Formononetin Attenuates IL-1β-Induced Apoptosis and NF-κB Activation in INS-1 Cells

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
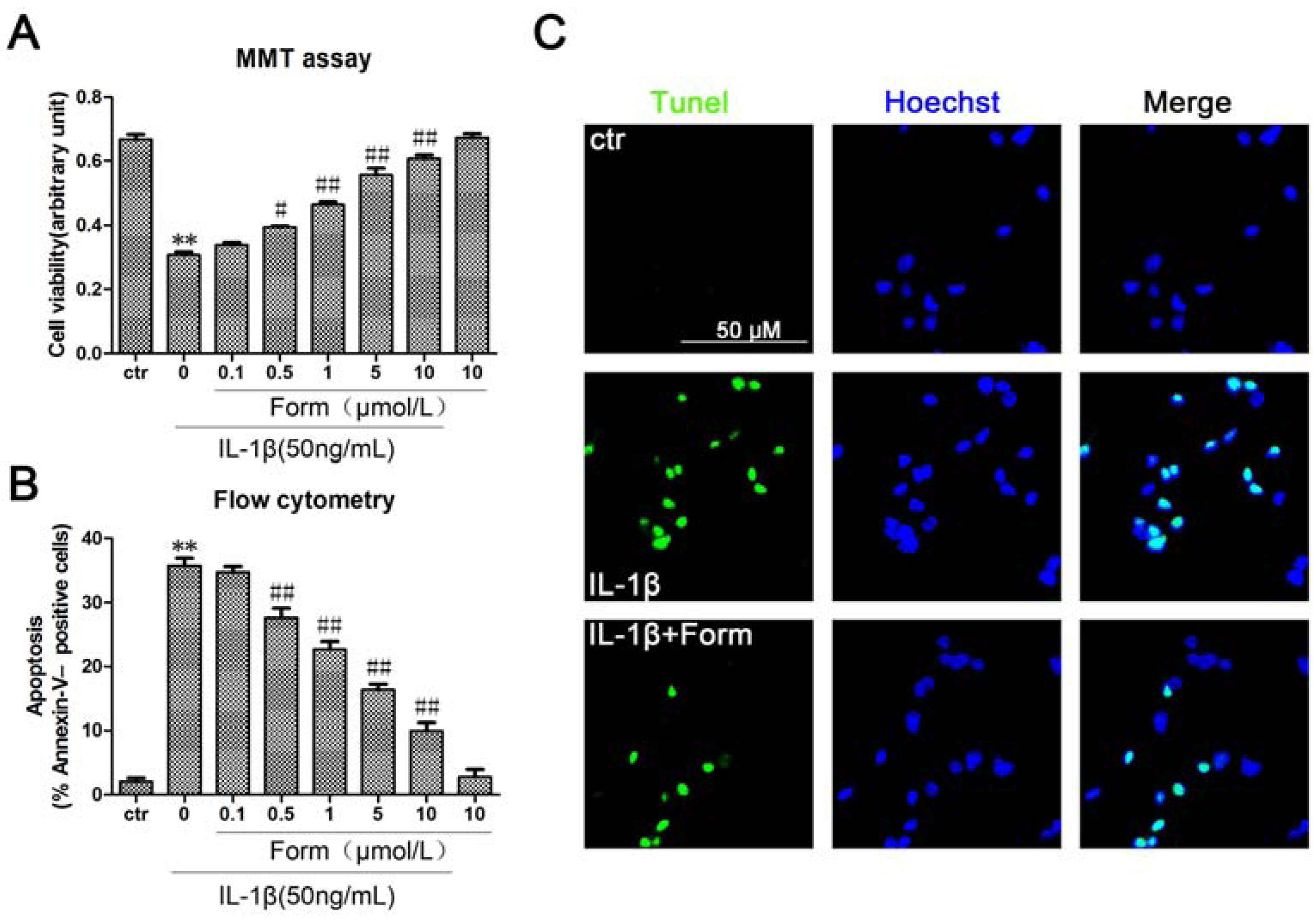
2.1. Formononetin (Form) Protects INS-1 Cells from IL-1β-Induced Apoptosis

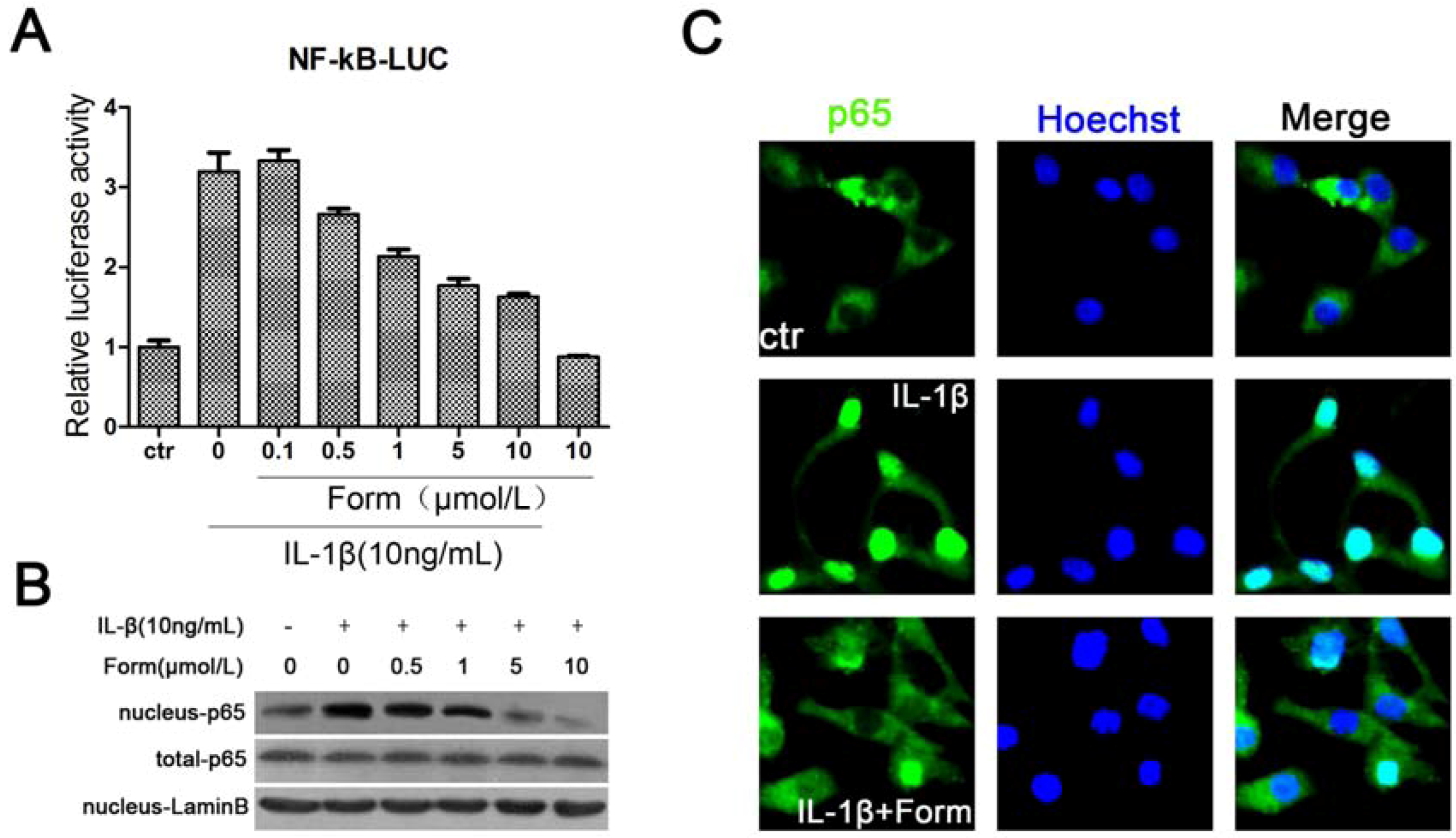
2.2. Formononetin (Form) Suppressed IL-1β-Induced Activation of NF-κB in INS-1 Cells

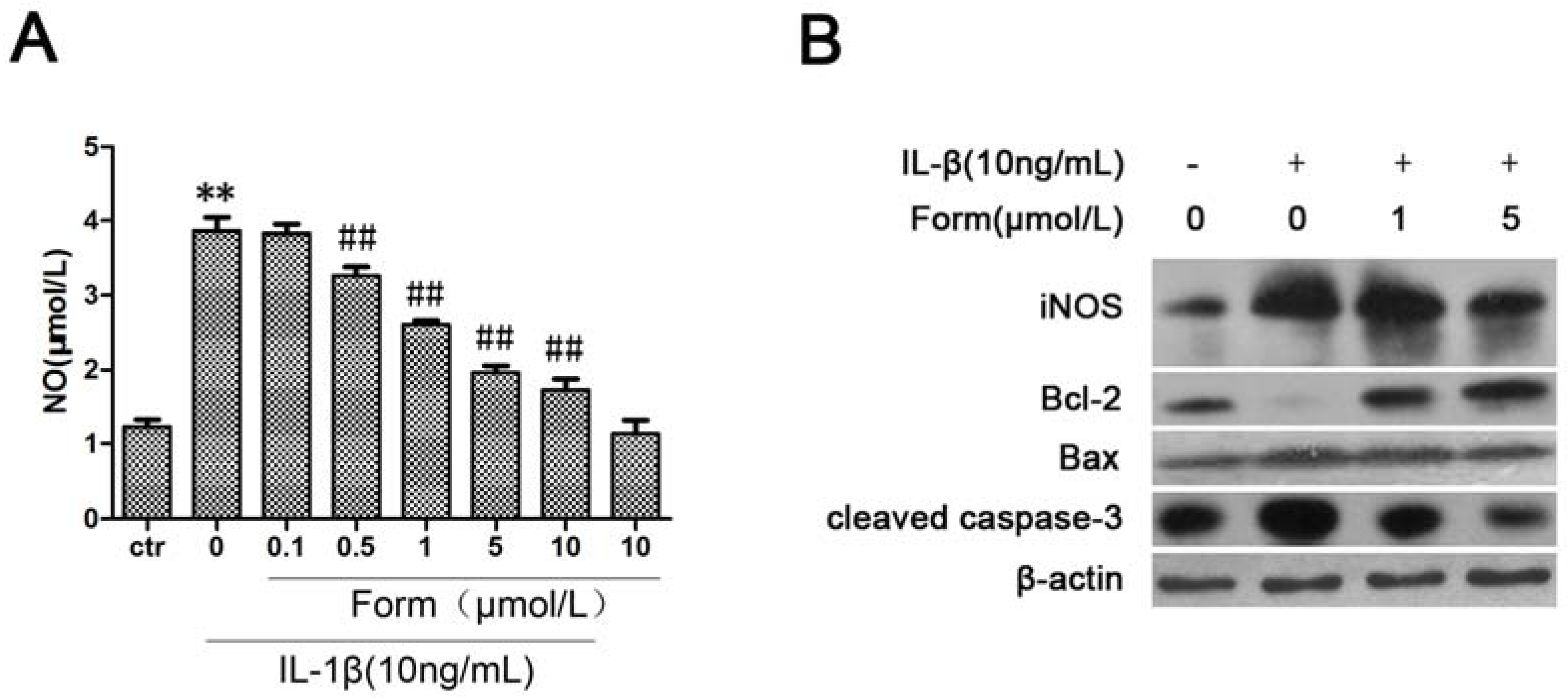
2.3. Formononetin (Form) Inhibited IL-1β-Induced NO Formation and Apoptotic Pathway in INS-1 Cells

3. Discussion
4. Experimental
4.1. Reagents
4.2. Cell Culture
4.3. MTT Assay
4.4. Flow Cytometry
4.5. Tunel Staining
4.6. Immunofluorescence Microscopy
4.7. Transient Transfection and Luciferase Reporter Assay
4.8. Western Blot
4.9. NO Assay
4.10. Statistical Analysis
5. Conclusions
Acknowledgements
Conflict of Interests
References
- Wang, C.; Guan, Y.; Yang, J. Cytokines in the Progression of Pancreatic beta-Cell Dysfunction. Int. J. Endocrinol. 2010, 2010, 515136:1–515136:10. [Google Scholar]
- Rabinovitch, A.; Suarez-Pinzon, W.L.; Strynadka, K.; Lakey, J.R.; Rajotte, R.V. Human pancreatic islet beta-cell destruction by cytokines involves oxygen free radicals and aldehyde production. J. Clin. Endocrinol. Metab. 1996, 81, 3197–3202. [Google Scholar] [CrossRef]
- Collier, J.J.; Burke, S.J.; Eisenhauer, M.E.; Lu, D.; Sapp, R.C.; Frydman, C.J.; Campagna, S.R. Pancreatic beta-cell death in response to pro-inflammatory cytokines is distinct from genuine apoptosis. PLoS One 2011, 6, e22485. [Google Scholar]
- Bowie, A.; O’Neill, L.A. The interleukin-1 receptor/Toll-like receptor superfamily: Signal generators for pro-inflammatory interleukins and microbial products. J. Leuk. Biol. 2000, 67, 508–514. [Google Scholar]
- Ortis, F.; Miani, M.; Colli, M.L.; Cunha, D.A.; Gurzov, E.N.; Allagnat, F.; Chariot, A.; Eizirik, D.L. Differential usage of NF-kappaB activating signals by IL-1beta and TNF-alpha in pancreatic beta cells. FEBS Lett. 2012, 586, 984–989. [Google Scholar] [CrossRef]
- Eda, H.; Shimada, H.; Beidler, D.R.; Monahan, J.B. Proinflammatory cytokines, IL-1beta and TNF-alpha, Induce expression of interleukin-34 mRNA via JNK- and p44/42 MAPK-NF-kappaB pathway but not p38 pathway in osteoblasts. Rheumatol. Int. 2011, 31, 1525–1530. [Google Scholar] [CrossRef]
- Choi, E.M.; Lee, Y.S. Luteolin suppresses IL-1beta-induced cytokines and MMPs production via p38 MAPK, JNK, NF-kappaB and AP-1 activation in human synovial sarcoma cell line, SW982. Food Chem. Toxicol. 2010, 48, 2607–2611. [Google Scholar] [CrossRef]
- Sandberg, J.O.; Eizirik, D.L.; Sandler, S. IL-1 receptor antagonist inhibits recurrence of disease after syngeneic pancreatic islet transplantation to spontaneously diabetic non-obese diabetic (NOD) mice. Clin. Exp. Immunol. 1997, 108, 314–317. [Google Scholar]
- Sauter, N.S.; Schulthess, F.T.; Galasso, R.; Castellani, L.W.; Maedler, K. The antiinflammatory cytokine interleukin-1 receptor antagonist protects from high-fat diet-induced hyperglycemia. Endocrinology 2008, 149, 2208–2218. [Google Scholar]
- Mandrup-Poulsen, T.; Pickersgill, L.; Donath, M.Y. Blockade of interleukin 1 in type 1 diabetes mellitus. Nat. Rev. Endocrinol. 2010, 6, 158–166. [Google Scholar] [CrossRef]
- Lin, J.; Yan, G.T. Function of eukaryotic transcription factor NF-kappaB in the signal transduction of acute inflammatory response. Zhongguo Wei Zhong Bing Ji Jiu Yi Xue 2003, 15, 45–48. [Google Scholar]
- Gu, Q.; Kong, Y.; Yu, Z.B.; Bai, L.; Xiao, Y.B. Hypoxia-induced SOCS3 is limiting STAT3 phosphorylation and NF-kappaB activation in congenital heart disease. Biochimie 2011, 93, 909–920. [Google Scholar] [CrossRef]
- Wang, Q.S.; Xiang, Y.; Cui, Y.L.; Lin, K.M.; Zhang, X.F. Dietary blue pigments derived from genipin, Attenuate inflammation by inhibiting LPS-Induced iNOS and COX-2 expression via the NF-kappaB inactivation. PLoS One 2012, 7, e34122. [Google Scholar]
- Mattson, M.P.; Camandola, S. NF-kappaB in neuronal plasticity and neurodegenerative disorders. J. Clin. Invest. 2001, 107, 247–254. [Google Scholar] [CrossRef]
- Stark, L.A.; Dunlop, M.G. Nucleolar sequestration of RelA (p65) regulates NF-kappaB-driven transcription and apoptosis. Mol. Cell. Biol. 2005, 25, 5985–6004. [Google Scholar] [CrossRef]
- Autieri, M.V.; Yue, T.L.; Ferstein, G.Z.; Ohlstein, E. Antisense oligonucleotides to the p65 subunit of NF-kB inhibit human vascular smooth muscle cell adherence and proliferation and prevent neointima formation in rat carotid arteries. Biochem. Biophys. Res. Commun. 1995, 213, 827–836. [Google Scholar] [CrossRef]
- Heimberg, H.; Heremans, Y.; Jobin, C.; Leemans, R.; Cardozo, A.K.; Darville, M.; Eizirik, D.L. Inhibition of cytokine-induced NF-kappaB activation by adenovirus-mediated expression of a NF-kappaB super-repressor prevents beta-cell apoptosis. Diabetes 2001, 50, 2219–2224. [Google Scholar]
- Eizirik, D.L.; Darville, M.I. Beta-cell apoptosis and defense mechanisms: Lessons from type 1 diabetes. Diabetes 2001, 50 Suppl. 1, S64–S69. [Google Scholar] [CrossRef]
- Spinas, G.A. The dual role of nitric oxide in islet beta-cells. News Physiol. Sci. 1999, 14, 49–54. [Google Scholar]
- Welsh, N.; Eizirik, D.L.; Bendtzen, K.; Sandler, S. Interleukin-1 beta-induced nitric oxide production in isolated rat pancreatic islets requires gene transcription and may lead to inhibition of the Krebs cycle enzyme aconitase. Endocrinology 1991, 129, 3167–3173. [Google Scholar] [CrossRef]
- Storling, J.; Binzer, J.; Andersson, A.K.; Zullig, R.A.; Tonnesen, M.; Lehmann, R.; Spinas, G.A.; Sandler, S.; Billestrup, N.; Mandrup-Poulsen, T. Nitric oxide contributes to cytokine-induced apoptosis in pancreatic beta cells via potentiation of JNK activity and inhibition of Akt. Diabetologia 2005, 48, 2039–2050. [Google Scholar] [CrossRef]
- Marzocchella, L.; Fantini, M.; Benvenuto, M.; Masuelli, L.; Tresoldi, I.; Modesti, A.; Bei, R. Dietary flavonoids: Molecular mechanisms of action as anti-inflammatory agents. Recent Pat. Inflamm. Allergy Drug Discov. 2011, 5, 200–220. [Google Scholar]
- Chen, C.Y.; Peng, W.H.; Tsai, K.D.; Hsu, S.L. Luteolin suppresses inflammation-associated gene expression by blocking NF-kappaB and AP-1 activation pathway in mouse alveolar macrophages. Life Sci. 2007, 81, 1602–1614. [Google Scholar] [CrossRef]
- Hamalainen, M.; Nieminen, R.; Vuorela, P.; Heinonen, M.; Moilanen, E. Anti-inflammatory effects of flavonoids: Genistein, Kaempferol, Quercetin, and Daidzein inhibit STAT-1 and NF-kappaB activations, Whereas flavone, Isorhamnetin, Naringenin, and Pelargonidin inhibit only NF-kappaB activation along with their inhibitory effect on iNOS expression and NO production in activated macrophages. Mediators Inflamm. 2007, 45673. [Google Scholar]
- Shams, M.E.; Al-Gayyar, M.M.; Barakat, E.A. Type 2 Diabetes mellitus-induced hyperglycemia in patients with nafld and normal LFTs: Relationship to lipid profile, Oxidative stress and pro-Inflammatory cytokines. Sci. Pharm. 2011, 79, 623–634. [Google Scholar]
- Papaccio, G.; Graziano, A.; D’Aquino, R.; Valiante, S.; Naro, F. A biphasic role of nuclear transcription factor (NF)-kappaB in the islet beta-cell apoptosis induced by interleukin (IL)-1beta. J. Cell. Physiol. 2005, 204, 124–130. [Google Scholar] [CrossRef]
- Castrillo, A.; Bodelon, O.G.; Bosca, L. Inhibitory effect of IGF-I on type 2 nitric oxide synthase expression in Ins-1 cells and protection against activation-dependent apoptosis: Involvement of phosphatidylinositol 3-kinase. Diabetes 2000, 49, 209–217. [Google Scholar]
- Noguchi, A.; Takada, M.; Nakayama, K.; Ishikawa, T. cGMP-independent anti-apoptotic effect of nitric oxide on thapsigargin-induced apoptosis in the pancreatic beta-cell line INS-1. Life Sci. 2008, 83, 865–870. [Google Scholar] [CrossRef]
- Dinarello, C.A.; Donath, M.Y.; Mandrup-Poulsen, T. Role of IL-1beta in type 2 diabetes. Curr. Opin. Endocrinol. Diabetes Obes. 2010, 17, 314–321. [Google Scholar]
- Doganay, S.; Evereklioglu, C.; Er, H.; Turkoz, Y.; Sevinc, A.; Mehmet, N.; Savli, H. Comparison of serum NO, TNF-alpha, IL-1beta, sIL-2R, IL-6 and IL-8 levels with grades of retinopathy in patients with diabetes mellitus. Eye (Lond) 2002, 16, 163–170. [Google Scholar] [CrossRef]
- Cavelti-Weder, C.; Furrer, R.; Keller, C.; Babians-Brunner, A.; Solinger, A.M.; Gast, H.; Fontana, A.; Donath, M.Y.; Penner, I.K. Inhibition of IL-1beta improves fatigue in type 2 diabetes. Diabetes Care 2011, 34, e158. [Google Scholar] [CrossRef]
- O’Connor, J.C.; Satpathy, A.; Hartman, M.E.; Horvath, E.M.; Kelley, K.W.; Dantzer, R.; Johnson, R.W.; Freund, G.G. IL-1beta-mediated innate immunity is amplified in the db/db mouse model of type 2 diabetes. J. Immunol. 2005, 174, 4991–4997. [Google Scholar]
- Yin, H.; Miao, J.; Ma, C.; Sun, G.; Zhang, Y. Beta-Casomorphin-7 cause decreasing in oxidative stress and inhibiting NF-kappaB-iNOS-NO signal pathway in pancreas of diabetes rats. J. Food Sci. 2012, 77, C278–C282. [Google Scholar]
- Oriot, P.; Hermans, M.P.; Selvais, P.; Buysschaert, M.; de la Tribonniere, X. Exenatide improves weight loss insulin sensitivity and beta-cell function following administration to a type 2 diabetic HIV patient on antiretroviral therapy. Ann. Endocrinol. 2011, 72, 244–246. [Google Scholar] [CrossRef]
- Liadis, N.; Murakami, K.; Eweida, M.; Elford, A.R.; Sheu, L.; Gaisano, H.Y.; Hakem, R.; Ohashi, P.S.; Woo, M. Caspase-3-dependent beta-cell apoptosis in the initiation of autoimmune diabetes mellitus. Mol. Cell. Biol. 2005, 25, 3620–3629. [Google Scholar] [CrossRef]
- Gurzov, E.N.; Eizirik, D.L. Bcl-2 proteins in diabetes: Mitochondrial pathways of beta-cell death and dysfunction. Trends Cell Biol. 2011, 21, 424–431. [Google Scholar] [CrossRef]
- Li, Y.H.; Zhuo, Y.H.; Lu, L.; Chen, L.Y.; Huang, X.H.; Zhang, J.L.; Li, S.Y.; Wang, X.G. Caspase-dependent retinal ganglion cell apoptosis in the rat model of acute diabetes. Chin. Med. J. (Engl.) 2008, 121, 2566–2571. [Google Scholar]
- Augstein, P.; Bahr, J.; Wachlin, G.; Heinke, P.; Berg, S.; Salzsieder, E.; Harrison, L.C. Cytokines activate caspase-3 in insulinoma cells of diabetes-prone NOD mice directly and via upregulation of Fas. J. Autoimmun. 2004, 23, 301–309. [Google Scholar] [CrossRef]
- Ghosh, S.; Khazaei, M.; Moien-Afshari, F.; Ang, L.S.; Granville, D.J.; Verchere, C.B.; Dunn, S.R.; McCue, P.; Mizisin, A.; Sharma, K.; et al. Moderate exercise attenuates caspase-3 activity, Oxidative stress, And inhibits progression of diabetic renal disease in db/db mice. Am. J. Physiol. Renal Physiol. 2009, 296, F700–F708. [Google Scholar]
- Sun, M.; Zhou, T.; Zhou, L.; Chen, Q.; Yu, Y.; Yang, H.; Zhong, K.; Zhang, X.; Xu, F.; Cai, S.; et al. Formononetin protects neurons against hypoxia-induced cytotoxicity through upregulation of ADAM10 and sAbetaPPalpha. J. Alzheimers Dis. 2012, 28, 795–808. [Google Scholar]
- Wang, Q.Y.; Meng, Q.H.; Zhang, Z.T.; Tian, Z.J.; Liu, H. Synthesis, solubility, Lipids-lowering and liver-protection activities of sulfonated formononetin. Yao Xue Xue Bao 2009, 44, 386–389. [Google Scholar]
- Harini, R.; Ezhumalai, M.; Pugalendi, K.V. Antihyperglycemic effect of biochanin A, A soy isoflavone, on streptozotocin-diabetic rats. Eur. J. Pharmacol. 2012, 676, 89–94. [Google Scholar] [CrossRef]
- Kole, L.; Giri, B.; Manna, S.K.; Pal, B.; Ghosh, S. Biochanin-A, An isoflavon, Showed anti-proliferative and anti-inflammatory activities through the inhibition of iNOS expression, p38-MAPK and ATF-2 phosphorylation and blocking NFkappaB nuclear translocation. Eur. J. Pharmacol. 2011, 653, 8–15. [Google Scholar] [CrossRef]
- Han, X.; Sun, Y.; Scott, S.; Bleich, D. Tissue inhibitor of metalloproteinase-1 prevents cytokine-mediated dysfunction and cytotoxicity in pancreatic islets and beta-cells. Diabetes 2001, 50, 1047–1055. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, L.; Li, Y.; Chen, H.; Sun, Z. Nifedipine protects INS-1 beta-cell from high Glucose-induced ER stress and apoptosis. Int. J. Mol. Sci. 2011, 12, 7569–7580. [Google Scholar]
- Sample Availability: All samples used in this study are available from the author.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, Y.; Zhu, Y.; Gao, L.; Yin, H.; Xie, Z.; Wang, D.; Zhu, Z.; Han, X. Formononetin Attenuates IL-1β-Induced Apoptosis and NF-κB Activation in INS-1 Cells. Molecules 2012, 17, 10052-10064. https://doi.org/10.3390/molecules170910052
Wang Y, Zhu Y, Gao L, Yin H, Xie Z, Wang D, Zhu Z, Han X. Formononetin Attenuates IL-1β-Induced Apoptosis and NF-κB Activation in INS-1 Cells. Molecules. 2012; 17(9):10052-10064. https://doi.org/10.3390/molecules170910052
Chicago/Turabian StyleWang, Yao, Yunxia Zhu, Lu Gao, Han Yin, Zuoling Xie, Dong Wang, Zhengqiu Zhu, and Xiao Han. 2012. "Formononetin Attenuates IL-1β-Induced Apoptosis and NF-κB Activation in INS-1 Cells" Molecules 17, no. 9: 10052-10064. https://doi.org/10.3390/molecules170910052