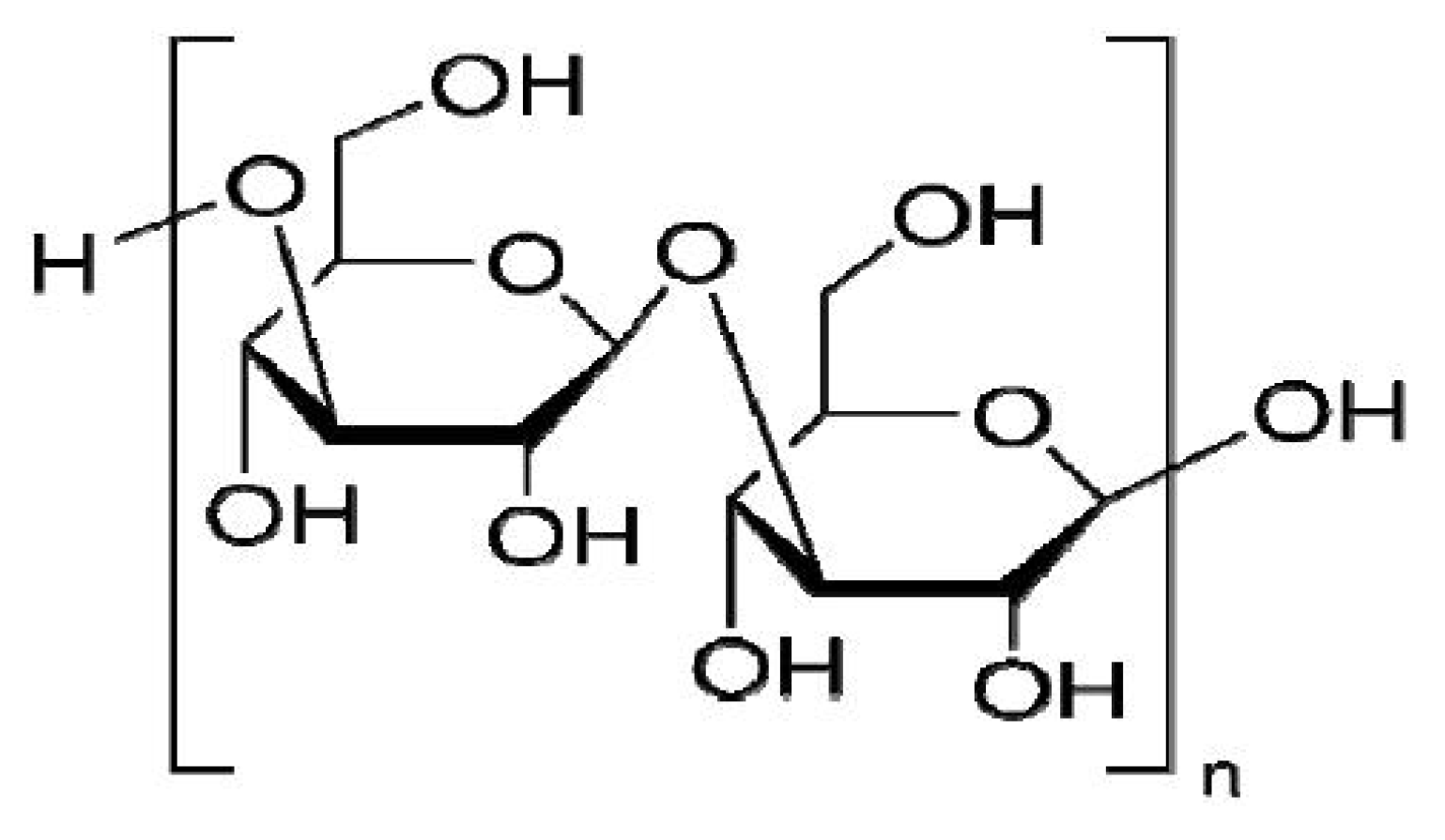
Laminarin Induces Apoptosis of Human Colon Cancer LOVO Cells through a Mitochondrial Pathway

Abstract
:1. Introduction
2. Results and Discussion
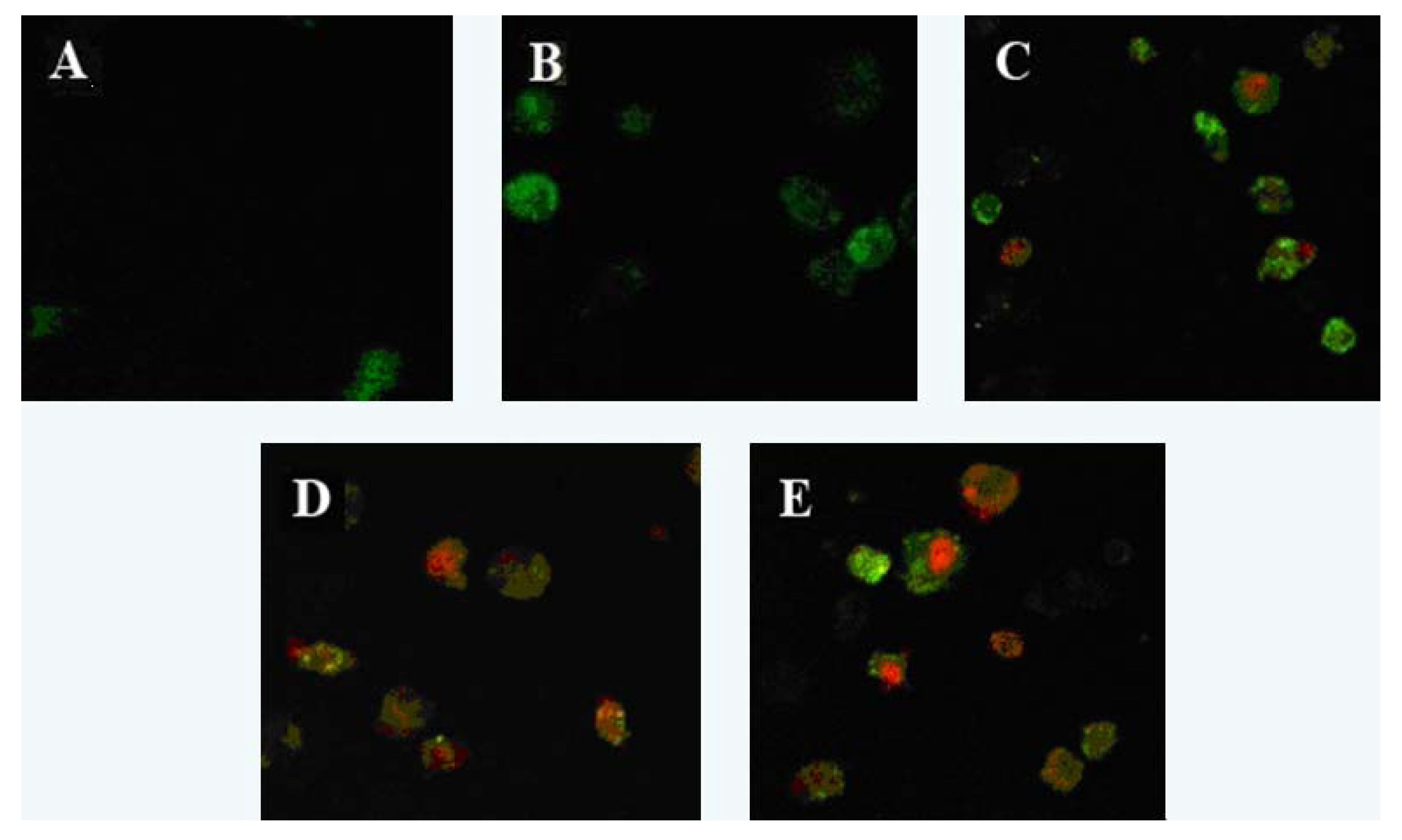
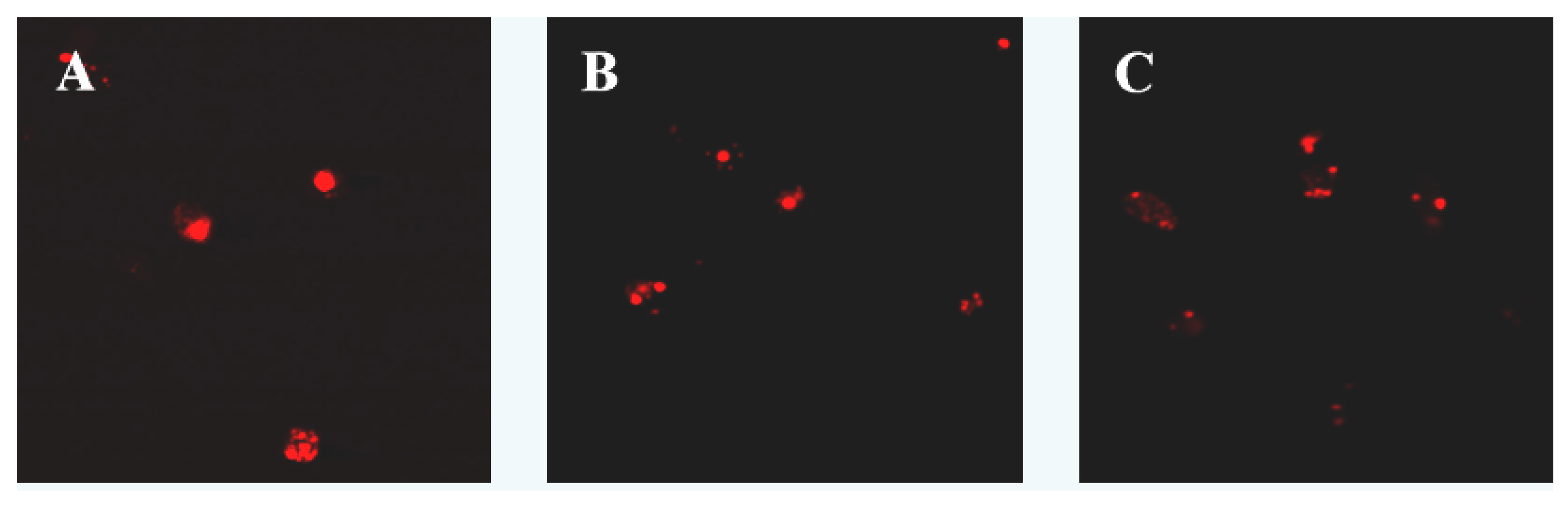
2.1. Apoptosis Morphology Observation
2.2. Effect of Laminarin on Reactive Oxygen Species (ROS), Calcium Ion Concentration (Ca2+) and pH Value in LOVO Cells
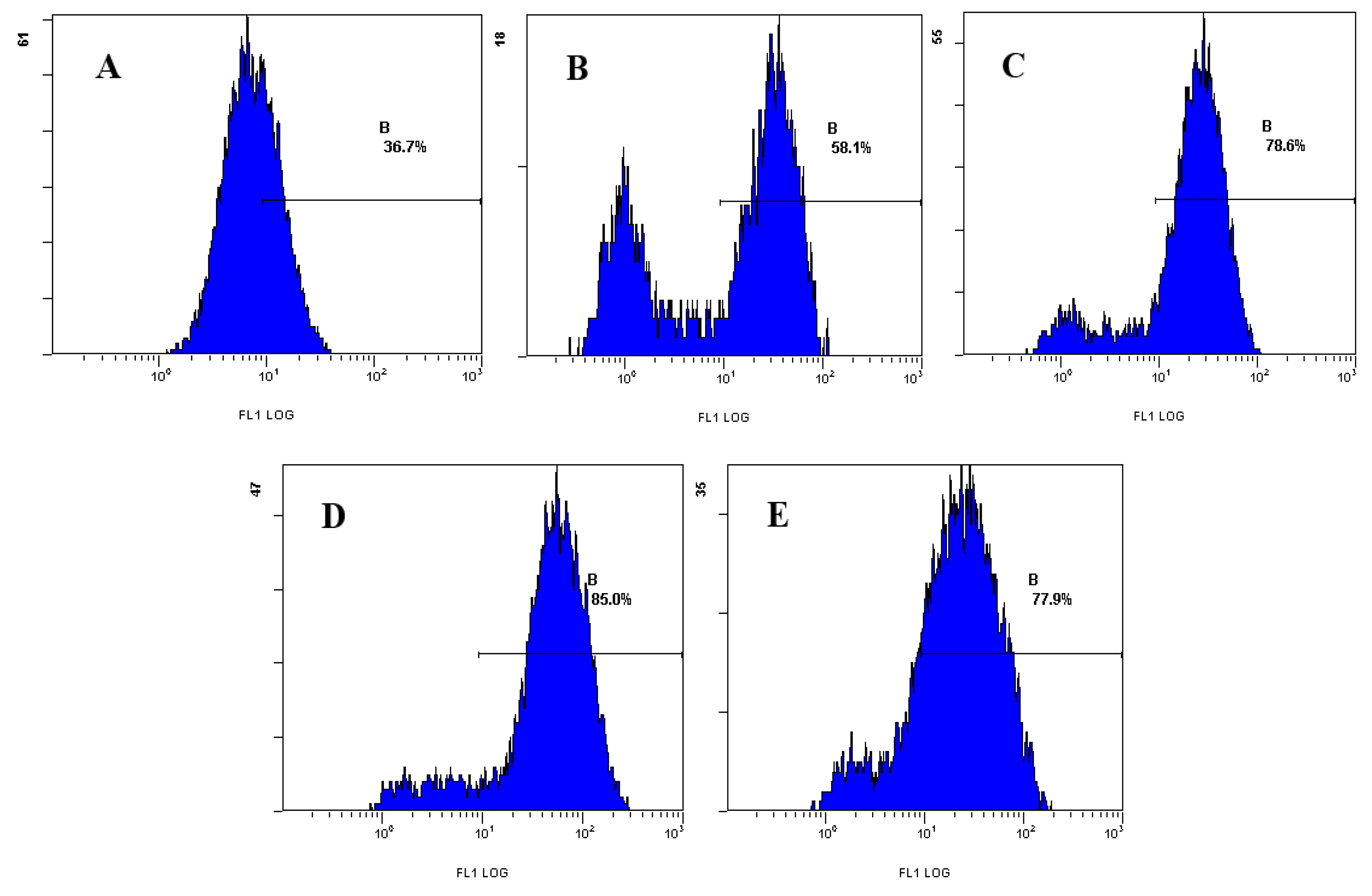
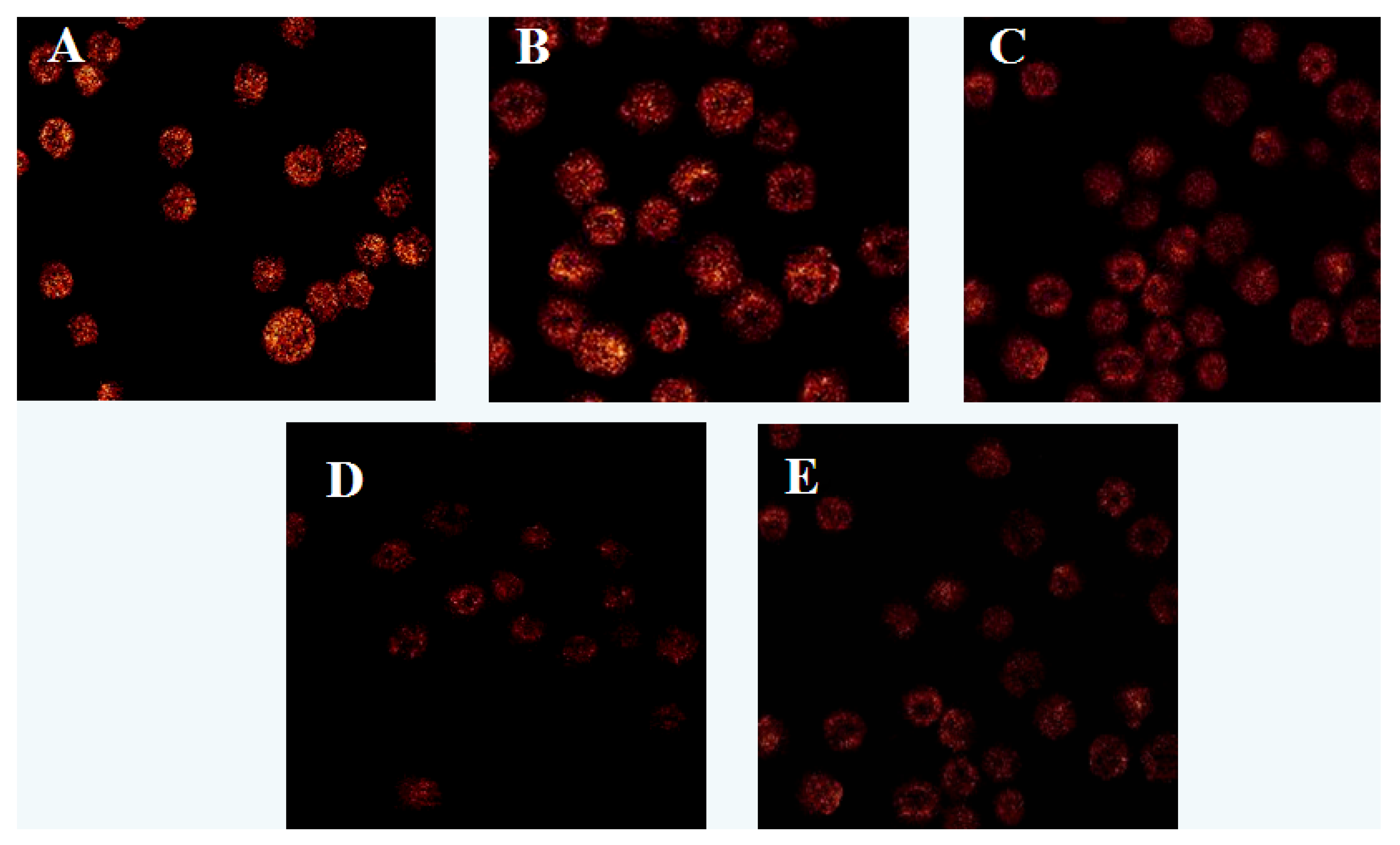
2.2.1. Effect of Laminarin on ROS in LOVO Cells
2.2.2. Effect of Laminarin on Ca2+ in LOVO Cells
2.2.3. Effect of Laminarin on pH in LOVO Cells
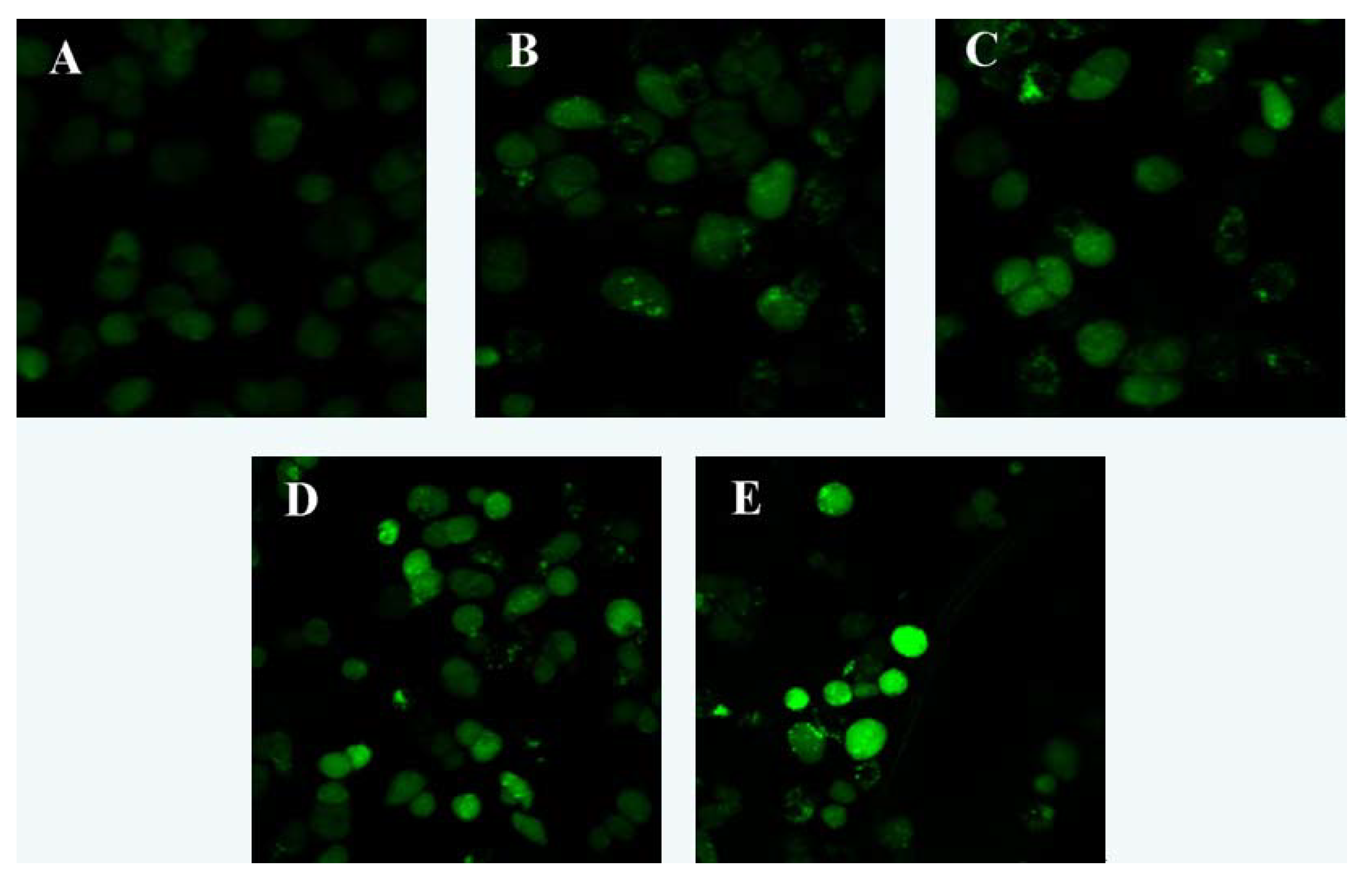
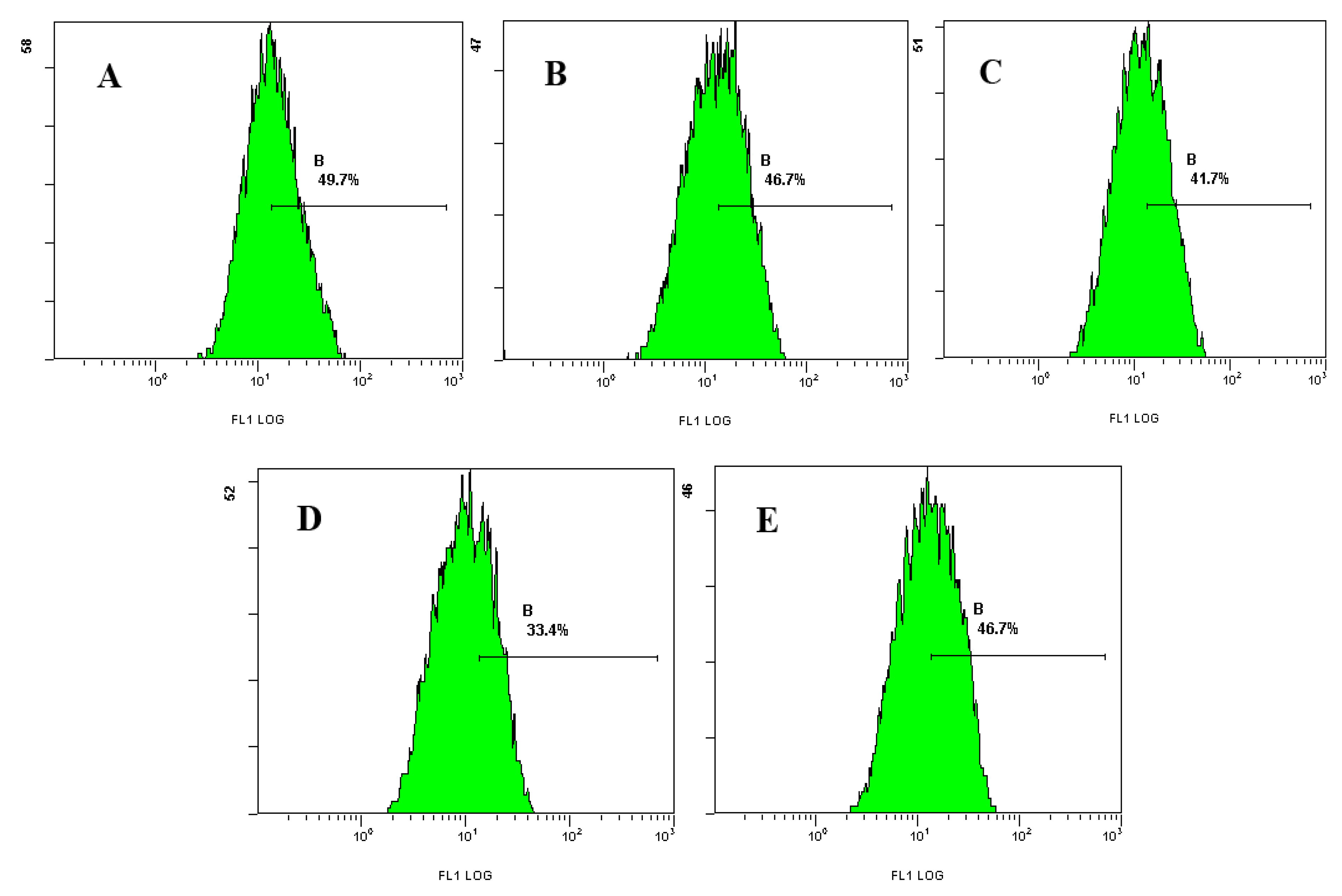
2.3. Effect of Laminarin on Mitochondrion Permeability Transition Pore (MPTP) and Mitochondrial Membrane Potential (MMP) in LOVO Cells
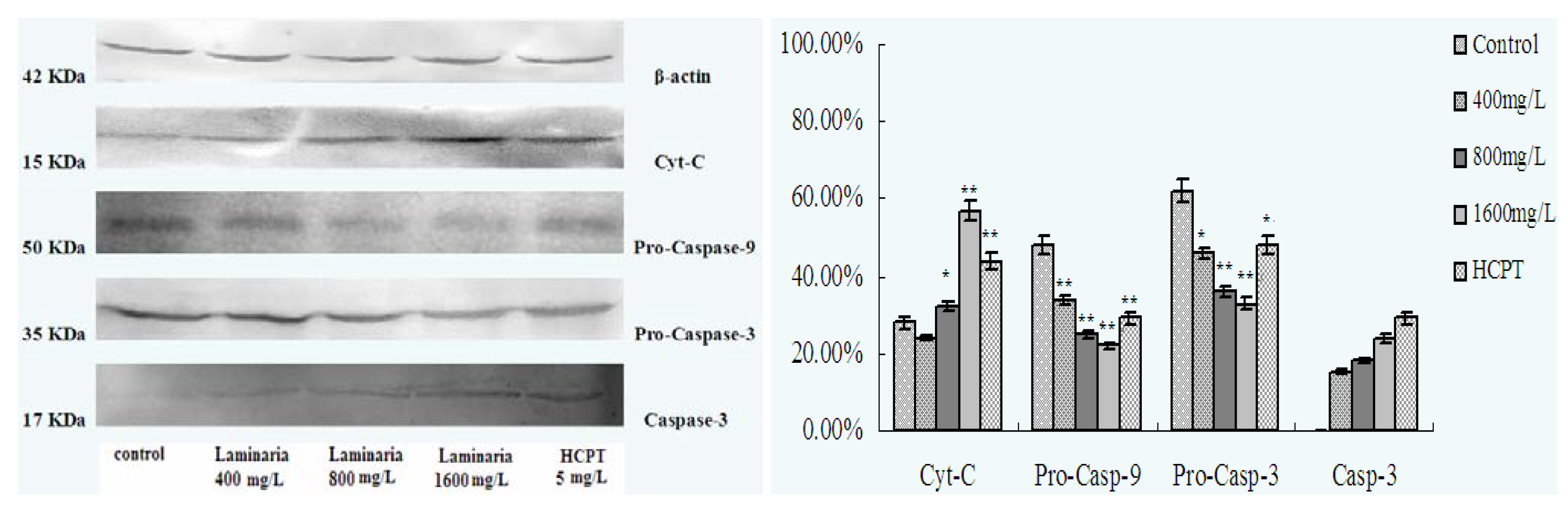
2.4. Effect of Laminarin on the Expression of Cyt-C, Caspase-9 and Caspase-3 in LOVO Cells
3. Experimental
3.1. Reagents
3.2. Cell Culture
3.3. Apoptosis Morphology Observation
3.4. Effect of Laminarin on Reactive Oxygen Species (ROS) in LOVO Cells
3.5. Effect of Laminarin on Calcium Ion Concertration in LOVO Cells
3.6. Effect of Laminarin on pH Value in LOVO Cells
3.7. Effect of Laminarin on Mitochondrion Permeability Transition Pore (MPTP) and Mitochondrial Membrane Potential (MMP) in LOVO Cells
3.8. Effect of Laminarin on the Expression of Cyt-C, Caspase-9 and Caspase-3 in LOVO Cells
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Yuan, X.L.; Lu, J.L.; Xiao, J.; Ma, Q. Research progress of Laminarin. J. Anhui Agric. Sci. 2010, 38, 15447–15448. [Google Scholar]
- Ji, Y.B.; Lou, Y.H.; Gao, S.Y. Separation and purification of Laminarin and study on anti-tumor activity. Chin. Tradil. Herb. Drugs 2009, 40, 132–135. [Google Scholar]
- Fuentes, A.L.; Millis, L.; Sigola, L.B. Laminarin, a soluble beta-glucan, inhibits macrophage phagocytosis of zymosan but has no effect on lipopolysaccharide mediated augmentation of phagocytosis. Int. Immunopharmacol. 2011, 11, 1939–1945. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Kim, Y.J.; Kim, H.J.; Kim, Y.S.; Park, W. Immunostimulatory effect of Laminarin on RAW 264.7 Mouse macrophages. Molecules 2012, 17, 5404–5411. [Google Scholar] [CrossRef] [PubMed]
- Shen, A.; Ji, C.F.; Ji, Y.B. Effect of Laminaria on caspsae-8,3,7 in process of LOVO cells apoptosis. J. Harbin Commer. Univ. (Nat. Sci.) 2010, 26, 641–644. [Google Scholar]
- Indran, I.R.; Tufo, G.; Pervaiz, S.; Brenner, C. Recent advances in apoptosis, mitochondria and drug resistance in cancer cells. Biochim. Biophys. Acta 2011, 1807, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Ju, H.K.; Lee, H.W.; Chung, K.S.; Choi, J.H.; Cho, J.G.; Baek, N.I.; Chung, H.G.; Lee, K.T. Standardized flavonoid-rich fraction of Artemisia princeps Pampanini cv. Sajabal induces apoptosisvia mitochondrial pathway in human cervical cancer HeLa cells. J. Ethnopharmacol. 2012, 141, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Karbowski, M. Mitochondria on guard: Role of mitochondrial fusion and fission in the regulation of apoptosis. Adv. Exp. Med. Biol. 2010, 687, 131–142. [Google Scholar] [PubMed]
- Benarba, B.; Meddah, B.; Aoues, A. Bryonia dioica aqueous extract induces apoptosis through mitochondrial intrinsic pathway in BL41 Burkitt’s lymphoma cells. J. Ethnopharmacol. 2012, 141, 510–516. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, H.; Jin, J.; Zhu, X.; Lu, L.; Jiang, H. The role of endogenous reactive oxygen species in oxymatrine-induced caspase-3-dependent apoptosis in human melanoma A375 cells. Anti-Cancer Drugs 2010, 21, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.M.; Choi, Y.J.; Han, Y.N. N-Dimethyl phytosphingosine induces caspase-8-dependent cytochrome C release and apoptosis through ROS generation in human leukemia cells. Toxicol. Appl. Pharm. 2009, 239, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Li, Y.C.; Ip, S.W.; Hsu, S.C.; Chang, N.W.; Tang, N.Y.; Yu, C.S.; Chou, S.T.; Lin, S.S.; Lino, C.C.; et al. The role of Ca2+ in baicalein-induced apoptosis in human breast MDA-MB-231 cancer cells through mitochondria- and caspase-3-dependent pathway. Anticancer Res. 2008, 28, 1701–1711. [Google Scholar] [PubMed]
- Chattopadhyay, P.; Chaudhury, P.; Wahi, A.K. Ca2+ concentrations are key determinants of ischemia-reperfusion-induced apoptosis: Significance for the molecular mechanism of Bcl-2 action. Appl. Biochem. Biotechnol. 2010, 160, 1968–1977. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.Y.; Liu, X.C.; Ma, W.Y. Study of the relationship between apoptosis and intracellular pH in single living cells using a three-channel real-time fluorescence imaging method (in Chinese). Spectrosc. Spect. Anal. 2009, 29, 1581–1585. [Google Scholar]
- Gao, Y.; Zhao, W.; Shen, W.W.; Fu, A.F.; Zheng, L.; Fu, G.H.; Wu, P.P. Measurement of intracellular pH of gastric cancer AGS cell and effect of acidic extracellular environment on proliferation and apoptosis of AGS cell. Prog. Mod. Biomed. 2008, 8, 1656–1659. [Google Scholar]
- Lee, J.M. The mitochondrial permeability transition pore: A molecular target for amyotrophic lateral sclerosis therapy. BBA-Mol. Basis Dis. 2010, 1802, 186–197. [Google Scholar]
- Leung, A.W.; Halestrap, A.P. Recent progress in elucidating the molecular mechanism of the mitochondrial permeability transition pore. BBA-Bioenergetics 2008, 1777, 946–952. [Google Scholar] [CrossRef] [PubMed]
- Evgeny, P.; Carolina, G.R.; Yuan, Z.; Audrey, C.K.; Ferdinand, J.E.; Michael, A.C. High-frequency photoconductive stimulation reveals central role of mitochondrial permeability transition pore in activity-driven neuronal cell death. Biophys. J. 2010, 98. [Google Scholar] [CrossRef]
- Lemeshko, W. Potential-dependent membrane permeabilization and mitochondrial aggregation caused by anticancer polyarginine-KLA peptides. Arch. Biochem. Biophys. 2010, 493, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Lago, J.; Santaclara, F.; Vieites, J.M.; Cabado, A.G. Collapse of mitochondrial membrane potential and caspases activation are early events in okadaic acid-treated Caco-2 cells. Toxicon 2005, 46, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, J.; Singh, N.; Meena, S.; Sinha, S. Staurosporine induces apoptosis in human papillomavirus positive oral cancer cells at G2/M phase by disrupting mitochondrial membrane potential and modulation of cell cytoskeleton. Oral Oncol. 2009, 45, 974–979. [Google Scholar] [CrossRef] [PubMed]
- Luca, S. Opening the doors to cytochrome C: Changes in mitochondrial shape and apoptosis. Int. J. Biochem. Cell B 2010, 41, 1875–1883. [Google Scholar]
- Dos Santos, A.B.; Dorta, D.J.; Pestana, C.R.; Maioli, M.A.; Curti, C.; Mingatto, F.E. Dehydromonocrotaline induces cyclosporine A-insensitive mitochondrial permeability transition/cytochrome c release. Toxicon 2009, 54, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Kagan, V.E.; Bayir, H.A.; Belikova, N.A.; Kapralov, O.; Tyurina, Y.Y.; Tyurin, V.A.; Jiang, J.; Stoyanovsky, D.A.; Wipf, P.; Kochanek, P.M.; et al. Cytochrome C/cardiolipin relations in mitochondria: A kiss of death. Free Radic. Biol. Med. 2009, 46, 1439–1453. [Google Scholar] [CrossRef] [PubMed]
- Mohan, S.; Abdul, A.B.; Abdelwahab, S.I.; Al-Zubairi, A.S.; Sukari, M.A.; Abdullah, R.; Elhassan Taha, M.M.; Ibrahim, M.Y.; Syam, S. Typhonium flagelliforme induces apoptosis in CEMss cells via activation of caspase-9, PARP cleavage and cytochrome c release: Its activation coupled with G0/G1 phase cell cycle arrest. J. Ethnopharmacol. 2010, 131, 592–600. [Google Scholar] [CrossRef] [PubMed]
- Chien, S.Y.; Wu, Y.C.; Chung, J.G.; Yang, J.S.; Lu, H.F.; Tsou, M.F.; Wood, W.G.; Kuo, S.J.; Chen, D.R. Quercetin-induced apoptosis acts through mitochondrial- and caspase-3-dependent pathways in human breast cancer MDA-MB-231 cells. Hum. Exp. Toxicol. 2009, 28, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Lee, H.J.; Lee, E.O.; Ko, S.G.; Bae, H.S.; Kim, C.H.; Ahn, K.S.; Lu, J.; Kim, S.H. Mitochondria-cytochrome C-caspase-9 cascade mediates isorhamnetin-induced apoptosis. Cancer Lett. 2008, 270, 342–353. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the Laminarin is available from the authors. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Concentration/(µg·mL−1) | MFI/( ± s) |
|---|---|---|
| Control | 0 | 17.1 ± 0.2 |
| I | 400 | 30.6 ± 0.9 ** |
| II | 800 | 36.0 ± 0.2 ** |
| III | 1,600 | 69.4 ± 0.8 ** |
| HCPT | 5 | 37.8 ± 0.6 ** |
| Groups | Concentration/(µg·mL−1) | MFI/( ± s) |
|---|---|---|
| Control | 0 | 24.79 ± 2.49 |
| I | 400 | 35.45 ± 2.82 ** |
| II | 800 | 46.56 ± 1.98 ** |
| III | 1,600 | 51.95 ± 2.39 ** |
| HCPT | 5 | 85.89 ± 2.21 ** |
| Groups | Concentration/(µg·mL−1) | MFI |
|---|---|---|
| Control | 0 | 20.50 ± 1.15 |
| I | 400 | 15.95 ± 0.76 * |
| II | 800 | 13.63 ± 0.74 ** |
| III | 1,600 | 13.08 ± 0.58 ** |
| HCPT | 5 | 18.44 ± 0.45 |
| Groups | Concentration/(µg·mL−1) | MFI/( ± s) | |
|---|---|---|---|
| MPTP | ΔΨm | ||
| Control | 0 | 46.40 ± 2.39 | 64.13 ± 4.22 |
| I | 400 | 38.64 ± 1.87 * | 44.80 ± 1.97 ** |
| II | 800 | 31.30 ± 1.41 ** | 28.37 ± 7.85 ** |
| III | 1,600 | 3.62 ± 2.03 ** | 20.17 ± 4.06 ** |
| HCPT | 5 | 6.89 ± 1.38 ** | 27.55 ± 9.16 ** |
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ji, Y.B.; Ji, C.F.; Zhang, H. Laminarin Induces Apoptosis of Human Colon Cancer LOVO Cells through a Mitochondrial Pathway. Molecules 2012, 17, 9947-9960. https://doi.org/10.3390/molecules17089947
Ji YB, Ji CF, Zhang H. Laminarin Induces Apoptosis of Human Colon Cancer LOVO Cells through a Mitochondrial Pathway. Molecules. 2012; 17(8):9947-9960. https://doi.org/10.3390/molecules17089947
Chicago/Turabian StyleJi, Yu Bin, Chen Feng Ji, and He Zhang. 2012. "Laminarin Induces Apoptosis of Human Colon Cancer LOVO Cells through a Mitochondrial Pathway" Molecules 17, no. 8: 9947-9960. https://doi.org/10.3390/molecules17089947