaran, C. Activating stress-activated protein kinase-mediated cell death and inhibiting epidermal growth factor receptor signaling: A promising therapeutic strategy for prostate cancer. Mol. Cancer Ther. 2010, 9, 2488–2496. [Google Scholar] [CrossRef]
- Krenn, L.; Paper, D.H. Inhibition of angiogenesis and inflammation by an extract of red clover (Trifolium pratense L.). Phytomedicine 2009, 16, 1083–1088. [Google Scholar]
- Ozmen, A.; Bauer, S.; Gridling, M.; Singhuber, J.; Krasteva, S.; Madlener, S.; Vo, T.P.; Stark, N.; Saiko, P.; Fritzer-Szekeres, M.; et al. In vitro anti-neoplastic activity of the ethno-pharmaceutical plant Hypericum adenotrichum Spach endemic to Western Turkey. Oncol. Rep. 2009, 22, 845–52. [Google Scholar]
- Ozmen, A.; Madlener, S.; Bauer, S.; Krasteva, S.; Vonach, C.; Giessrigl, B.; Gridling, M.; Viola, K.; Stark, N.; Saiko, P.; et al. In vitro anti-leukemic activity of the ethno-pharmacological plant Scutellaria orientalis ssp. carica endemic to western Turkey. Phytomedicine 2010, 17, 55–62. [Google Scholar] [CrossRef]
- Szliszka, E.; Zydowicz, G.; Janoszka, B.; Dobosz, C.; Kowalczyk-Ziomek, G.; Krol, W. Ethanolic extract of Brazilian green propolis sensitizes prostate cancer cells to TRAIL-induced apoptosis. Int. J. Oncol. 2011, 38, 941–953. [Google Scholar]
- Szliszka, E.; Krol, W. Soy isoflavones augment the effect of TRAIL-mediated apoptotic death in prostate cancer cells. Oncol. Rep. 2011, 26, 533–541. [Google Scholar]
- Szliszka, E.; Krol, W. The role of dietary polyphenols in tumor necrosis factor-related apoptosis inducing ligand (TRAIL)-induced apoptosis for cancer chemoprevention. Eur. J. Cancer Prev. 2011, 20, 63–69. [Google Scholar] [CrossRef]
- Vanamala, J.; Leonardi, T.; Patil, B.S.; Taddeo, S.S.; Murphy, M.E.; Pike, L.M.; Chapkin, RS.; Lupton, J.R.; Turner, N.D. Suppression of colon carcinogenesis by bioactive compounds in grapefruit. Carcinogenesis 2006, 27, 1257–1265. [Google Scholar] [CrossRef]
- Turner, N.D.; Paulhill, K.J.; Warren, C.A.; Davidson, L.A.; Chapkin, R.S.; Lupton, J.R.; Carroll, R.J.; Wang, N. Quercetin suppresses early colon carcinogenesis partly through inhibition of inflammatory mediators. Acta Hortic. 2009, 841, 237–242. [Google Scholar]
- Warren, C.A.; Paulhill, K.J.; Davidson, L.A.; Lupton, J.R.; Taddeo, S.S.; Hong, M.Y.; Carroll, R.J.; Chapkin, R.S.; Turner, N.D. Quercetin may suppress rat aberrant crypt foci formation by suppressing inflammatory mediators that influence proliferation and apoptosis. J. Nutr. 2009, 139, 101–105. [Google Scholar]
- Leonardi, T.; Vanamala, J.; Taddeo, S.S.; Davidson, L.A.; Murphy, M.E.; Patil, B.S.; Wang, N.; Carroll, R.J.; Chapkin, R.S.; Lupton, J.R.; et al. Apigenin and naringenin suppress colon carcinogenesis through the aberrant crypt stage in azoxymethane-treated rats. Exp. Biol. Med. 2010, 235, 710–717. [Google Scholar]
- Lee, J.Y.; Huerta-Yepez, S.; Vega, M.; Baritaki, S.; Spandidos, D.A.; Bonavida, B. The NO TRAIL to YES TRAIL in cancer therapy (review). Int. J. Oncol. 2007, 31, 685–691. [Google Scholar]
- Szliszka, E.; Helewski, K.J.; Mizgala, E.; Krol, W. The dietary flavonolfisetin enhances the apoptosis-inducing potential of TRAIL in prostate cancer cells. Int. J. Oncol. 2011, 39, 771–779. [Google Scholar]
- Horinanka, M.; Yoshida, T.; Shiraishi, T.; Nakata, S.; Wakada, M.; Nakanishi, R.; Nishino, H.; Sakai, T. The combination of TRAIL and luteolin enhances apoptosis in human cervival cancer HeLa cells. Biochem. Biophys. Res. Comun. 2005, 333, 833–838. [Google Scholar] [CrossRef]
- Szliszka, E.; Mazur, B.; Zydowicz, G.; Czuba, Z.P.; Krol, W. TRAIL-induced apoptosis and expression of death receptor TRAIL-R1 and TRAIL-R2 in bladder cancer cells. Folia Histochem. Cytobiol. 2009, 47, 579–585. [Google Scholar] [CrossRef]
- Szliszka, E.; Czuba, Z.P.; Sedek, L.; Paradysz, A.; Krol, W. Enhanced TRAIL-mediated apoptosis in prostate cancer cells by the bioactive compounds neobavaisoflavone and psoralidin isolated from Psoralea corylifolia. Pharmacol. Rep. 2011, 63, 139–148. [Google Scholar]
- Latha, P.G.; Evans, D.A.; Panikkar, K.R.; Jayavardhanan, K.K. Immunomodulatory and antitumour properties of Psoralea corylifolia seeds. Fitoterapia 2000, 71, 223–231. [Google Scholar] [CrossRef]
- Khatune, N.A.; Islam, M.E.; Haque, M.E.; Khondkar, P.; Rahman, M.M. Antibacterial compounds from seed of Psoralea corylifolia. Fitoterapia 2004, 75, 228–230. [Google Scholar] [CrossRef]
- Ruan, B.; Kong, L.Y.; Takaya, Y.; Niwa, M. Studies on the chemical constituents of Psoralea corylifolia L. J. Asian Nat. Prod. Res. 2007, 9, 41–44. [Google Scholar] [CrossRef]
- Lee, S.; Nam, K.; Mar, W. Induction of quinonereductase activity by psoralidin isolated from Psoralea corylifolia in mouse Hepa 1c1c7 cells. Arch. Pharm. Res. 2009, 32, 1061–1065. [Google Scholar] [CrossRef]
- 32. Szliszka, E.; Skaba, D.; Czuba, Z.P.; Krol, W. Inhibition of inflammatory mediators by neobavaisoflavone in activated RAW264.7 macrophages. Molecules 2011, 16, 3701–3712. [Google Scholar] [CrossRef]
- Chiou, W.; Don, M.; Liao, J., Wei. Psoralidin inhibits LPS-induced iNOS expression via repressing Syk-mediated activation of P13K-IKK-Iκβ signaling pathways. Eur. J. Pharmacol. 2011, 650, 102–109. [Google Scholar]
- Szliszka, E.; Czuba, Z.P.; Jernas, K.; Krol, W. Dietary flavonoids sensitize HeLa cells to tumor necrosis factor-related apoptosis-inducing ligand (TRAIL). Int. J. Mol. Sci. 2008, 9, 56–64. [Google Scholar] [CrossRef]
- Szliszka, E.; Czuba, Z.P.; Domino, M.; Mazur, B.; Zydowicz, G.; Krol, W. Ethanolic extract of propolis (EEP) enhances the apoptosis-inducing potential of TRAIL in cancer cells. Molecules 2009, 14, 738–754. [Google Scholar] [CrossRef]
- Bronikowska, J.; Szliszka, E.; Czuba, Z.P.; Zwolinski, D.; Szmydki, B.; Krol, W. The combination of TRAIL and isoflavones enhances apoptosis in cancer cells. Molecules 2010, 15, 2000–2015. [Google Scholar] [CrossRef]
- Szliszka, E.; Majcher, A.; Domino, M.; Pietsz, G.; Krol, W. Cytotoxic activity of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) against bladder cancer cells after using chemotherapeutic drugs. Urol. Pol. 2007, 60, 138–142. [Google Scholar]
- Szliszka, E.; Bronikowska, J.; Majcher, A.; Miszkiewicz, J.; Krol, W. Enhanced sensitivity of hormone-refractory prostate cancer cells to tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) mediated cytotoxicity by taxanes. CEJ Urol. 2009, 62, 29–34. [Google Scholar]
- Szliszka, E.; Gebka, J.; Bronikowska, J.; Krol, W. Dietary flavones enhance the effect of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) on bladder cancer cells. CEJ Urol. 2010, 63, 138–143. [Google Scholar]
- Szliszka, E.; Czuba, Z.P.; Kawczyk-Krupka, A.; Sieron-Stoltny, A.; Sieron, A.; Krol, W. Chlorin-based photodynamic therapy enhances the effect of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) in bladder cancer cells. Med. Sci. Monit. 2012, 18, BR47–BR53. [Google Scholar]
- Webb, J.L. Effect of more than One Inhibitor. In Enzyme and Metabolic Inhibitors, 1st ed; Academic Press: New York, NY, USA, 1963; Volume 1, pp. 487–512. [Google Scholar]
- Kok, S.H.; Yeh, C.C.; Chen, M.L.; Kuo, M.Y. Esculetin enhances TRAIL-induced apoptosis through DR5 upregulation in human oral cancer SAS cells. Oral Oncol. 2009, 45, 1067–1072. [Google Scholar] [CrossRef]
- Chuang, J.; Huang, Y.; Lu, H.; Ho, H.; Yang, J.; Li, T.; Chang, N.; Chung, J. Coumarin induces cell cycle arrest and apoptosis in human cervical cancer HeLa cells through a mitochondria- and caspase-3 dependent mechanism and NF-κB down-regulation. In Vivo 2007, 21, 1003–1010. [Google Scholar]
- Goel, A.; Prasad, A.K.; Parmar, V.S.; Ghosh, B.; Saini, N. Apoptogenic effect of 7,8-diacetoxy-4-methylcoumarin and 7,8-diacetoxy-4-methylthiocoumarin in human lung adenocarcinoma cell line: Role of NF-κB, Akt, ROS and MAP kinase pathway. Chem. Biol. Interact. 2009, 179, 363–374. [Google Scholar] [CrossRef]
- Yang, J.; Xiao, Y.; He, X.; Qiu, G.; Hu, X. Aesculetin-induced apoptosis through a ROS-mediated mitochondrial dysfunction pathway in human cervical cancer cells. J. Asian Nat. Prod. Res. 2010, 12, 185–193. [Google Scholar] [CrossRef]
- Bhattacharyya, S.S.; Paul, S.; De, A.; Das, D.; Samadder, A.; Boujedaini, N.; Khuda-Bukhsh, A.R. Poly (lactide-co-glycolide) acid nanoencapsulation of synthetic coumarin: Cytotoxicity and bio-distribution in mice, in cancer cell line and interaction with calf thymus DNA as target. Toxicol. Appl. Pharmacol. 2011, 253, 270–281. [Google Scholar] [CrossRef]
- Szliszka, E.; Czuba, Z.P.; Mertas, A.; Paradysz, A.; Krol, W. The dietary isoflavone biochanin-A sensitizes prostate cancer cells to TRAIL-induced apoptosis. Urol. Oncol. 2011. [Google Scholar] [CrossRef]
- Stankovic, M.S.; Curcic, M.G.; Zizic, J.B.; Topuzovic, M.D.; Solujic, S.R.; Markovic, S.D. Teucrium plant species as natural sources of novel anticancer compounds: Antiproliferative, proapoptotic and antioxidant properties. Int. J. Mol. Sci. 2011, 12, 4190–4205. [Google Scholar] [CrossRef]
- Cheah, S.; Appleton, D.R.; Lee, S.; Lam, M.; Hamid, A.; Hadi, A.; Mustafa, M.R. Panduratin A inhibits the growth of A549 cells through induction of apoptosis and inhibiotion of NF-KappaB translocation. Molecules 2011, 16, 2583–2598. [Google Scholar] [CrossRef]
- Szliszka, E.; Czuba, Z.P.; Mazur, B.; Sedek, L.; Paradysz, A.; Krol, W. Chalcones enhance TRAIL-induced apoptosis in prostate cancer cells. Int. J. Mol. Sci. 2010, 11, 1–13. [Google Scholar]
- Szliszka, E.; Czuba, Z.P.; Mazur, B.; Paradysz, A.; Krol, W. Chalcones and dihydrochalcones augment TRAIL-mediated apoptosis in prostate cancer cells. Molecules 2010, 15, 5336–5353. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bronikowska, J.; Szliszka, E.; Jaworska, D.; Czuba, Z.P.; Krol, W. The Coumarin Psoralidin Enhances Anticancer Effect of Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand (TRAIL). Molecules 2012, 17, 6449-6464. https://doi.org/10.3390/molecules17066449
Bronikowska J, Szliszka E, Jaworska D, Czuba ZP, Krol W. The Coumarin Psoralidin Enhances Anticancer Effect of Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand (TRAIL). Molecules. 2012; 17(6):6449-6464. https://doi.org/10.3390/molecules17066449
Chicago/Turabian StyleBronikowska, Joanna, Ewelina Szliszka, Dagmara Jaworska, Zenon P. Czuba, and Wojciech Krol. 2012. "The Coumarin Psoralidin Enhances Anticancer Effect of Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand (TRAIL)" Molecules 17, no. 6: 6449-6464. https://doi.org/10.3390/molecules17066449