The Effect of Camphor and Borneol on Rat Thymocyte Viability and Oxidative Stress

Abstract
:1. Introduction
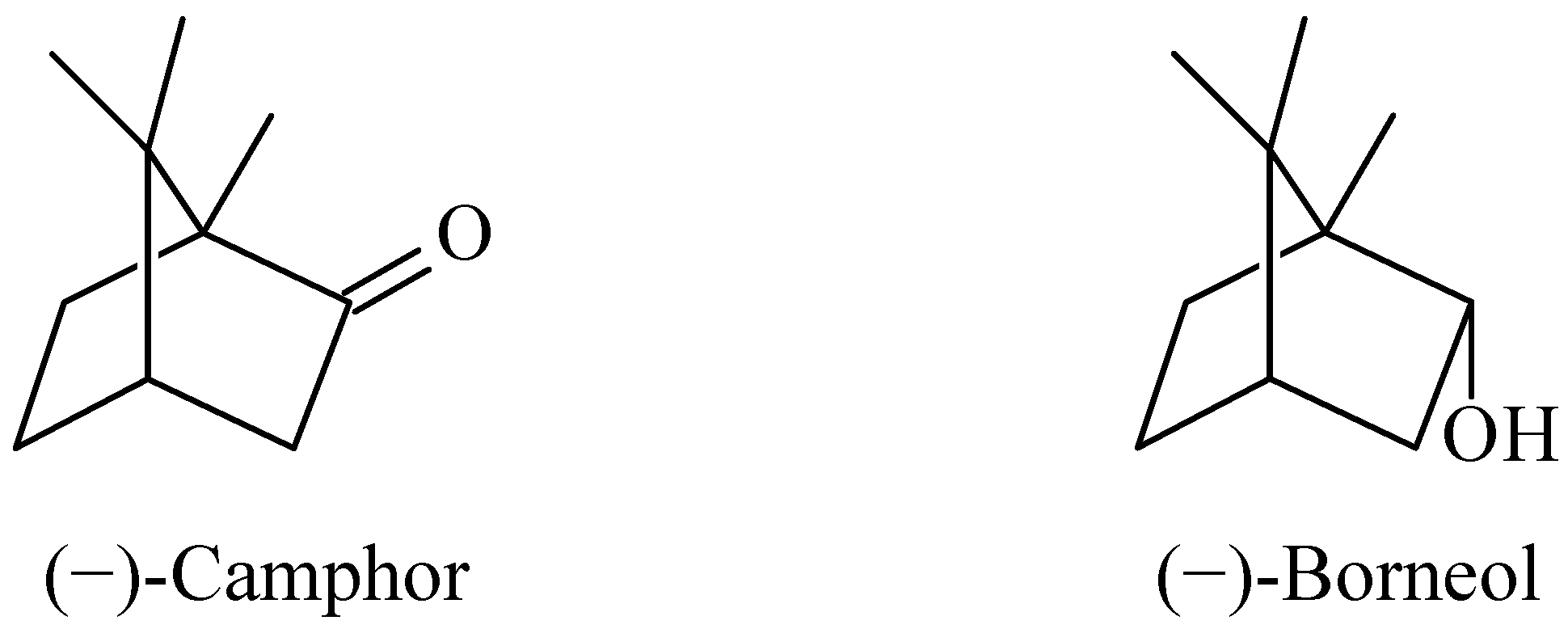
2. Results and Discussion

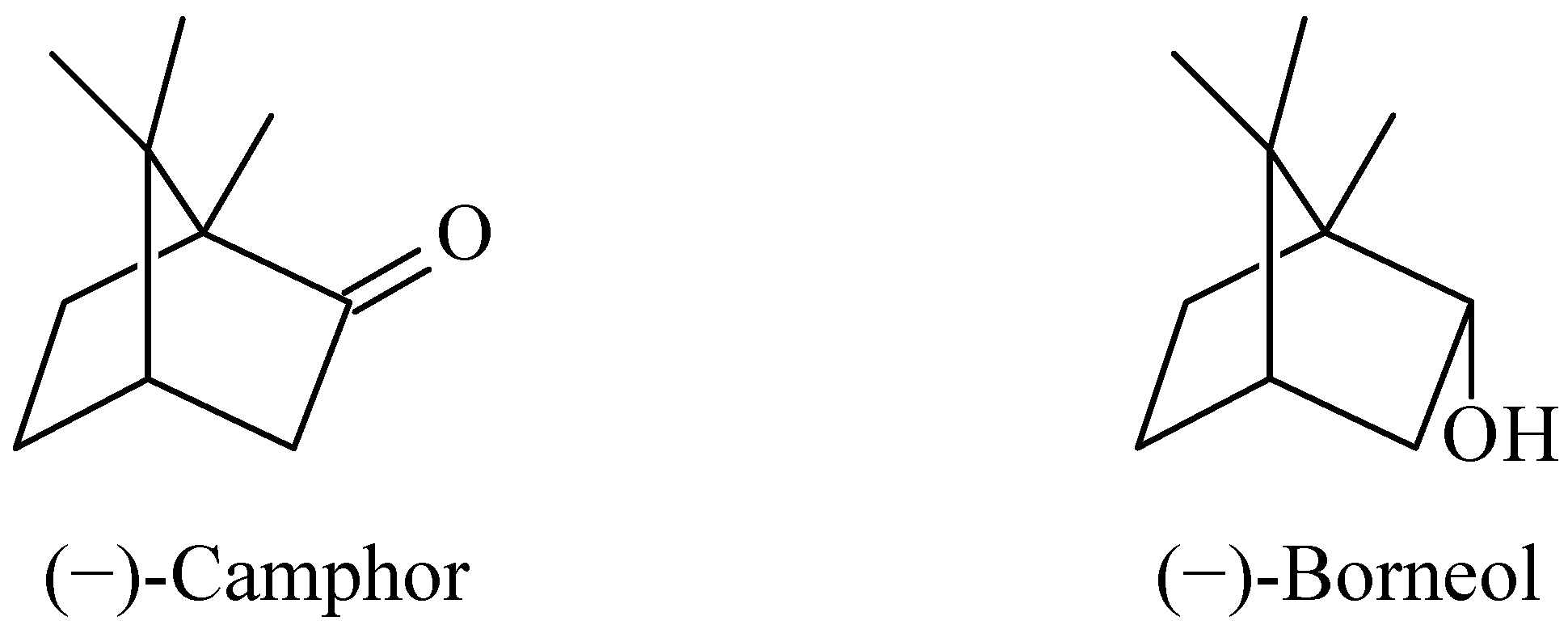{kind=link}
| Compound | Concentration (µg/mL) | Cytotoxicity (CCK-8 assay) |
|---|---|---|
| 0.5 | 0.927 ± 0.132 | |
| Camphor | 5 | 1.024 ± 0.083 |
| 50 | 1.111 ± 0.033 * | |
| 0.5 | 1.067 ± 0.004 ** | |
| Borneol | 5 | 1.070 ± 0.015 ** |
| 50 | 0.975 ± 0.144 | |
| Control cells | 0.999 ± 0.005 |
| Compound | Concentration (µg/mL) | Intracellular ROS production |
|---|---|---|
| 0.5 | 1.413 ± 0.068 *** | |
| Camphor | 5 | 1.399 ± 0.093 ** |
| 50 | 1.251 ± 0.049 * | |
| 0.5 | 1.268 ± 0.102 | |
| Borneol | 5 | 1.224 ± 0.108 |
| 50 | 1.164 ± 0.191 | |
| Control cells | 1 ± 0.042 |
| Compound | Concentration (µg/mL) | Mitochondrial membrane potential |
|---|---|---|
| 0.5 | 0.740 ± 0.071 ** | |
| Camphor | 5 | 0.919 ± 0.047 |
| 50 | 0.925 ± 0.132 | |
| 0.5 | 0.799 ± 0.022 *** | |
| Borneol | 5 | 0.856 ± 0.084 |
| 50 | 0.876 ± 0.110 | |
| Control cells | 0.999 ± 0.076 |
3. Experimental
3.1. Animals
3.2. Materials
3.3. Preparation of Thymocytes
3.4. Cell Culture
3.5. Analysis of Cell Viability
3.6. Measurement of Intracellular Reactive Oxygen Species (ROS) Production
3.7. Determination of Mitochondrial Membrane Potential
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Croteau, R. Biosynthesis and catabolism of monoterpenoids. Chem. Rev. 1987, 87, 929–954. [Google Scholar]
- Prasad, B.; Rojubally, A.; Plettner, E. Identification of camphor oxidation and reduction products in Pseudomonas putida: New activity of the cytochrome P450cam system. J. Chem. Ecol. 2011, 37, 657–667. [Google Scholar] [CrossRef]
- Tucker, A.O.; Maciarello, M.J. Essential oils of cultivars of Dalmatian sage (Salvia officinalis L.). J. Essent. Oil Res. 1990, 2, 139–144. [Google Scholar]
- Love, J.N.; Sammon, M.; Smereck, J. Are one or two dangerous? Camphor exposure in toddlers. J. Emerg. Med. 2004, 27, 49–54. [Google Scholar] [CrossRef]
- Nakahashi, H.; Miyazawa, M. Biotransformation of (−)-camphor by Salmonella typhimurium OY1002/2A6 expressing human CYP2A6 and NADPH-P450 reductase. J. Oleo Sci. 2011, 60, 545–548. [Google Scholar] [CrossRef]
- Dai, J.-P.; Chen, J.; Bei, Y.-F.; Han, B.-X.; Wang, S. Influence of borneol on primary mice oral fibroblasts: A penetration enhancer may be used in oral submucous fibrosis. J. Oral Pathol. Med. 2009, 38, 276–281. [Google Scholar] [CrossRef]
- Gershenzon, J.; Dudareva, N. The function of terpene natural products in the natural world. Nat. Chem. Biol. 2007, 3, 408–414. [Google Scholar] [CrossRef]
- Langenheim, J.H. Higher plant terpenoids: A phytocentric overview of their ecological roles. J. Chem. Ecol. 1994, 20, 1223–1280. [Google Scholar] [CrossRef]
- Bhatia, S.P.; Letizia, C.S.; Api, A.M. Fragrance material review on borneol. Food Chem. Toxicol. 2008, 46, S77–S80. [Google Scholar]
- The RIFM EXPERT Panel; Belsito, D.; Bickers, D.; Bruze, M.; Calow, P.; Greim, H.; Hanifin, J.M.; Rogers, A.E.; Saurat, J.H.; Sipes, I.G.; Tagami, H. A toxicologic and dermatologic assessment of cyclic and non-cyclic terpene alcohols when used as fragrance ingredients. Food Chem. Toxicol. 2008, 46, S1–S71. [Google Scholar]
- Gherlardini, C.; Galeotti, N.; Mazzanti, G. Local anaesthetic activity of monoterpenes and phenylpropanes of essential oils. Planta Med. 2001, 67, 564–566. [Google Scholar] [CrossRef]
- Santos, F.A.; Rao, V.S.N. Antiinflammatory and antinociceptive effects of 1,8-cineole a terpenoid oxide present in many plant essential oils. Phytother. Res. 2000, 14, 240–244. [Google Scholar] [CrossRef]
- Hattori, A. Camphor in the Edo era—Camphor and borneol for medicines. Yakushigaku Zasshi 2000, 35, 49–54. [Google Scholar]
- Granger, R.E.; Campbell, E.L.; Johnston, G.A. (+)- And (−)-borneol: Efficacious positive modulators of GABA action at human recombinant alpha1beta2gamma2L GABA(A) receptors. Biochem. Pharmacol. 2005, 69, 1101–1111. [Google Scholar]
- Shunying, Z.; Yang, Y.; Huaidong, Y.; Yue, Y.; Guolin, Z. Chemical composition and antimicrobial activity of the essential oils of Chrysanthemum indicum. J. Ethnopharmacol. 2005, 96, 151–158. [Google Scholar] [CrossRef]
- Park, T.-J.; Park, Y.-S.; Lee, T.-G.; Ha, H.; Kim, K.-T. Inhibition of acetylcholine-mediated effects by borneol. Biochem. Pharmacol. 2003, 65, 83–90. [Google Scholar]
- Mühlbauer, R.C.; Lozano, A.; Palacio, S.; Reinli, A.; Felix, R. Common herbs, essential oils, and monoterpenes potently modulate bone metabolism. Bone 2003, 32, 372–380. [Google Scholar] [CrossRef]
- Mohsenzadeh, F.; Chehregani, A.; Amiri, H. Chemical composition, antibacterial activity and cytotoxicity of essential oils of Tanacetum parthenium in different developmental stages. Pharm. Biol. 2011, 49, 920–926. [Google Scholar] [CrossRef]
- Horváthová, E.; Slamenová, D.; Marsálková, L.; Sramková, M.; Wsólová, L. Effects of borneol on the level of DNA damage induced in primary rat hepatocytes and testicular cells by hydrogen peroxide. Food Chem. Toxicol. 2009, 47, 1318–1323. [Google Scholar] [CrossRef]
- Slamenová, D.; Horváthová, E.; Wsólová, L.; Sramková, M.; Navarová, J. Investigation of anti-oxidative, cytotoxic, DNA-damaging and DNA-protective effects of plant volatiles eugenol and borneol in human-derived HepG2, Caco-2 and VH10 cell lines. Mutat. Res. 2009, 677, 46–52. [Google Scholar]
- Horváthová, E.; Kozics, K.; Srancíková, A.; Hunáková, L.; Gálová, E.; Sevcovicová, A.; Slamenová, D. Borneol administration protects primary rat hepatocytes against exogenous oxidative DNA damage. Mutagenesis 2012. [Google Scholar] [CrossRef]
- Victor, V.M.; Guayerbas, N.; de la Fuente, M. Changes in the antioxidant content of mononuclear leukocytes from mice with endotoxin-induced oxidative stress. Mol. Cell Biochem. 2002, 29, 107–111. [Google Scholar]
- Hildeman, D.A.; Mitchell, T.; Aronow, B.; Wojciechowski, S.; Kappler, J.; Marrack, P. Control of Bcl-2 expression by reactive oxygen species. Proc. Natl. Acad. Sci. USA 2003, 100, 15035–15040. [Google Scholar]
- Pavlovic, V.; Sarac, M. A short overview of vitamin C and selected cells of the immune system. Cent. Eur. J. Med. 2011, 6, 1–10. [Google Scholar] [CrossRef]
- Ciobica, A.; Bild, V.; Hritcu, L.; Padurariu, M.; Bild, W. Effects of angiotensin II receptor antagonists on anxiety and some oxidative stress markers in rat. Cent. Eur. J. Med. 2011, 6, 331–340. [Google Scholar] [CrossRef]
- Exline, M.C.; Crouser, E.D. Mitochondrial mechanisms of sepsis induced organ failure. Front. Biosci. 2008, 13, 5030–5041. [Google Scholar]
- Leist, M.; Single, B.; Castoldi, A.F.; Kuhnle, S.; Nicotera, P. Intracellular adenosine triphosphate (ATP) concentration: A switch in the decision between apoptosis and necrosis. J. Exp. Med. 1997, 185, 1481–1486. [Google Scholar] [CrossRef]
- Pavlovic, V.; Cekic, S.; Kocic, G.; Sokolovic, D.; Zivkovic, V. Effect of monosodium glutamate on apoptosis and Bcl-2/Bax protein level in rat thymocyte culture. Physiol. Res. 2007, 56, 619–626. [Google Scholar]
- Cekic, S.; Pavlovic, D.; Sarac, M.; Kamenov, B.; Dimic, A.; Pavlovic, V. The effect of vitamina C on amiodarone-induced toxicity in rat thymocytes. Cent. Eur. J. Med. 2011, 6, 58–63. [Google Scholar] [CrossRef]
- Hori, R.; Kashiba, M.; Toma, T.; Yachie, A.; Goda, N.; Makino, N; Soejima, A.; Nagasawa, T.; Nakabayashi, K.; Suematsu, M. Gene transfection of H25A mutant heme oxygenase-1 protects cells against hydroperoxide-induced cytotoxicity. J. Biol. Chem. 2002, 277, 10712–10718. [Google Scholar]
- Das, A.; Hazra, T.K.; Boldogh, I.; Mitra, S.; Bhakat, K.K. Induction of the human oxidized base-specific DNA glycosylase Neil1 by reactive oxygen species. J. Biol. Chem. 2005, 280, 35272–35280. [Google Scholar]
- Boldogh, I.; Roy, G.; Lee, M.S.; Bacsi, A; Hazra, T.K.; Bhakat, K.K.; Das, G.C., Mitra. Reduced DNA double strand breaks in chlorambucil resistant cells are related to high DNAPKCs activity and low oxidative stress. Toxicology 2003, 193, 137–152. [Google Scholar]
- Wang, Z.B.; Liu, Y.Q.; Zhang, Y.; Li, Y.; An, X.X.; Xu, H.; Guo, Y.; Jin, W.; Jiang, Z.J.; Cui, Y.F. Reactive oxygen species, but not mitochondrial membrane potential, is associated with radiation-induced apoptosis of AHH-1 human lymphoblastoid cells. Cell. Biol. Int. 2007, 31, 1353–1358. [Google Scholar] [CrossRef]
- Pathak, N.; Khandelwal, S. Modulation of cadmiuminducedalterations in murine thymocytes by piperine: Oxidative stress, apoptosis, phenotyping and blastogenesis. Biochem. Pharmacol. 2006, 72, 486–497. [Google Scholar]
- Yang, Z.; Wu, D.; Bui, T.; Ho, R.J. A novel human multidrug resistance gene MDR1 variant G571A (G191R) modulates cancer drug resistance and efflux transport. J. Pharmacol. Exp. Ther. 2008, 327, 474–481. [Google Scholar] [CrossRef]
- Samples Availability: Samples of the compounds are available from the authors.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cherneva, E.; Pavlovic, V.; Smelcerovic, A.; Yancheva, D. The Effect of Camphor and Borneol on Rat Thymocyte Viability and Oxidative Stress. Molecules 2012, 17, 10258-10266. https://doi.org/10.3390/molecules170910258
Cherneva E, Pavlovic V, Smelcerovic A, Yancheva D. The Effect of Camphor and Borneol on Rat Thymocyte Viability and Oxidative Stress. Molecules. 2012; 17(9):10258-10266. https://doi.org/10.3390/molecules170910258
Chicago/Turabian StyleCherneva, Emiliya, Voja Pavlovic, Andrija Smelcerovic, and Denitsa Yancheva. 2012. "The Effect of Camphor and Borneol on Rat Thymocyte Viability and Oxidative Stress" Molecules 17, no. 9: 10258-10266. https://doi.org/10.3390/molecules170910258