Salvianolic Acid A Protects the Peripheral Nerve Function in Diabetic Rats through Regulation of the AMPK-PGC1α-Sirt3 Axis

Abstract
:1. Introduction

2. Results and Discussion
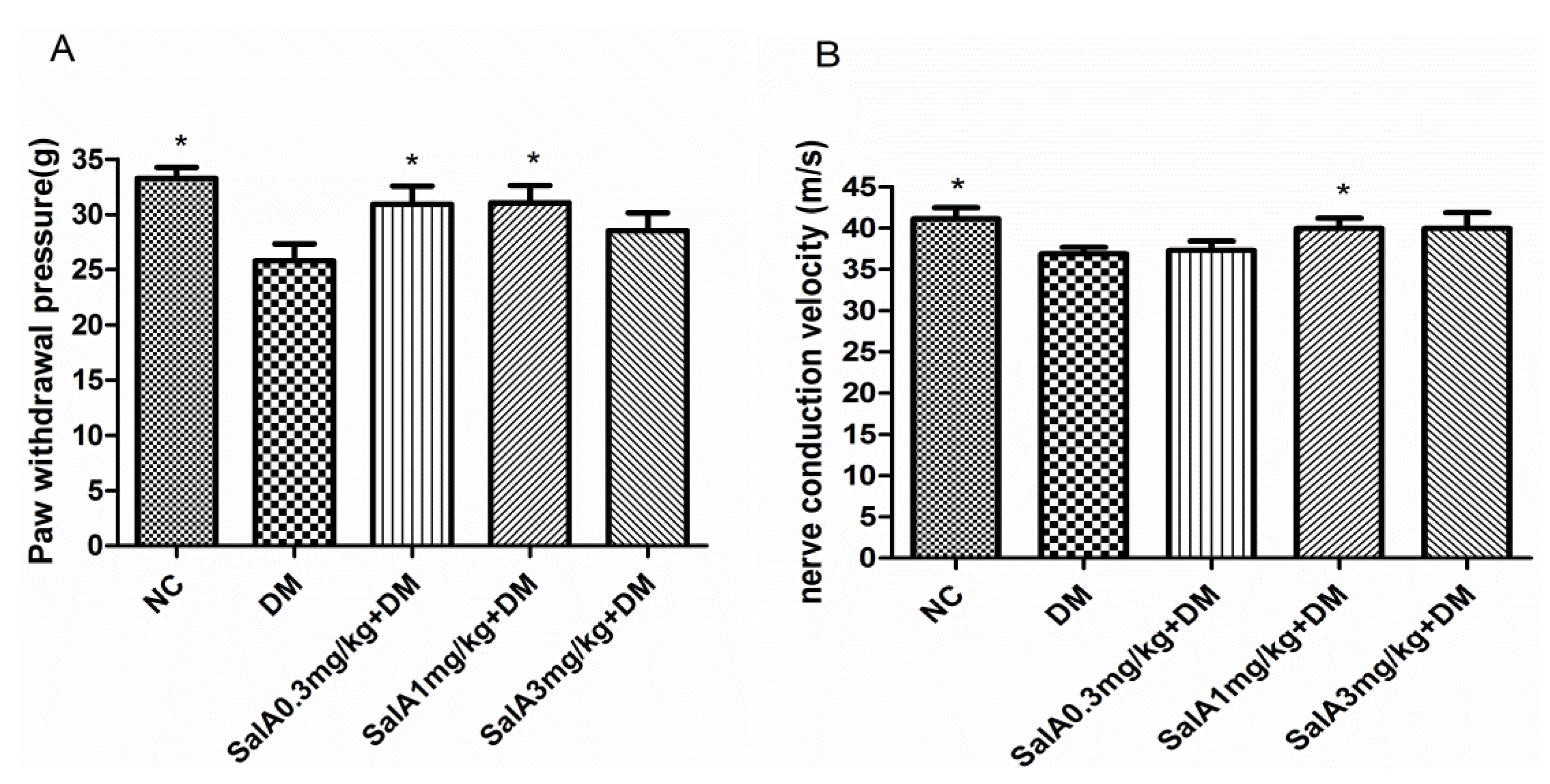
2.1. Effects of SalA on Mechanical Allodynia and Motor Nerve Conduction Velocity (MNCV) in Diabetic Rats

2.2. SalA Attenuated the Ultramicrostructure Change of Sciatic Nerve in Diabetic Rats

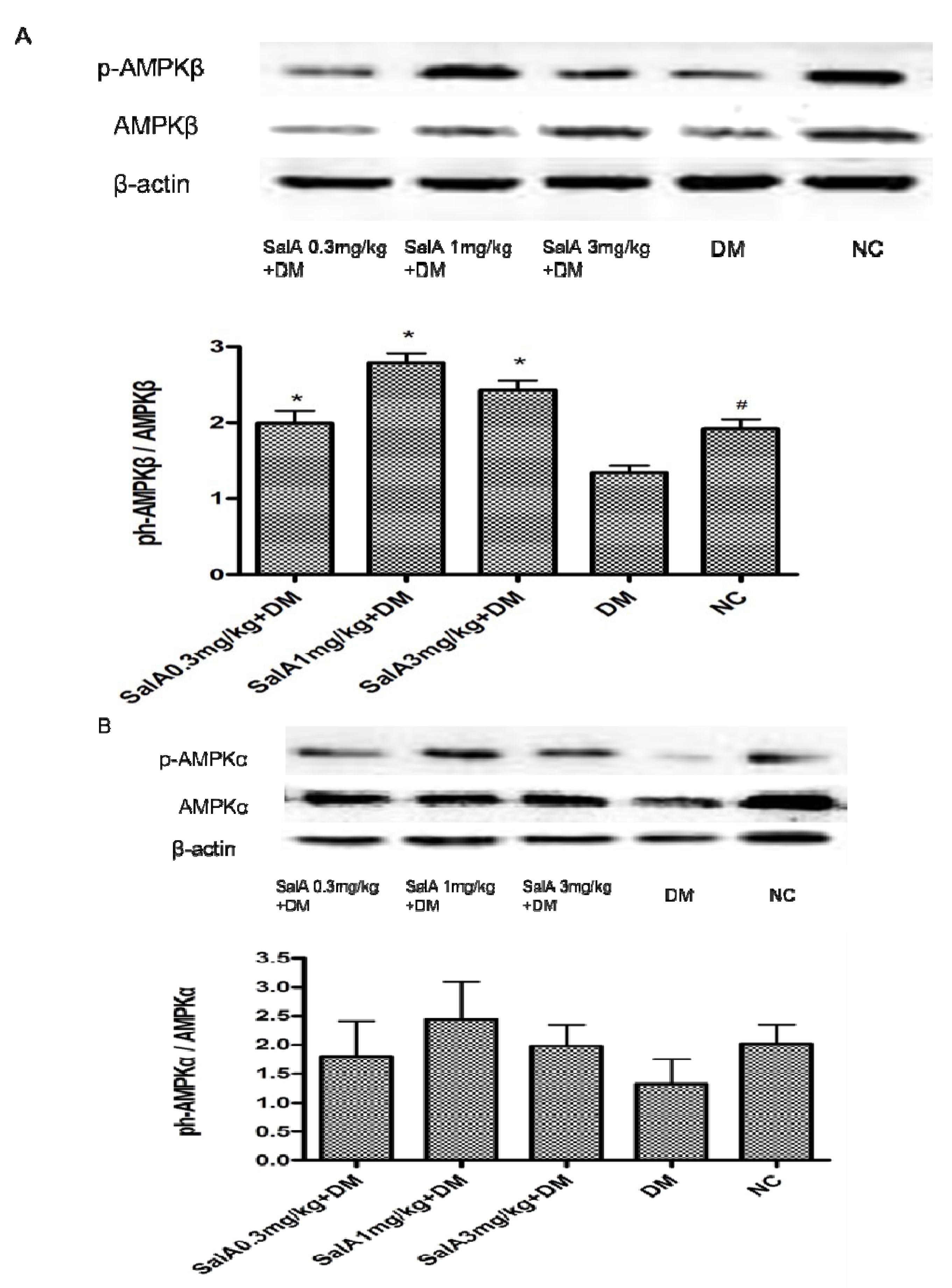
2.3. Effects of Sal A on p-AMPK/AMPKα and p-AMPKβ/AMPKβ in Diabetic Rats

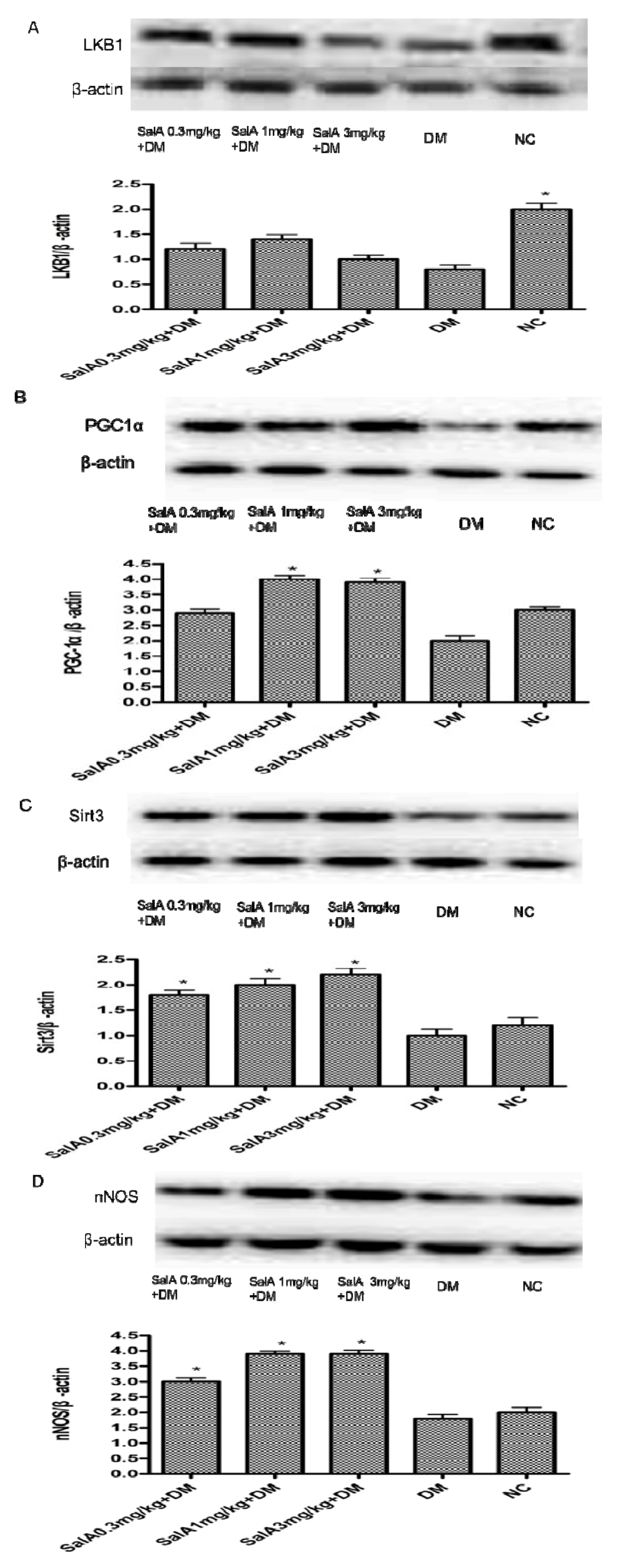
2.4. Effects of SalA on the Expression of LKB, PGC-1α, Sirt3 and nNOS in Diabetic Rat Sciatic Nerve

2.5. Effects of SalA on Body Weight, Blood Glucose, Malondialdehyde (MDA), Superoxide Dismutase (SOD) and Advanced Glycosylation End-products (AGEs)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | BW (g) | BG (mmol/L) | MDA (nmol/L) | SOD (U/mL) | AGEs (ug/mL) |
|---|---|---|---|---|---|
| NC | 511.4 ± 49.0 * | 7.6 ± 3.8 * | 14.0 ± 2.1 * | 148.8 ± 35.3 * | 10.0 ± 0.7 * |
| DM | 350.1 ± 43.1 | 16.2 ± 0.6 | 22.9 ± 8.2 | 98.7 ± 32.1 | 17.2 ± 1.6 |
| DM + SalA0.3 mg/kg | 348.4 ± 24.8 | 16.1 ± 0.4 | 15.7 ± 4.7 * | 136.4 ± 37.7 * | 16.0 ± 0.9 |
| DM + SalA1 mg/kg | 356.8 ± 30.5 | 15.8 ± 0.4 | 12.9 ± 3.8 * | 149.7 ± 33.1 * | 14.6 ± 1.4 * |
| DM + SalA3 mg/kg | 363.3 ± 33.4 | 16.1 ± 0.5 | 14.6 ± 5.4 * | 153.2 ± 52.1 * | 15.2 ± 1.0 |
3. Experimental
3.1. Animals
3.2. Induction of Experimental Diabetes
3.3. Treatment
3.4. Measurement of Paw Withdrawal Mechanical Threshold (PWMT)
3.5. Evaluation of Motor Nerve Conduction Velocity (MNCV)

3.6. Observation of Ultramicrostructure of Sciatic Nerve
3.7. Measurement of Body Weight, Blood Glucose, MDA, SOD and AGEs.
3.8. Western Blot
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Fernyhough, P.; Calcutt, N.A. Abnormal calcium homeostasis in peripheral neuropathies. Cell Calcium. 2010, 47, 130–139. [Google Scholar] [CrossRef]
- Tesfaye, S.; Boulton, A.J.; Dyck, P.J.; Freeman, R.; Horowitz, M.; Kempler, P.; Lauria, G.; Malik, R.A.; Spallone, V.; Vinik, A.; et al. Diabetic neuropathies: Update on definitions, diagnostic criteria, estimation of severity, and treatments. Diabetes Care 2010, 33, 2285–2293. [Google Scholar] [CrossRef] [Green Version]
- Brownlee, M. Biochemistry and molecular cell biology of diabetic complications. Nature 2001, 414, 813–820. [Google Scholar] [CrossRef]
- Huang, Q.; Wu, Y.T.; Tan, H.L.; Ong, C.N.; Shen, H.M. A novel function of poly(ADP-ribose) polymerase-1 in modulation of autophagy and necrosis under oxidative stress. Cell Death Differ. 2009, 16, 264–277. [Google Scholar] [CrossRef]
- Ronnett, G.V.; Ramamurthy, S.; Kleman, A.M.; Landree, L.E.; Aja, S. AMPK in the brain: Its roles in energy balance and neuroprotection. J. Neurochem. 2009, 109, 17–23. [Google Scholar] [CrossRef]
- Shin, S.M.; Cho, I.J.; Kim, S.G. Resveratrol protects mitochondria against oxidative stress through AMP-activated protein kinase-mediated glycogen synthasekinase-3β inhibition downstream of poly(ADP-ribose)polymerase-LKB1 pathway. Mol. Pharmacol. 2009, 76, 884–895. [Google Scholar] [CrossRef]
- Melemedjian, O.K.; Asiedu, M.N.; Tillu, D.V.; Sanoja, R.; Yan, J.; Lark, A.; Khoutorsky, A.; Johnson, J.; Peebles, K.A.; Lepow, T.; et al. Targeting adenosine monophosphate-activated protein kinase (AMPK) in preclinical models reveals a potential mechanism for the treatment of neuropathic pain. Mol. Pain 2011, 7. [Google Scholar]
- Chowdhury, S.R.; Dobrowsky, R.T.; Fernyhough, P. Nutrient excess and altered mitochondrial proteome and function contribute to neurodegeneration in diabetes. Mitochondrion 2011, 11, 845–854. [Google Scholar] [CrossRef]
- Hirschey, M.D.; Shimazu, T.; Goetzman, E.; Jing, E.; Schwer, B.; Lombard, D.B.; Grueter, C.A.; Harris, C.; Biddinger, S.; Ilkayeva, O.R.; et al. SIRT3 regulates mitochondrial fatty-acid oxidation by reversible enzyme deacetylation. Nature 2010, 464, 121–125. [Google Scholar]
- Smith, B.C.; Hallows, W.C.; Denu, J.M. A continuous microplate assay for sirtuins and nicotinamide-producing enzymes. Anal. Biochem. 2009, 394, 101–109. [Google Scholar]
- Cheng, T.O. Cardiovascular effects of Danshen. Int. J. Cardiol. 2007, 121, 9–22. [Google Scholar] [CrossRef]
- Wang, S.B.; Yang, X.Y.; Tian, S.; Yang, H.G.; Du, G.H. Effect of salvianolic acid A on vascular reactivity of streptozotocin-induced diabetic rats. Life Sci. 2009, 85, 499–504. [Google Scholar]
- Yang, X.Y.; Sun, L.; Xu, P.; Gong, L.L.; Qiang, G.F.; Zhang, L.; Du, G.H. Effects of salvianolic scid A on plantar microcirculation and peripheral nerve function in diabetic rats. Eur. J. Pharmacol. 2011, 665, 40–46. [Google Scholar] [CrossRef]
- Hardie, D.G.; Sakamoto, K. AMPK: A key sensor of fuel and energy status in skeletal muscle. Physiology (Bethesda) 2006, 21, 48–60. [Google Scholar] [CrossRef]
- Emerling, B.M.; Weinberg, F.; Snyder, C.; Burgess, Z.; Mutlu, G.M.; Viollet, B.; Budinger, G.R.S.; Chandel, N.S. Hypoxic activation of AMPK is dependent on mitochondrial ROS but independent of an increase in AMP/ATP ratio. Free Radic. Biol. Med. 2009, 46, 1386–1391. [Google Scholar] [CrossRef]
- Fogarty, S.; Hardie, D.G. Development of protein kinase activators: AMPK as a target in metabolic disorders and cancer. Biochim. Biophys. Acta 2010, 1804, 581–591. [Google Scholar] [CrossRef]
- Hao, W.S.; Chang, C.P.B.; Tsao, C.C.; Xu, J. Oligomycin-induced bioenergetic adaptation in cancer cells with heterogeneous bioenergetic organization. J. Biol. Chem. 2010, 285, 12647–12654. [Google Scholar]
- Polekhina, G.; Gupta, A.; van Denderen, B.J.; Feil, S.C.; Kemp, B.E.; Stapleton, D.; Parker, M.W. Structural basis for glycogen recognition by AMP-activated protein kinase. Structure 2005, 13, 1453–1462. [Google Scholar] [CrossRef]
- Ho, J.H.C.; Hong, C.Y. Salvianolic acids: Small compounds with multiple mechanisms for cardiovascular protection. J. Biomed. Sci. 2011, 18, 1–5. [Google Scholar] [CrossRef]
- Sperl, B.; Seifert, M.H.J.; Berg, T. Natural product inhibitors of protein-protein interactions mediated by Src-family SH2 domains. Bioorg. Med. Chem. Lett. 2009, 19, 3305–3309. [Google Scholar] [CrossRef]
- Shackelford, D.B.; Shaw, R.J. The LKB1-AMPK pathway: Metabolism and growth control in tumor suppression. Nat. Rev. Cancer 2009, 9, 563–575. [Google Scholar] [CrossRef]
- Handschin, J.S.; Pierre, C.S; Spiegelman, B.M. AMP-activated protein kinase (AMPK) action in skeletal muscle via direct phosphorylation of PGC-1alpha. Proc. Natl. Acad. Sci. USA 2007, 104, 12017–12022. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, M.D.; Tauler, P.; Sureda, A.; Tur, J.A.; Pons, A. Antioxidant regulatory mechanisms in neutrophils and lymphocytes after intense exercise. J. Sports Sci. 2009, 27, 49–58. [Google Scholar] [CrossRef]
- Kim, D.; Nguyen, M.D.; Dobbin, M.M.; Fischer, A.; Sananbenesi, F.; Rodgers, J.T.; Delalle, I.; Baur, J.A.; Sui, G.; Armour, S.M.; et al. SIRT1 deacetylase protects against neurodegeneration in models for Alzheimer’s disease and amyotrophic lateral sclerosis. EMBO J. 2007, 26, 3169–3179. [Google Scholar] [CrossRef]
- Li, J.; Hu, X.Y.; Selvakumar, P.; Russell, R.R., 3rd.; Cushman, S.W.; Holman, G.D.; Young, L.H. Role of the nitric oxide pathway in AMPK-mediated glucose uptake and GLUT4 translocation in heart muscle. Am. J. Physiol. Endocrinol. Metab. 2004, 287, E834–E841. [Google Scholar] [CrossRef]
- Chen, Z.P.; McConell, G.K.; Michell, B.J.; Snow, R.J.; Canny, B.J.; Kemp, B.E. AMPK signaling in contracting human skeletal muscle: Acetyl-CoA carboxylase and NO synthase phosphorylation. Am. J. Physiol. Endocrinol. Metab. 2000, 279, E1202–E1206. [Google Scholar]
- Gangula, P.R.R.; Mukhopadhyay, S.; Pasricha, P.J.; Ravella, K. Sepiapterin reverses the changes in gastric nNOS dimerization and function in diabetic gastroparesis. Neurogastroenterol. Motil. 2010, 22, e1325–e1352. [Google Scholar] [CrossRef]
- Vareniuk, I.; Pacher, P.; Pavlov, I.A.; Drel, V.R.; Obrosova, I.G. Peripheral neuropathy in mice with neuronal nitric oxide synthase gene deficiency. Int. J. Mol. Med. 2009, 23, 571–580. [Google Scholar]
- Vincent, A.M.; Russell, J.W.; Low, P.; Feldman, E.L. Oxidative stress in the pathogenesis of diabetic neuropathy. Endocr. Rev. 2004, 25, 612–628. [Google Scholar] [CrossRef]
- Nishikawa, T.; Edelstein, D.; Du, X.L.; Yamagishi, S.; Matsumura, T.; Kaneda, Y.; Yorek, A.; Beebe, D.; Oates, P.J.; Hammes, H.P.; et al. Normalizing mitochondrial superoxide production blocks three pathways of hyperglycaemic damage. Nature 2000, 404, 787–790. [Google Scholar]
- Yang, X.Y.; Qiang, G.F.; Zhang, L.; Zhu, X.M.; Wang, S.B.; Sun, L.; Yang, H.G.; Du, G.H. Salvianolic acid A protects against vascular endothelial dysfunction in high-fat diet fed and streptozotocin-induced diabetic rats. J. Asian Nat. Prod. Res. 2011, 13, 884–894. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yu, X.; Zhang, L.; Yang, X.; Huang, H.; Huang, Z.; Shi, L.; Zhang, H.; Du, G. Salvianolic Acid A Protects the Peripheral Nerve Function in Diabetic Rats through Regulation of the AMPK-PGC1α-Sirt3 Axis. Molecules 2012, 17, 11216-11228. https://doi.org/10.3390/molecules170911216
Yu X, Zhang L, Yang X, Huang H, Huang Z, Shi L, Zhang H, Du G. Salvianolic Acid A Protects the Peripheral Nerve Function in Diabetic Rats through Regulation of the AMPK-PGC1α-Sirt3 Axis. Molecules. 2012; 17(9):11216-11228. https://doi.org/10.3390/molecules170911216
Chicago/Turabian StyleYu, Xiaoyan, Li Zhang, Xiuying Yang, Huakang Huang, Zhonglin Huang, Lili Shi, Hengai Zhang, and Guanhua Du. 2012. "Salvianolic Acid A Protects the Peripheral Nerve Function in Diabetic Rats through Regulation of the AMPK-PGC1α-Sirt3 Axis" Molecules 17, no. 9: 11216-11228. https://doi.org/10.3390/molecules170911216