Specific Binding of Anionic Porphyrin and Phthalocyanine to the G-Quadruplex with a Variety of in Vitro and in Vivo Applications


Abstract
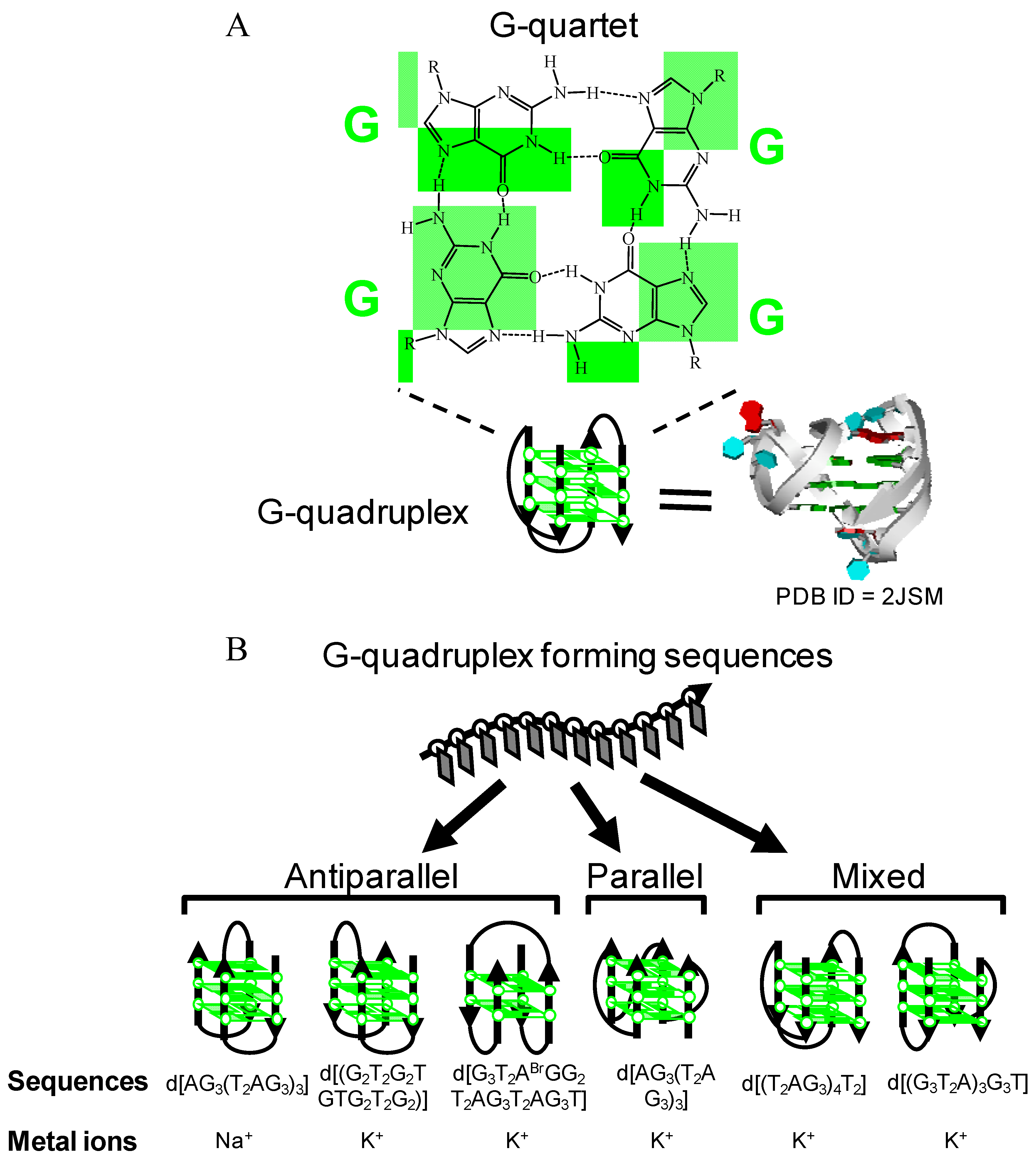
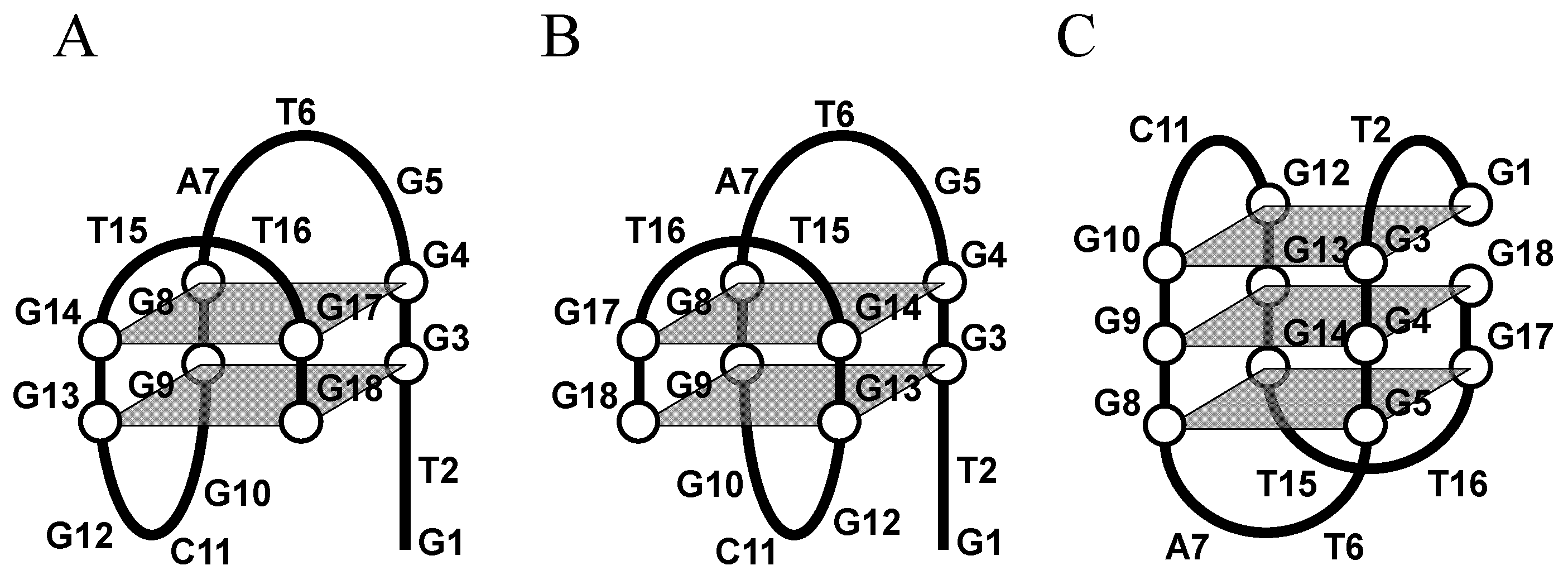
:1. Introduction



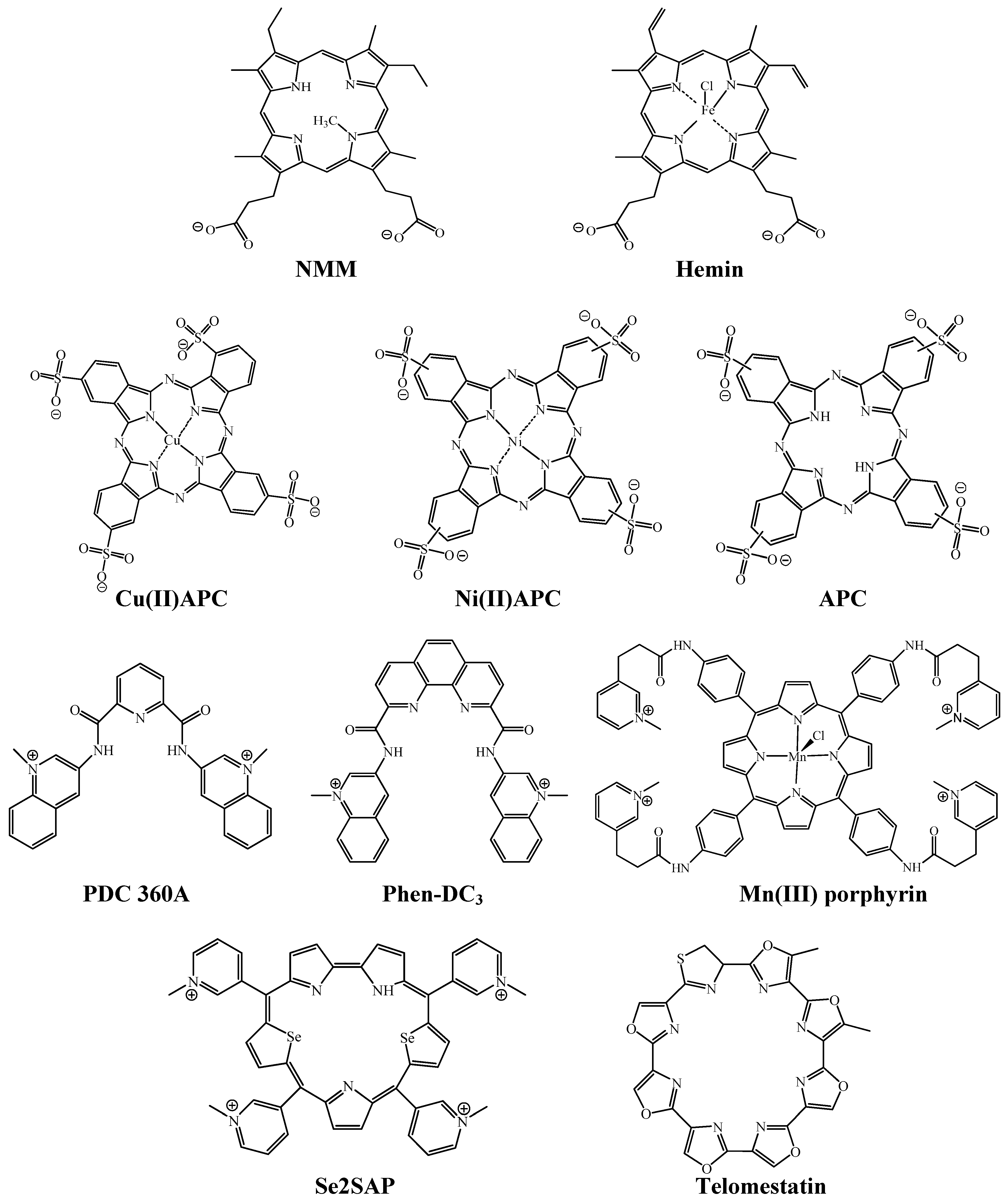
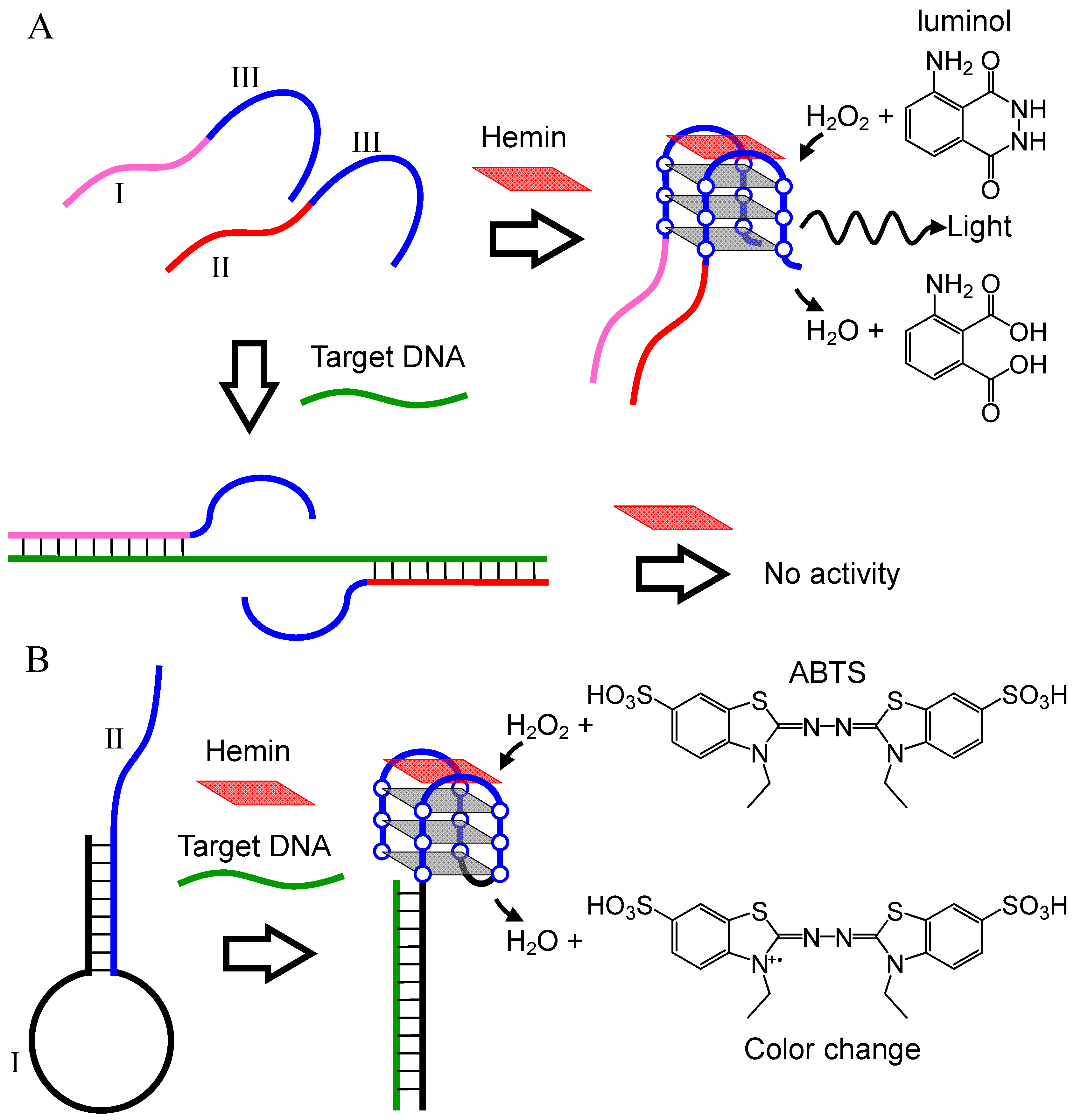
2. Hemin-G-quadruplex Interaction for in Vitro Applications
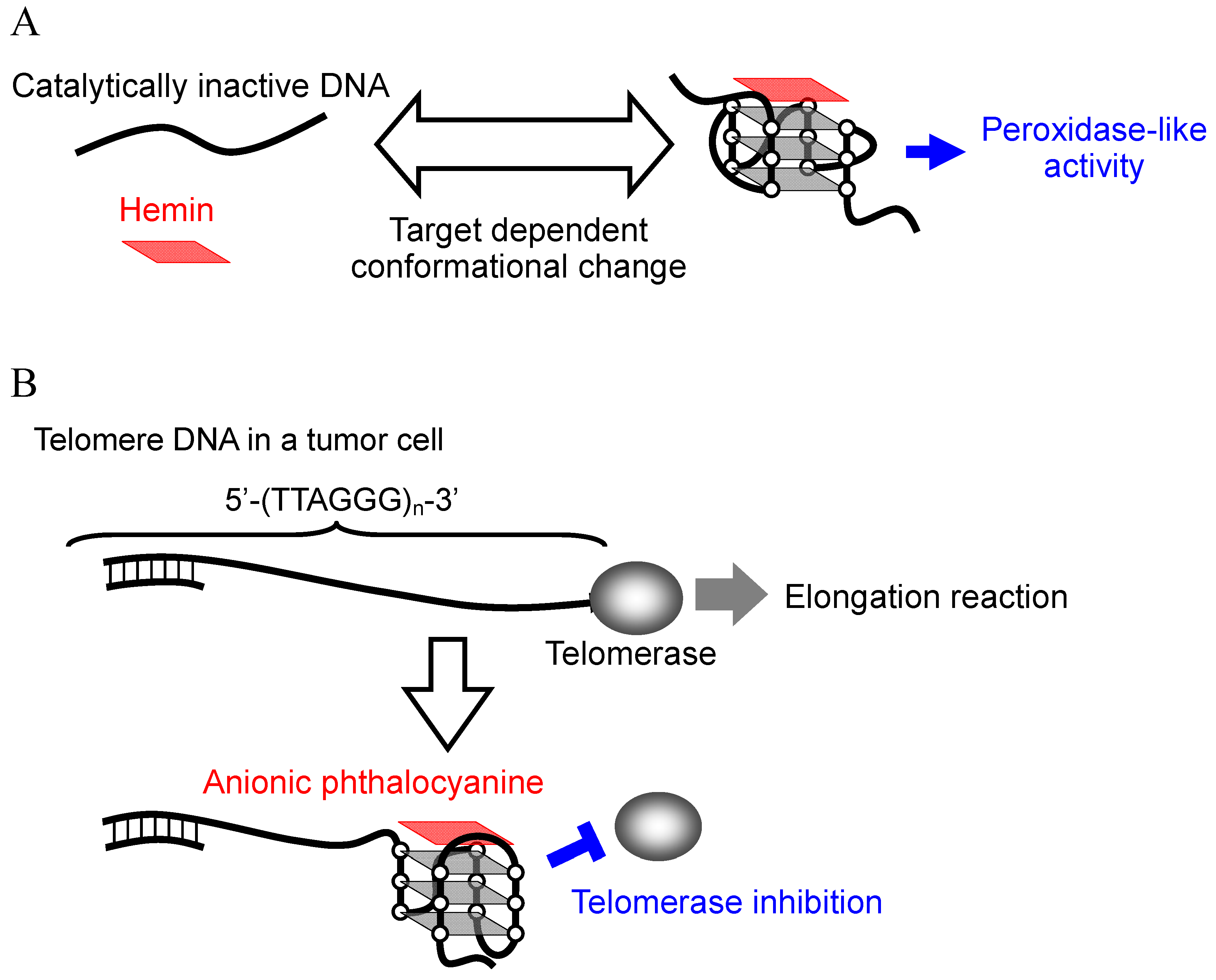
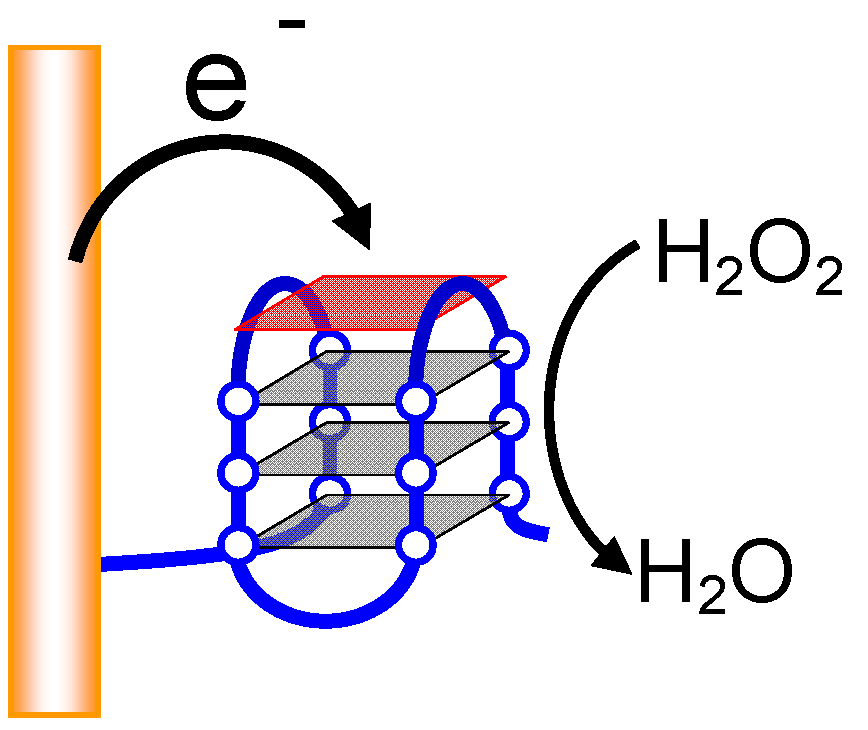
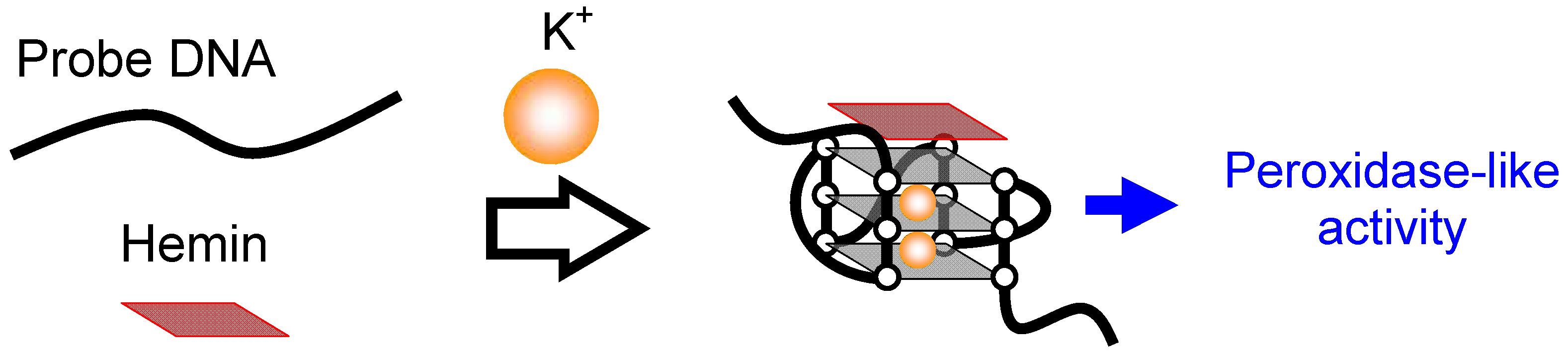
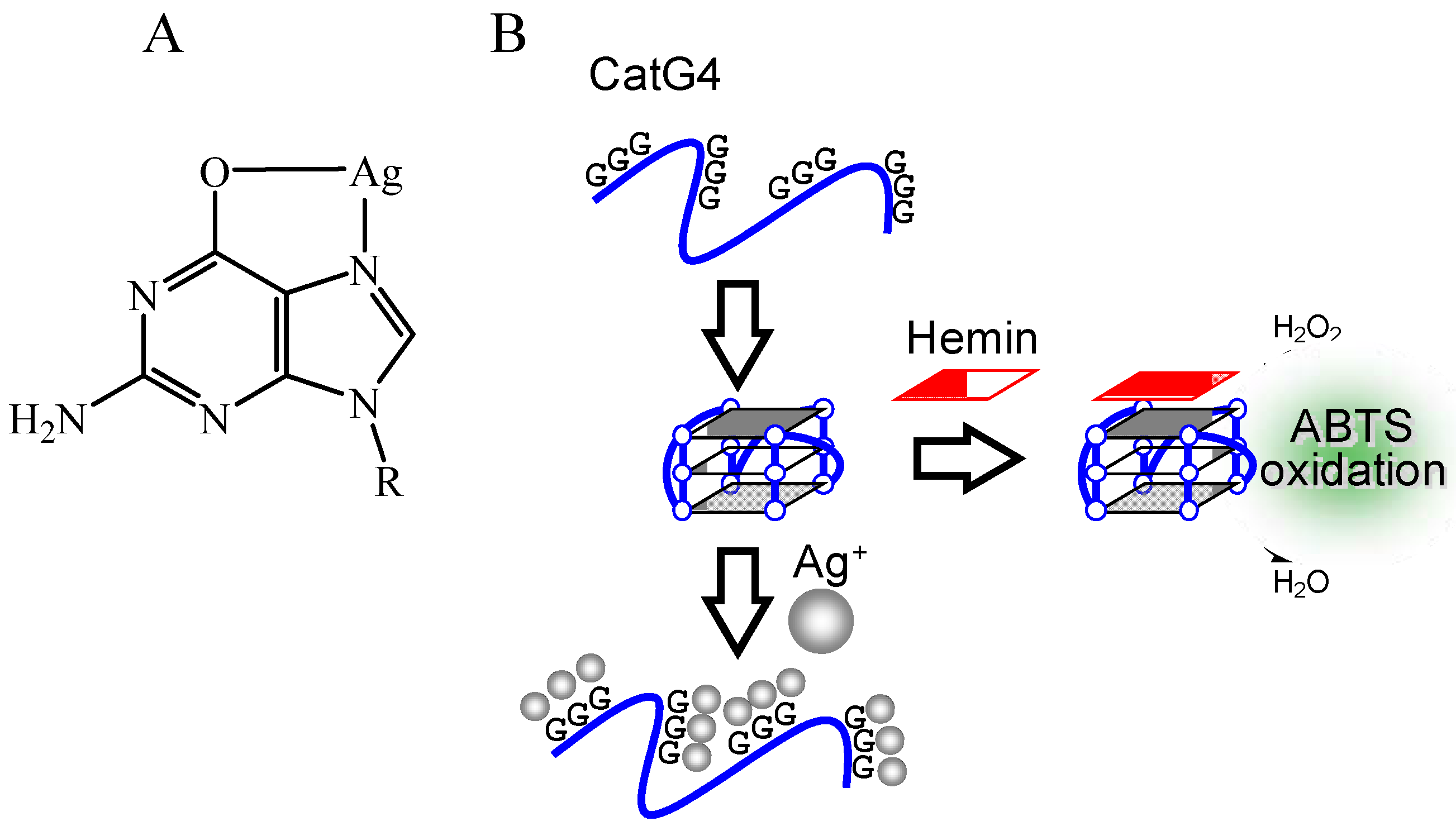
2.1. Mechanism of Induction of Peroxidase-Like Activity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequences (5'→3') |
|---|---|
| PS2 | TTGCC TAACC GTGAA GGTAA AACGA TTTAG TCAAA CGTGG GAGGG CGGTG GTGTT GACTG ATCGA TTTTA TTCCA |
| PS5 | GTGTC GAAGA TCGTG GGTCA TTGTG GGTGG GTGTG GCTGG TCCGA TCCGC GATCT GCTGA CGCTG GTTAG GT |
| PS2.M | GTGGG TAGGG CGGGT TGG |
| PS5.M | GTGGG TCATT GTGGG TGGGT GTGG |
| CatG4 | TGGGT AGGGC GGGTT GGGAA A |
| TBA | GGTTG GTGTG GTTGG |
| AGRO100 | GGTGG TGGTG GTTGT GGTGG TGGTG G |



2.2. DNA Detection
2.2.1. DNA Detection Utilizing Conformational Change of Probe DNA
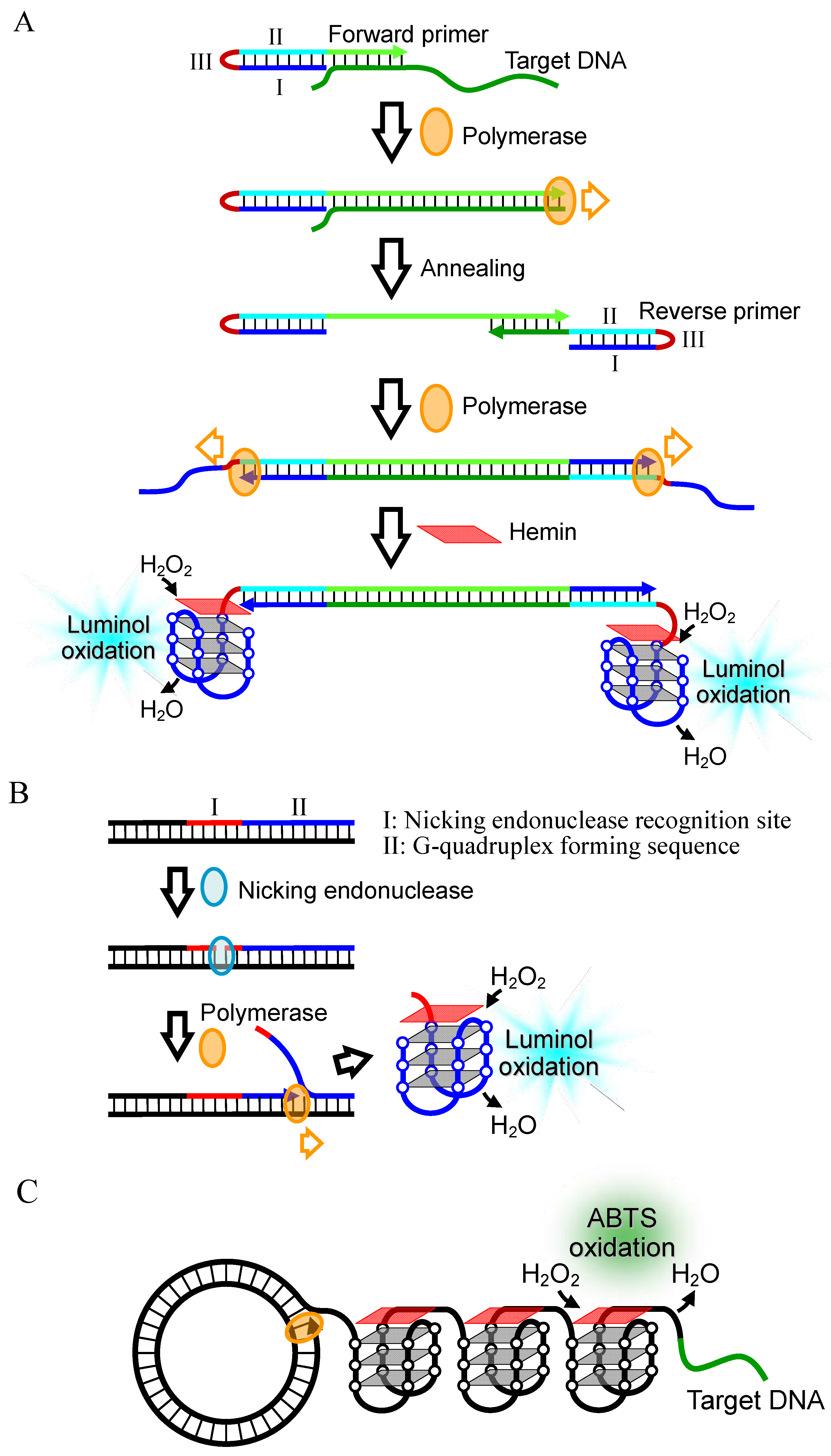
2.2.2. Signal Amplification by Polymerase Reaction

| Ref. | Principle | Target DNA | Indicator | Detection Limit | Experimental Conditions |
|---|---|---|---|---|---|
| [43] | Figure 7A | 5'-TTGAG CATGC GCATT ATCTG AGCCA GTACC GAATC G-3' | Luminol | 0.6 μM | 25 mM HEPES buffer, 20 mM KCl, 200 mM NaCl, 0.5 mM luminol, 30 mM H2O2, pH 9.0 a |
| [44] | Figure 7B | 5'-AATGG CAGCA ATTTC ACCAG TACTA CAGTT AAGGC-3' | ABTS | 0.2 μM | 0.1 M Tris buffer, 20 mM MgCl2, 0.43 μM hemin, 3.2 mM ABTS, 3.2 mM H2O2, pH 8.1 |
| [45] | Figure 8A | M13 phage DNA | Luminol | 1.2 aM | 25 mM HEPES buffer, 20 mM KCl, 200 mM NaCl, 1 nM hemin, 0.5 mM luminol, 30 mM H2O2, pH 9.0 |
| [46] | Figure 8B | M13 phage DNA | ABTS or luminol | 10 fM | Colorimetric measurements (ABTS) : 25 mM HEPES buffer, 20 mM KCl, 200 mM NaCl, 0.4 μM hemin, 182 μM ABTS, 44 μM H2O2, pH 7.4; Chemiluminescence measurements (luminol): 25 mM HEPES buffer, 20 mM KCl, 200 mM NaCl, 1 nM hemin, 0.5 mM luminol, 30 mM H2O2, pH 9.0 |
| [47] | Figure 8C | 5'-CTCAC ACGAA TTCAT CTGAC-3' | ABTS | 1 pM | 15 mM Tris-HCl, 3 mM MgCl2, 1 mM (NH4)2SO4, 0.01 mg/mL BSA, 0.005% Triton X-100, 0.5 μM hemin, 1.17 mM ABTS, 2.82 mM H2O2 |
| [48] | Figure 9A | 5'-CCTCC CGGTG TTCGA TCC-3' | Luminol | 76 aM | b |
| [49] | Figure 9B | 5'-TCGAA TAAGC ACTGA GGT-3'5'-ATAAA TTGCC AAGAT GAT-3'5'-TATCA ATACT CCCCC AGG-3' | CRET (luminol) | 10 nM | 25 mM HEPES buffer, 20 mM KNO3, 200 mM NaNO3, 10 nM hemin, pH 9.0 c |
| [50] | Figure 9C | 5'-TTGAG CATGC GCATT ATCTG AGCCA GTACC GAATC G-3' | Luminol | 100 pM | 25 mM HEPES buffer, 20 mM KCl, 200 mM NaCl, 0.5 mM luminol, 30 mM H2O2, pH 9.0 |
| [51] | Figure 10 | 5'-TCGAA TAAGC ACTGA GGT-3' | Electrocatalytic cathodic current | 1 pM | 10 mM HEPES buffer, 50 mM KCl, 150 mM NaCl, 1 μM hemin, 1 mM H2O2, pH 7.2 |

2.2.3. Combination with Nanoparticle

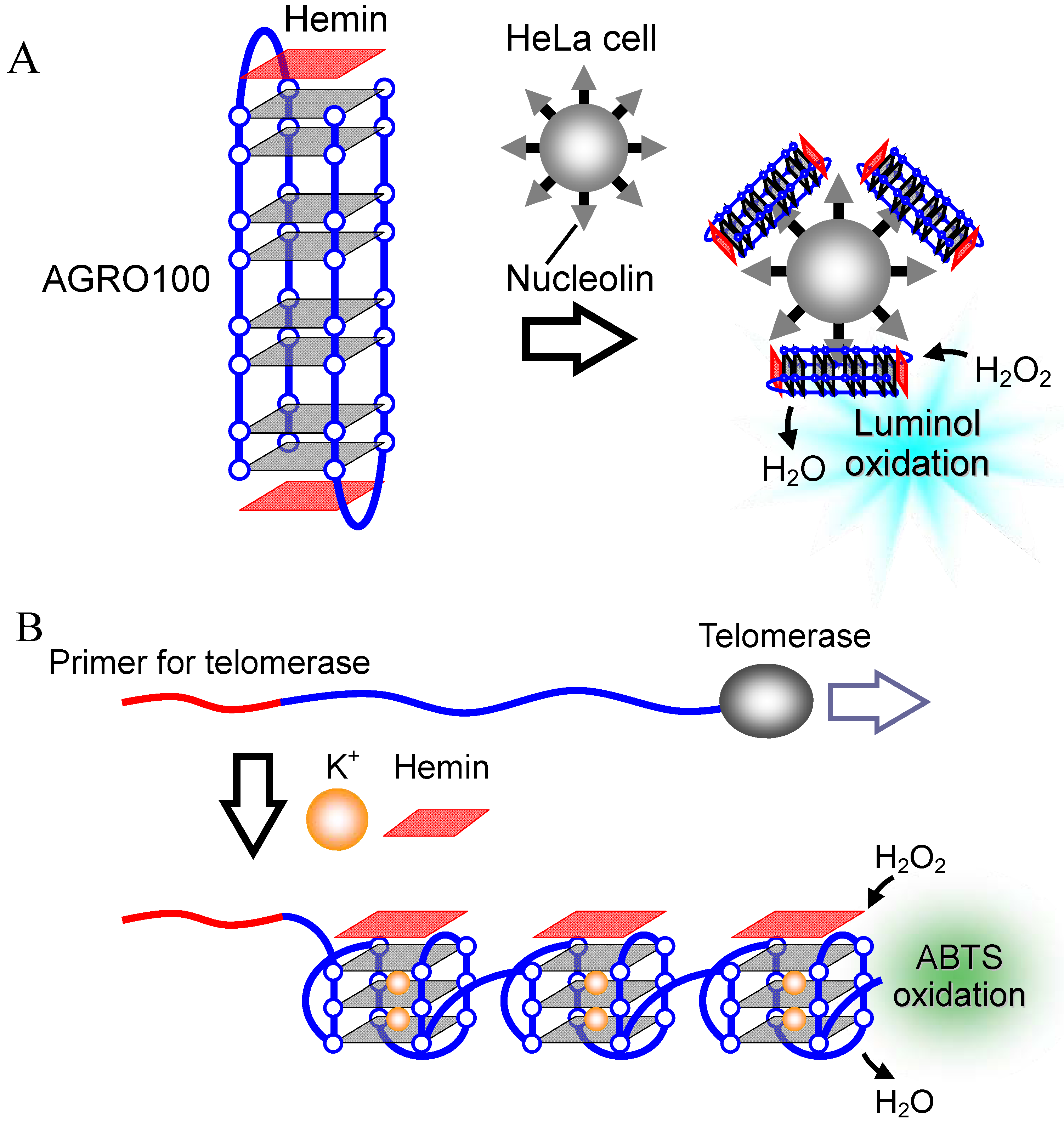
2.2.4. Electrochemical DNA Detection

2.3. Biosensors for Various Targets
2.3.1. Metal Ions



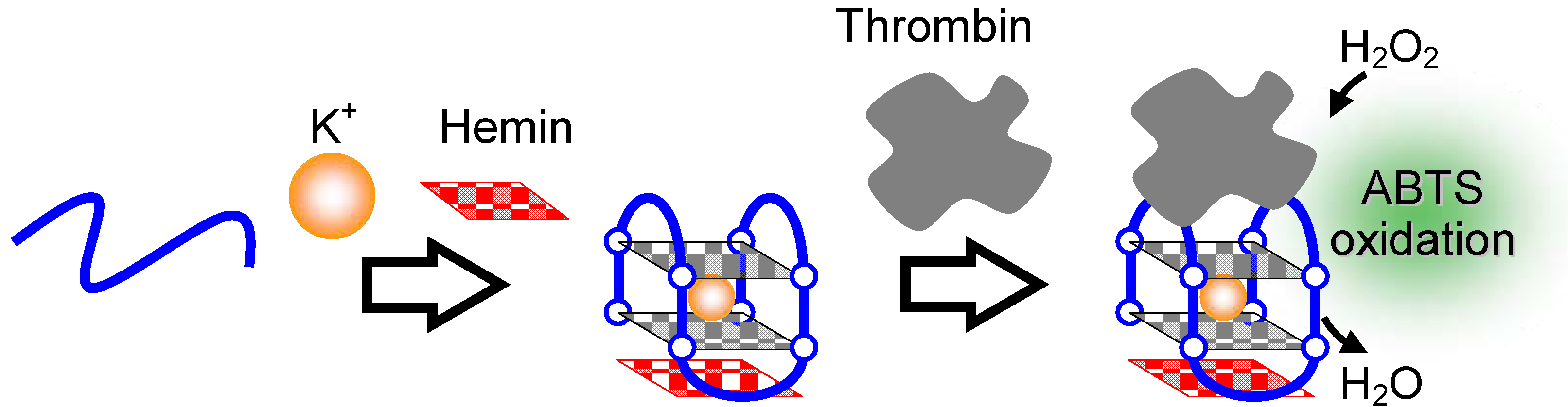
2.3.2. Proteins


3. Anionic Phthalocyanine-G-Quadruplex Interaction for in Vivo Applications
3.1. G-Quadruplex-Ligands as Telomerase Inhibitors
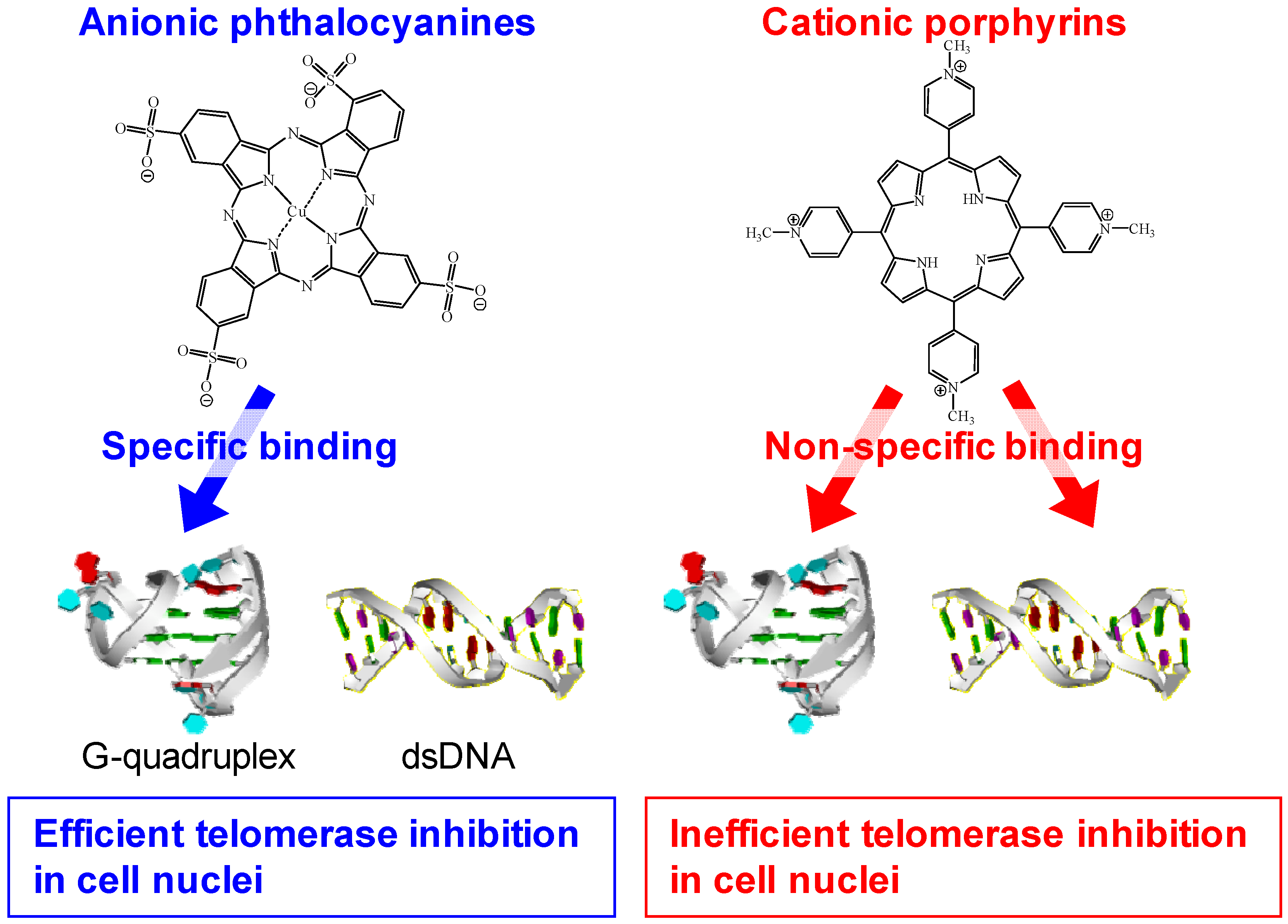
3.2. Selective Binding of Anionic Phthalocyanines to the Human Telomere G-quadruplex
| Kd/μM | IC50/μM | |||||||
|---|---|---|---|---|---|---|---|---|
| G-quadruplex a (−Lambda DNA) | ssDNA b | dsDNA c | G-quadruplex (+Lambda DNA) | −Lambda DNA | +Lambda DNA | |||
| Cu(II)APC | 42 | >250 | >250 | 36 | 1.2 | 1.2 | ||
| Ni(II)APC | 56 | >250 | >250 | 71 | 1.9 | 1.4 | ||
| APC | - | - | - | - | 2.4 | 2.3 | ||
| TMPyP4 | 0.7 | - | - | No binding d | 3.7 | No inhibition e | ||
3.3. Telomerase Inhibition by Anionic Phthalocyanines

4. Summary and Outlook

Acknowledgments
References
- Endo, M.; Shiroyama, T.; Fujitsuka, M.; Majima, T. Four-way-branched DNA-porphyrin conjugates for construction of four double-helix-DNA assembled structures. J. Org. Chem. 2005, 70, 7468–7472. [Google Scholar] [CrossRef]
- Liu, X.J.; Qi, C.; Bing, T.; Cheng, X.H.; Shangguan, D.H. Highly selective phthalocyanine-thymine conjugate sensor for Hg2+ based on target induced aggregation. Anal. Chem. 2009, 81, 3699–3704. [Google Scholar]
- Hammer, R.P.; Owens, C.V.; Hwang, S.H.; Sayes, C.M.; Soper, S.A. Asymmetrical, water-soluble phthalocyanine dyes for covalent labeling of oligonucleotides. Bioconjug.Chem. 2002, 13, 1244–1252. [Google Scholar] [CrossRef]
- Bianco, A.; Da Ros, T.; Prato, M.; Toniolo, C. Fullerene-based amino acids and peptides. J. Pept. Sci. 2001, 7, 208–219. [Google Scholar] [CrossRef]
- Yang, W.R.; Thordarson, P.; Gooding, J.J.; Ringer, S.P.; Braet, F. Carbon nanotubes for biological and biomedical applications. Nanotechnology 2007, 18, 412001–412012. [Google Scholar] [CrossRef]
- Wang, H.; Yang, R.H.; Yang, L.; Tan, W.H. Nucleic acid conjugated nanomaterials for enhanced molecular recognition. ACS Nano 2009, 3, 2451–2460. [Google Scholar] [CrossRef]
- Zhang, S.G. Fabrication of novel biomaterials through molecular self-assembly. Nat. Biotechnol. 2003, 21, 1171–1178. [Google Scholar] [CrossRef]
- Wang, Z.D.; Lu, Y. Functional DNA directed assembly of nanomaterials for biosensing. J. Mater. Chem. 2009, 19, 1788–1798. [Google Scholar] [CrossRef]
- Endo, M.; Sugiyama, H. Chemical approaches to DNA nanotechnology. ChemBioChem 2009, 10, 2420–2443. [Google Scholar] [CrossRef]
- Gellert, M.; Lipsett, M.N.; Davies, D.R. Helix formation by guanylic acid. Proc. Natl. Acad. Sci. USA 1962, 48, 2013–2018. [Google Scholar] [CrossRef]
- Keniry, M.A. Quadruplex structures in nucleic acids. Biopolymers 2001, 56, 123–146. [Google Scholar] [CrossRef]
- Davis, J.T. G-quartets 40 years later: From 5'-GMP to molecular biology and supramolecular chemistry. Angew. Chem. Int. Ed.Engl. 2004, 43, 668–698. [Google Scholar] [CrossRef]
- Dai, J.X.; Carver, M.; Yang, D.Z. Polymorphism of human telomeric quadruplex structures. Biochimie 2008, 90, 1172–1183. [Google Scholar] [CrossRef]
- Kaushik, M.; Kaushik, S.; Bansal, A.; Saxena, S.; Kukreti, S. Structural diversity and specific recognition of four stranded G-quadruplex DNA. Curr. Mol. Med. 2011, 11, 744–769. [Google Scholar] [CrossRef]
- Willner, I.; Shlyahovsky, B.; Zayats, M.; Willner, B. DNAzymes for sensing, nanobiotechnology and logic gate applications. Chem. Soc. Rev. 2008, 37, 1153–1165. [Google Scholar]
- Liu, J.W.; Cao, Z.H.; Lu, Y. Functional nucleic acid sensors. Chem. Rev. 2009, 109, 1948–1998. [Google Scholar] [CrossRef]
- Kosman, J.; Juskowiak, B. Peroxidase-mimicking DNAzymes for biosensing applications: A review. Anal. Chim. Acta 2011, 707, 7–17. [Google Scholar] [CrossRef]
- Sen, D.; Poon, L.C.H. RNA and DNA complexes with hemin Fe(III) heme are efficient peroxidases and peroxygenases: How do they do it and what does it mean? Crit. Rev. Biochem. Mol. 2011, 46, 478–492. [Google Scholar]
- Wheelhouse, R.T.; Sun, D.K.; Han, H.Y.; Han, F.X.G.; Hurley, L.H. Cationic porphyrins as telomerase inhibitors: The interaction of tetra-(N-methyl-4-pyridyl)porphine with quadruplex DNA. J. Am. Chem. Soc. 1998, 120, 3261–3262. [Google Scholar] [CrossRef]
- Monchaud, D.; Teulade-Fichou, M.P. A hitchhiker’s guide to G-quadruplex ligands. Org. Biomol. Chem. 2008, 6, 627–636. [Google Scholar] [CrossRef]
- De Cian, A.; Lacroix, L.; Douarre, C.; Temime-Smaali, N.; Trentesaux, C.; Riou, J.F.; Mergny, J.L. Targeting telomeres and telomerase. Biochimie 2008, 90, 131–155. [Google Scholar] [CrossRef]
- Luedtke, N.W. Targeting G-quadruplex DNA with small molecules. Chimia 2009, 63, 134–139. [Google Scholar] [CrossRef]
- Neidle, S. Human telomeric G-quadruplex: The current status of telomeric G-quadruplexes as therapeutic targets in human cancer. FEBS J. 2010, 277, 1118–1125. [Google Scholar] [CrossRef]
- Ou, T.M.; Lu, Y.J.; Tan, J.H.; Huang, Z.S.; Wong, K.Y.; Gu, L.Q. G-quadruplexes: Targets in anticancer drug design. ChemMedChem 2008, 3, 690–713. [Google Scholar] [CrossRef]
- Yaku, H.; Murashima, T.; Miyoshi, D.; Sugimoto, N. Anionic phthalocyanines targeting G-quadruplexes and inhibiting telomerase activity in the presence of excessive DNA duplexes. Chem. Commun. 2010, 46, 5740–5742. [Google Scholar]
- Yaku, H.; Fujimoto, T.; Murashima, T.; Miyoshi, D.; Sugimoto, N. Phthalocyanines: A new class of G-quadruplex-ligands with many potential applications. Chem. Commun. 2012, 48, 6203–6216. [Google Scholar]
- Travascio, P.; Li, Y.F.; Sen, D. DNA-enhanced peroxidase activity of a DNA aptamer-hemin complex. Chem. Biol. 1998, 5, 505–517. [Google Scholar] [CrossRef]
- Moyzis, R.K.; Buckingham, J.M.; Cram, L.S.; Dani, M.; Deaven, L.L.; Jones, M.D.; Meyne, J.; Ratliff, R.L.; Wu, J.R. A highly conserved repetitive DNA-sequence, (TTAGGG)n, present at the telomeres of human-chromosomes. Proc. Natl. Acad. Sci. USA 1988, 85, 6622–6626. [Google Scholar]
- Harley, C.B.; Futcher, A.B.; Greider, C.W. Telomeres shorten during ageing of human fibroblasts. Nature 1990, 345, 458–460. [Google Scholar]
- Harley, C.B.; Vaziri, H.; Counter, C.M.; Allsopp, R.C. The telomere hypothesis of cellular aging. Exp. Gerontol. 1992, 27, 375–382. [Google Scholar] [CrossRef]
- Greider, C.W.; Blackburn, E.H. Identification of a specific telomere terminal transferase-activity in tetrahymena extracts. Cell 1985, 43, 405–413. [Google Scholar] [CrossRef]
- Morin, G.B. The human telomere terminal transferase enzyme is a ribonucleoprotein that synthesizes TTAGGG repeats. Cell 1989, 59, 521–529. [Google Scholar] [CrossRef]
- Zahler, A.M.; Williamson, J.R.; Cech, T.R.; Prescott, D.M. Inhibition of telomerase by G-quartet DNA structures. Nature 1991, 350, 718–720. [Google Scholar]
- Travascio, P.; Bennet, A.J.; Wang, D.Y.; Sen, D. A ribozyme and a catalytic DNA with peroxidase activity: Active sites versus cofactor-binding sites. Chem. Biol. 1999, 6, 779–787. [Google Scholar] [CrossRef]
- Li, Y.F.; Geyer, C.R.; Sen, D. Recognition of anionic porphyrins by DNA aptamers. Biochemistry 1996, 35, 6911–6922. [Google Scholar] [CrossRef]
- Li, Y.F.; Sen, D. A catalytic DNA for porphyrin metallation. Nat. Struct. Biol. 1996, 3, 743–747. [Google Scholar] [CrossRef]
- Li, Y.F.; Sen, D. Toward an efficient DNAzyme. Biochemistry 1997, 36, 5589–5599. [Google Scholar] [CrossRef]
- Lee, H.W.; Chinnapen, D.J.F.; Sen, D. Structure-function investigation of a deoxyribozyme with dual chelatase and peroxidase activities. Pure Appl. Chem. 2004, 76, 1537–1545. [Google Scholar] [CrossRef]
- Majhi, P.R.; Shafer, R.H. Characterization of an unusual folding pattern in a catalytically active guanine quadruplex structure. Biopolymers 2006, 82, 558–569. [Google Scholar] [CrossRef]
- Kong, D.M.; Cai, L.L.; Guo, J.H.; Wu, J.; Shen, H.X. Characterization of the G-quadruplex structure of a catalytic DNA with peroxidase activity. Biopolymers 2009, 91, 331–339. [Google Scholar] [CrossRef]
- Cheng, X.H.; Liu, X.J.; Bing, T.; Cao, Z.H.; Shangguan, D.H. General peroxidase activity of G-quadruplex-hemin complexes and its application in ligand screening. Biochemistry 2009, 48, 7817–7823. [Google Scholar] [CrossRef]
- Kong, D.M.; Yang, W.; Wu, J.; Li, C.X.; Shen, H.X. Structure-function study of peroxidase-like G-quadruplex-hemin complexes. Analyst 2010, 135, 321–326. [Google Scholar] [CrossRef]
- Yi, X.; Pavlov, V.; Gill, R.; Bourenko, T.; Willner, I. Lighting up biochemiluminescence by the surface self-assembly of DNA-hemin complexes. ChemBioChem 2004, 5, 374–379. [Google Scholar] [CrossRef]
- Xiao, Y.; Pavlov, V.; Niazov, T.; Dishon, A.; Kotler, M.; Willner, I. Catalytic beacons for the detection of DNA and telomerase activity. J. Am. Chem. Soc. 2004, 126, 7430–7431. [Google Scholar]
- Cheglakov, Z.; Weizmann, Y.; Beissenhirtz, M.K.; Willner, I. Ultrasensitive detection of DNA by the PCR-induced generation of DNAzymes: The DNAzyme primer approach. Chem. Commun. 2006, 3205–3207. [Google Scholar]
- Weizmann, Y.; Beissenhirtz, M.K.; Cheglakov, Z.; Nowarski, R.; Kotler, M.; Willner, I. A virus spotlighted by an autonomous DNA machine. Angew. Chem. Int. Ed.Engl. 2006, 45, 7384–7388. [Google Scholar] [CrossRef]
- Tian, Y.; He, Y.; Mao, C.D. Cascade signal amplification for DNA detection. ChemBioChem 2006, 7, 1862–1864. [Google Scholar] [CrossRef]
- Bi, S.; Zhang, J.L.; Zhang, S.S. Ultrasensitive and selective DNA detection based on nicking endonuclease assisted signal amplification and its application in cancer cell detection. Chem. Commun. 2010, 46, 5509–5511. [Google Scholar]
- Freeman, R.; Liu, X.Q.; Winner, I. Chemiluminescent and chemiluminescence resonance energy transfer (CRET) detection of DNA, metal ions, and aptamer-substrate complexes using hemin/G-quadruplexes and CdSe/ZnS quantum dots. J. Am. Chem. Soc. 2011, 133, 11597–11604. [Google Scholar]
- Niazov, T.; Pavlov, V.; Xiao, Y.; Gill, R.; Willner, I. DNAzyme-functionalized Au nanoparticles for the amplified detection of DNA or telomerase activity. Nano Lett. 2004, 4, 1683–1687. [Google Scholar] [CrossRef]
- Pelossof, G.; Tel-Vered, R.; Elbaz, J.; Willner, I. Amplified biosensing using the horseradish peroxidase-mimicking DNAzyme as an electrocatalyst. Anal. Chem. 2010, 82, 4396–4402. [Google Scholar] [CrossRef]
- Yang, Q.L.; Nie, Y.J.; Zhu, X.L.; Liu, X.J.; Li, G.X. Study on the electrocatalytic activity of human telomere G-quadruplex-hemin complex and its interaction with small molecular ligands. Electrochim. Acta 2009, 55, 276–280. [Google Scholar] [CrossRef]
- Teixeira, M.F.S.; Freitas, B.H.; Seraphim, P.M.; Salmazo, L.O.; Nobre, M.A.; Lanfredi, S. Development of an electrochemical sensor for potassium ions based on KSr2Nb5O15 modified electrode. Proced. Chem. 2009, 1, 293–296. [Google Scholar] [CrossRef]
- Fan, X.Y.; Li, H.T.; Zhao, J.; Lin, F.; Zhang, L.; Zhang, Y.; Yao, S. A novel label-free fluorescent sensor for the detection of potassium ion based on DNAzyme. Talanta 2012, 89, 57–62. [Google Scholar] [CrossRef]
- Ratte, H.T. Bioaccumulation and toxicity of silver compounds: A review. Environ. Toxicol. Chem. 1999, 18, 89–108. [Google Scholar] [CrossRef]
- Jarup, L. Hazards of heavy metal contamination. Br. Med. Bull. 2003, 68, 167–182. [Google Scholar] [CrossRef]
- Li, T.; Wang, E.; Dong, S.J. G-quadruplex-based DNAzyme as a sensing platform for ultrasensitive colorimetric potassium detection. Chem. Commun. 2009, 580–582. [Google Scholar]
- Yang, X.; Li, T.; Li, B.L.; Wang, E.K. Potassium-sensitive G-quadruplex DNA for sensitive visible potassium detection. Analyst 2010, 135, 71–75. [Google Scholar] [CrossRef]
- Li, T.; Wang, E.; Dong, S. Lead(II)-induced allosteric G-quadruplex DNAzyme as a colorimetric and chemiluminescence sensor for highly sensitive and selective Pb2+ detection. Anal. Chem. 2010, 82, 1515–1520. [Google Scholar] [CrossRef]
- Li, C.L.; Liu, K.T.; Lin, Y.W.; Chang, H.T. Fluorescence detection of lead(II) ions through their induced catalytic activity of DNAzymes. Anal. Chem. 2011, 83, 225–230. [Google Scholar] [CrossRef]
- Zhang, J.; Gao, Q.L.; Chen, P.P.; Chen, J.H.; Chen, G.N.; Fu, F.F. A novel Tb3+-promoted G-quadruplex-hemin DNAzyme for the development of label-free visual biosensors. Biosens. Bioelectron. 2011, 26, 4053–4057. [Google Scholar] [CrossRef]
- Li, T.; Dong, S.J.; Wang, E. Label-free colorimetric detection of aqueous mercury ion (Hg2+) using Hg2+-modulated G-quadruplex-cased DNAzymes. Anal. Chem. 2009, 81, 2144–2149. [Google Scholar] [CrossRef]
- Kong, D.M.; Wu, J.; Wang, N.; Yang, W.; Shen, H.X. Peroxidase activity-structure relationship of the intermolecular four-stranded G-quadruplex-hemin complexes and their application in Hg2+ ion detection. Talanta 2009, 80, 459–465. [Google Scholar] [CrossRef]
- Li, T.; Li, B.L.; Wang, E.K.; Dong, S.J. G-quadruplex-based DNAzyme for sensitive mercury detection with the naked eye. Chem. Commun. 2009, 3551–3553. [Google Scholar]
- Jia, S.M.; Liu, X.F.; Li, P.; Kong, D.M.; Shen, H.X. G-quadruplex DNAzyme-based Hg2+ and cysteine sensors utilizing Hg2+-mediated oligonucleotide switching. Biosens. Bioelectron. 2011, 27, 148–152. [Google Scholar] [CrossRef]
- Zhou, X.H.; Kong, D.M.; Shen, H.X. Ag+ and cysteine quantitation based on G-quadruplex-hemin DNAzymes disruption by Ag+. Anal. Chem. 2010, 82, 789–793. [Google Scholar] [CrossRef]
- Zhou, X.H.; Kong, D.M.; Shen, H.X. G-quadruplex-hemin DNAzyme-amplified colorimetric detection of Ag+ ion. Anal. Chim. Acta 2010, 678, 124–127. [Google Scholar] [CrossRef]
- Tanaka, K.; Tengeiji, A.; Kato, T.; Toyama, N.; Shiro, M.; Shionoya, M. Efficient incorporation of a copper hydroxypyridone base pair in DNA. J. Am. Chem. Soc. 2002, 124, 12494–12498. [Google Scholar]
- Tanaka, K.; Yamada, Y.; Shionoya, M. Formation of silver(I)-mediated DNA duplex and triplex through an alternative base pair of pyridine nucleobases. J. Am. Chem. Soc. 2002, 124, 8802–8803. [Google Scholar]
- Tanaka, K.; Tengeiji, A.; Kato, T.; Toyama, N.; Shionoya, M. A discrete self-assembled metal array in artificial DNA. Science 2003, 299, 1212–1213. [Google Scholar] [CrossRef]
- Tanaka, K.; Clever, G.H.; Takezawa, Y.; Yamada, Y.; Kaul, C.; Shionoya, M.; Carell, T. Programmable self-assembly of metal ions inside artificial DNA duplexes. Nat. Nanotechnol. 2006, 1, 190–194. [Google Scholar] [CrossRef]
- Muller, J. Chemistry: Metals line up for DNA. Nature 2006, 444, 698–698. [Google Scholar] [CrossRef]
- Takezawa, Y.; Maeda, W.; Tanaka, K.; Shionoya, M. Discrete Self-Assembly of iron(III) ions inside triple-stranded artificial DNA. Angew. Chem. Int. Ed.Engl. 2009, 48, 1081–1084. [Google Scholar]
- Bock, L.C.; Griffin, L.C.; Latham, J.A.; Vermaas, E.H.; Toole, J.J. Selection of single-stranded-DNA molecules that bind and inhibit human thrombin. Nature 1992, 355, 564–566. [Google Scholar]
- Macaya, R.F.; Schultze, P.; Smith, F.W.; Roe, J.A.; Feigon, J. Thrombin-binding DNA aptamer forms a unimolecular quadruplex structure in solution. Proc. Natl. Acad. Sci. USA 1993, 90, 3745–3749. [Google Scholar] [CrossRef]
- Li, T.; Wang, E.K.; Dong, S.J. G-quadruplex-based DNAzyme for facile colorimetric detection of thrombin. Chem. Commun. 2008, 3654–3656. [Google Scholar]
- Shen, B.J.; Wang, Q.; Zhu, D.; Luo, J.J.; Cheng, G.F.; He, P.A.; Fang, Y.Z. G-quadruplex-based DNAzymes aptasensor for the amplified electrochemical detection of thrombin. Electroanalysis 2010, 22, 2985–2990. [Google Scholar] [CrossRef]
- Girvan, A.C.; Teng, Y.; Casson, L.K.; Thomas, S.D.; Jüliger, S.; Ball, M.W.; Klein, J.B.; Pierce, W.M., Jr.; Barve, S.S.; Bates, P.J. AGRO100 inhibits activation of nuclear factor-kappa B (NF-kappa B) by forming a complex with NF-kappa B essential modulator (NEMO) and nucleolin. Mol. Cancer Ther. 2006, 5, 1790–1799. [Google Scholar] [CrossRef]
- Ireson, C.R.; Kelland, L.R. Discovery and development of anticancer aptamers. Mol. Cancer Ther. 2006, 5, 2957–2962. [Google Scholar] [CrossRef]
- Li, T.; Shi, L.L.; Wang, E.K.; Dong, S.J. Multifunctional G-quadruplex aptamers and their application to protein detection. Chem.-Eur. J. 2009, 15, 1036–1042. [Google Scholar] [CrossRef]
- He, Y.; Tian, Y.; Mao, C.D. Human telomeric DNA sequences have a peroxidase apoenzyme activity. Mol. Biosyst. 2009, 5, 238–240. [Google Scholar] [CrossRef]
- Freeman, R.; Sharon, E.; Teller, C.; Henning, A.; Tzfati, Y.; Willner, I. DNAzyme-like activity of hemin-telomeric G-quadruplexes for the optical analysis of telomerase and its inhibitors. ChemBioChem 2010, 11, 2362–2367. [Google Scholar] [CrossRef]
- Stefan, L.; Denat, F.; Monchaud, D. Deciphering the DNAzyme activity of multimeric quadruplexes: Insights into their actual role in the telomerase activity evaluation assay. J. Am. Chem. Soc. 2011, 133, 20405–20415. [Google Scholar] [CrossRef]
- Kosman, J.; Juskowiak, B. Optimization study of the catalytic activity of DNAzymes based on telomeric G-quadruplexes. Cent. Eur. J. Chem. 2012, 10, 368–372. [Google Scholar] [CrossRef]
- Izbicka, E.; Wheelhouse, R.T.; Raymond, E.; Davidson, K.K.; Lawrence, R.A.; Sun, D.; Windle, B.E.; Hurley, L.H.; Von Hoff, D.D. Effects of cationic porphyrins as G-quadruplex interactive agents in human tumor cells. Cancer Res. 1999, 59, 639–644. [Google Scholar]
- Rha, S.Y.; Izbicka, E.; Lawrence, R.A.; Davidson, K.K.; Sun, D.; Moyer, M.P.; Roodman, G.D.; Hurley, L.H.; Von Hoff, D.D. Effect of telomere and telomerase interactive agents on human tumor and normal cell lines. Clin. Cancer Res. 2000, 6, 987–993. [Google Scholar]
- Kim, M.Y.; Gleason-Guzman, M.; Izbicka, E.; Nishioka, D.; Hurley, L.H. The different biological effects of telomestatin and TMPyP4 can be attributed to their selectivity for interaction with intramolecular or intermolecular G-quadruplex structures. Cancer Res. 2003, 63, 3247–3256. [Google Scholar]
- Liu, W.; Sun, D.; Hurley, L.H. Binding of G-quadruplex-interactive agents to distinct G-quadruplexes induces different biological effects in MiaPaCa cells. Nucleosides Nucleotides Nucleic Acids 2005, 24, 1801–1815. [Google Scholar] [CrossRef]
- Mikami-Terao, Y.; Akiyama, M.; Yuza, Y.; Yanagisawa, T.; Yamada, O.; Yamada, H. Antitumor activity of G-quadruplex-interactive agent TMPyP4 in K562 leukemic cells. Cancer Lett. 2008, 261, 226–234. [Google Scholar] [CrossRef]
- Mikami-Terao, Y.; Akiyama, M.; Yuza, Y.; Yanagisawa, T.; Yamada, O.; Kawano, T.; Agawa, M.; Ida, H.; Yamada, H. Antitumor activity of TMPyP4 interacting G-quadruplex in retinoblastoma cell lines. Exp. Eye Res. 2009, 89, 200–208. [Google Scholar] [CrossRef]
- Gunaratnam, M.; Swank, S.; Haider, S.M.; Galesa, K.; Reszka, A.P.; Beltran, M.; Cuenca, F.; Fletcher, J.A.; Neidle, S. Targeting human gastrointestinal stromal tumor cells with a quadruplex-binding small molecule. J. Med. Chem. 2009, 52, 3774–3783. [Google Scholar] [CrossRef]
- Arthanari, H.; Basu, S.; Kawano, T.L.; Bolton, P.H. Fluorescent dyes specific for quadruplex DNA. Nucleic Acids Res. 1998, 26, 3724–3728. [Google Scholar] [CrossRef]
- Ren, J.S.; Chaires, J.B. Sequence and structural selectivity of nucleic acid binding ligands. Biochemistry 1999, 38, 16067–16075. [Google Scholar] [CrossRef]
- Kim, N.W.; Piatyszek, M.A.; Prowse, K.R.; Harley, C.B.; West, M.D.; Ho, P.L.; Coviello, G.M.; Weinrich, S.L.; Shay, J.W. Specific association of human telomerase activity with immortal cells and cancer. Science 1994, 266, 2011–2015. [Google Scholar]
- Yu, H.Q.; Zhang, D.H.; Gu, X.B.; Miyoshi, D.; Sugimoto, N. Regulation of telomerase activity by the thermodynamic stability of a DNA·RNA hybrid. Angew. Chem. Int. Ed.Engl. 2008, 47, 9034–9038. [Google Scholar]
- Shlyahovsky, B.; Li, Y.; Lioubashevski, O.; Elbaz, J.; Willner, I. Logic gates and antisense DNA devices operating on a translator nucleic acid scaffold. ACS Nano 2009, 3, 1831–1843. [Google Scholar] [CrossRef]
- Li, T.; Wang, E.K.; Dong, S.J. Potassium-lead-switched G-quadruplexes: A new class of DNA logic gates. J. Am. Chem. Soc. 2009, 131, 15082–15083. [Google Scholar]
- Li, T.; Ackermann, D.; Hall, A.M.; Famulok, M. Input-dependent induction of oligonucleotide structural motifs for performing molecular logic. J. Am. Chem. Soc. 2012, 134, 3508–3516. [Google Scholar]
- Rojas, A.M.; Gonzalez, P.A.; Antipov, E.; Klibanov, A.M. Specificity of a DNA-based (DNAzyme) peroxidative biocatalyst. Biotechnol. Lett. 2007, 29, 227–232. [Google Scholar] [CrossRef]
- Golub, E.; Freeman, R.; Willner, I. A hemin/G-quadruplex acts as an NADH oxidase and NADH peroxidase mimicking DNAzyme. Angew. Chem. Int. Ed. Engl. 2011, 50, 11710–11714. [Google Scholar] [CrossRef]
- Nakayama, S.; Wang, J.X.; Sintim, H.O. DNA-based peroxidation catalyst—What is the exact role of topology on catalysis and is there a special binding site for catalysis? Chem.-Eur. J. 2011, 17, 5691–5698. [Google Scholar] [CrossRef]
- Yang, X.J.; Fang, C.L.; Mei, H.C.; Chang, T.J.; Cao, Z.H.; Shangguan, D.H. Characterization of G-quadruplex/hemin peroxidase: Substrate specificity and inactivation kinetics. Chem.-Eur. J. 2011, 17, 14475–14484. [Google Scholar]
- Poon, L.C.H.; Methot, S.P.; Morabi-Pazooki, W.; Pio, P.; Bennet, A.J.; Sen, D. Guanine-rich RNAs and DNAs that bind heme robustly catalyze oxygen transfer reactions. J. Am. Chem. Soc. 2011, 133, 1877–1884. [Google Scholar]
- Mates, J.M. Effects of antioxidant enzymes in the molecular control of reactive oxygen species toxicology. Toxicology 2000, 153, 83–104. [Google Scholar] [CrossRef]
- D'Autreaux, B.; Toledano, M.B. ROS as signalling molecules: Mechanisms that generate specificity in ROS homeostasis. Nat. Rev. Mol. Cell Biol. 2007, 8, 813–824. [Google Scholar] [CrossRef]
- Huppert, J.L.; Balasubramanian, S. Prevalence of quadruplexes in the human genome. Nucleic Acids Res. 2005, 33, 2908–2916. [Google Scholar] [CrossRef]
- Todd, A.K.; Johnston, M.; Neidle, S. Highly prevalent putative quadruplex sequence motifs in human DNA. Nucleic Acids Res. 2005, 33, 2901–2907. [Google Scholar]
- Schumann, S.; Hatton, R.A.; Jones, T.S. Organic photovoltaic: Devices based on water-soluble copper phthalocyanine. J. Phys. Chem. C 2011, 115, 4916–4921. [Google Scholar]
- Ryan, J.W.; Anaya-Plaza, E.; Escosura, A.D.; Torres, T.; Palomares, E. Small molecule solar cells based on a series of water-soluble zinc phthalocyanine donors. Chem. Commun. 2012, 48, 6094–6096. [Google Scholar] [CrossRef]
- Liu, Y.; Lin, C.X.; Li, H.Y.; Yan, H. Aptamer-directed self-assembly of protein arrays on a DNA nanostructure. Angew. Chem. Int. Ed. Engl. 2005, 44, 4333–4338. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yaku, H.; Murashima, T.; Miyoshi, D.; Sugimoto, N. Specific Binding of Anionic Porphyrin and Phthalocyanine to the G-Quadruplex with a Variety of in Vitro and in Vivo Applications. Molecules 2012, 17, 10586-10613. https://doi.org/10.3390/molecules170910586
Yaku H, Murashima T, Miyoshi D, Sugimoto N. Specific Binding of Anionic Porphyrin and Phthalocyanine to the G-Quadruplex with a Variety of in Vitro and in Vivo Applications. Molecules. 2012; 17(9):10586-10613. https://doi.org/10.3390/molecules170910586
Chicago/Turabian StyleYaku, Hidenobu, Takashi Murashima, Daisuke Miyoshi, and Naoki Sugimoto. 2012. "Specific Binding of Anionic Porphyrin and Phthalocyanine to the G-Quadruplex with a Variety of in Vitro and in Vivo Applications" Molecules 17, no. 9: 10586-10613. https://doi.org/10.3390/molecules170910586
APA StyleYaku, H., Murashima, T., Miyoshi, D., & Sugimoto, N. (2012). Specific Binding of Anionic Porphyrin and Phthalocyanine to the G-Quadruplex with a Variety of in Vitro and in Vivo Applications. Molecules, 17(9), 10586-10613. https://doi.org/10.3390/molecules170910586