Chamaejasmine Arrests Cell Cycle, Induces Apoptosis and Inhibits Nuclear NF-κB Translocation in the Human Breast Cancer Cell Line MDA-MB-231

Abstract
:1. Introduction

2. Results
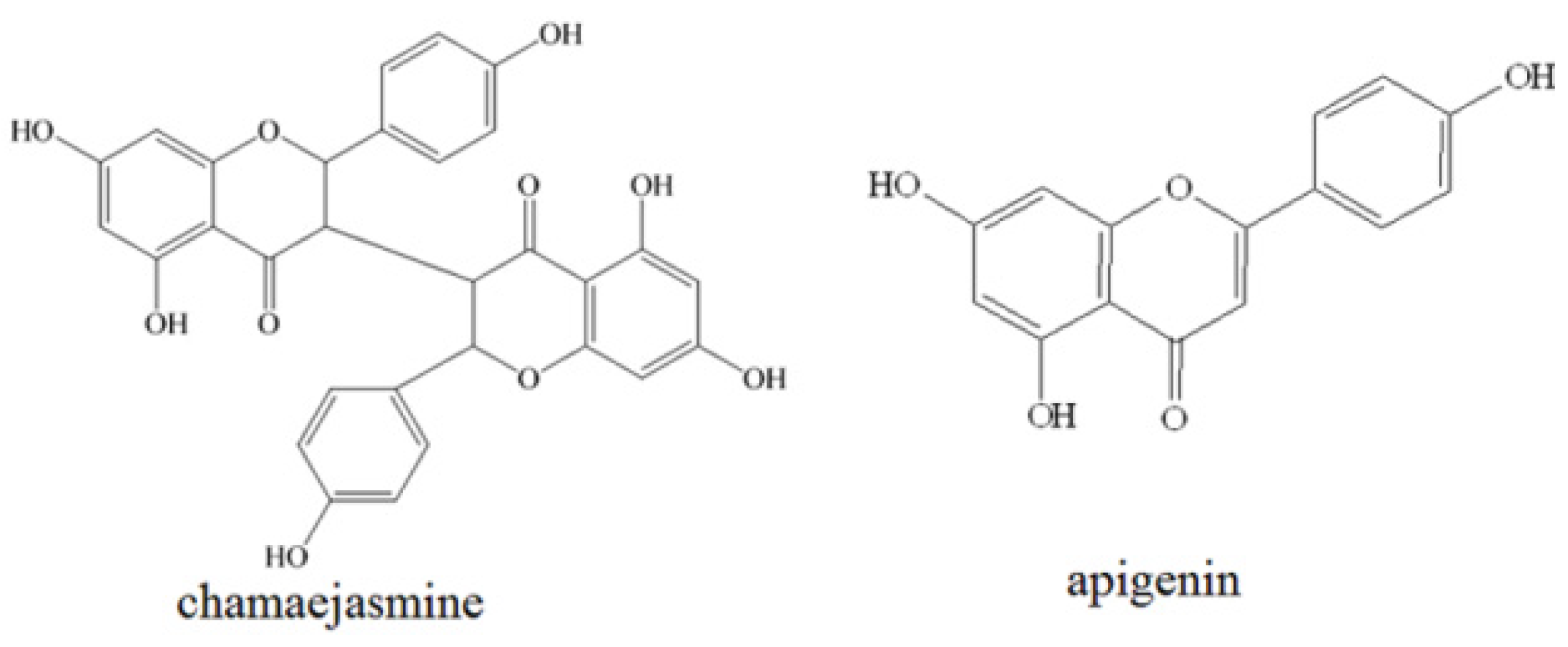
2.1. Heading Cytotoxicity Assays

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell lines | IC50 (μM) | |
|---|---|---|
| Chamaejasmine | Apigenin | |
| HCC1937 | 13.44 * | 35.97 |
| MDA-MB-453 | 5.66 * | 31.12 |
| MDA-MB-231 | 4.72 * | 21.77 |
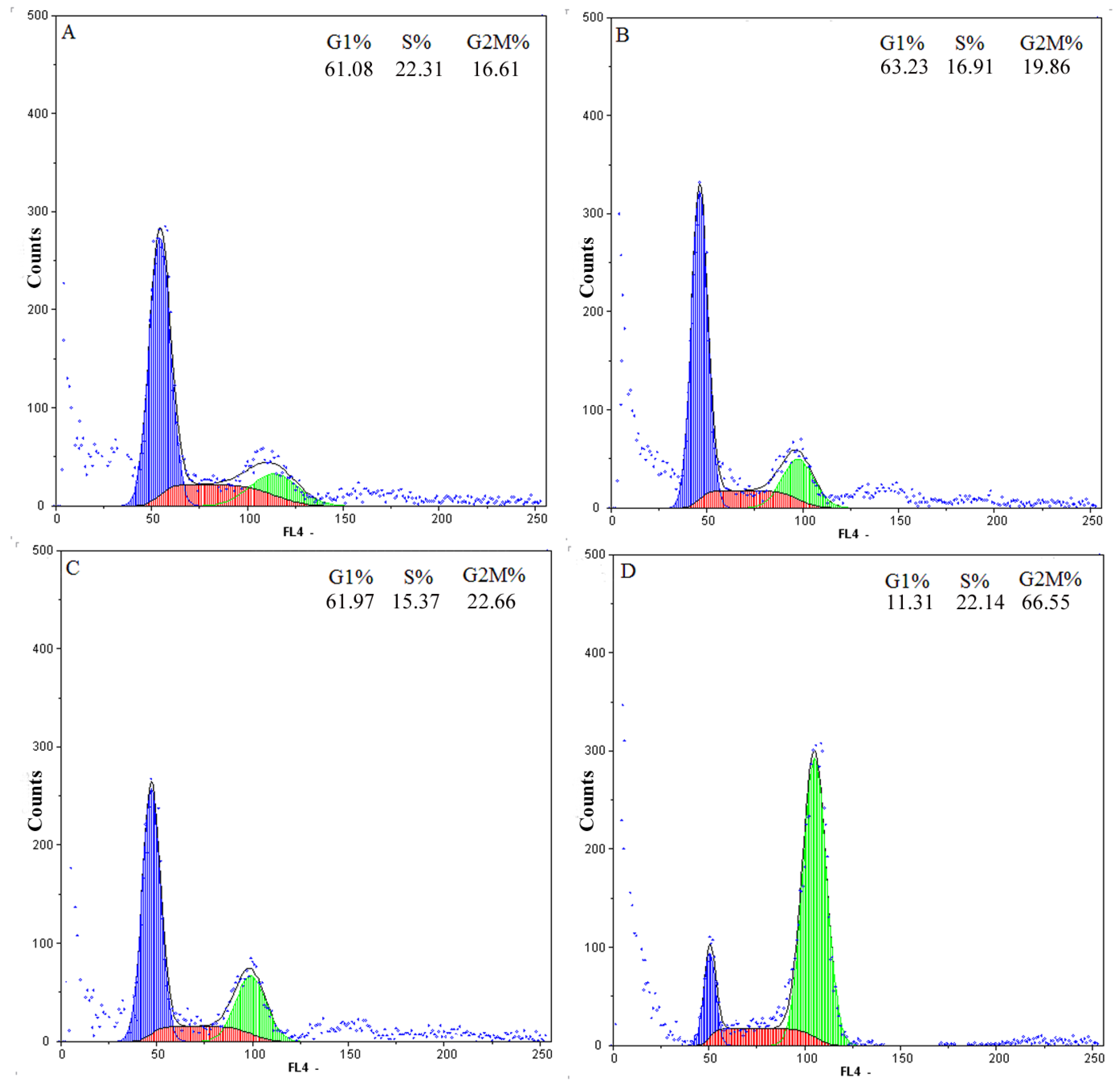
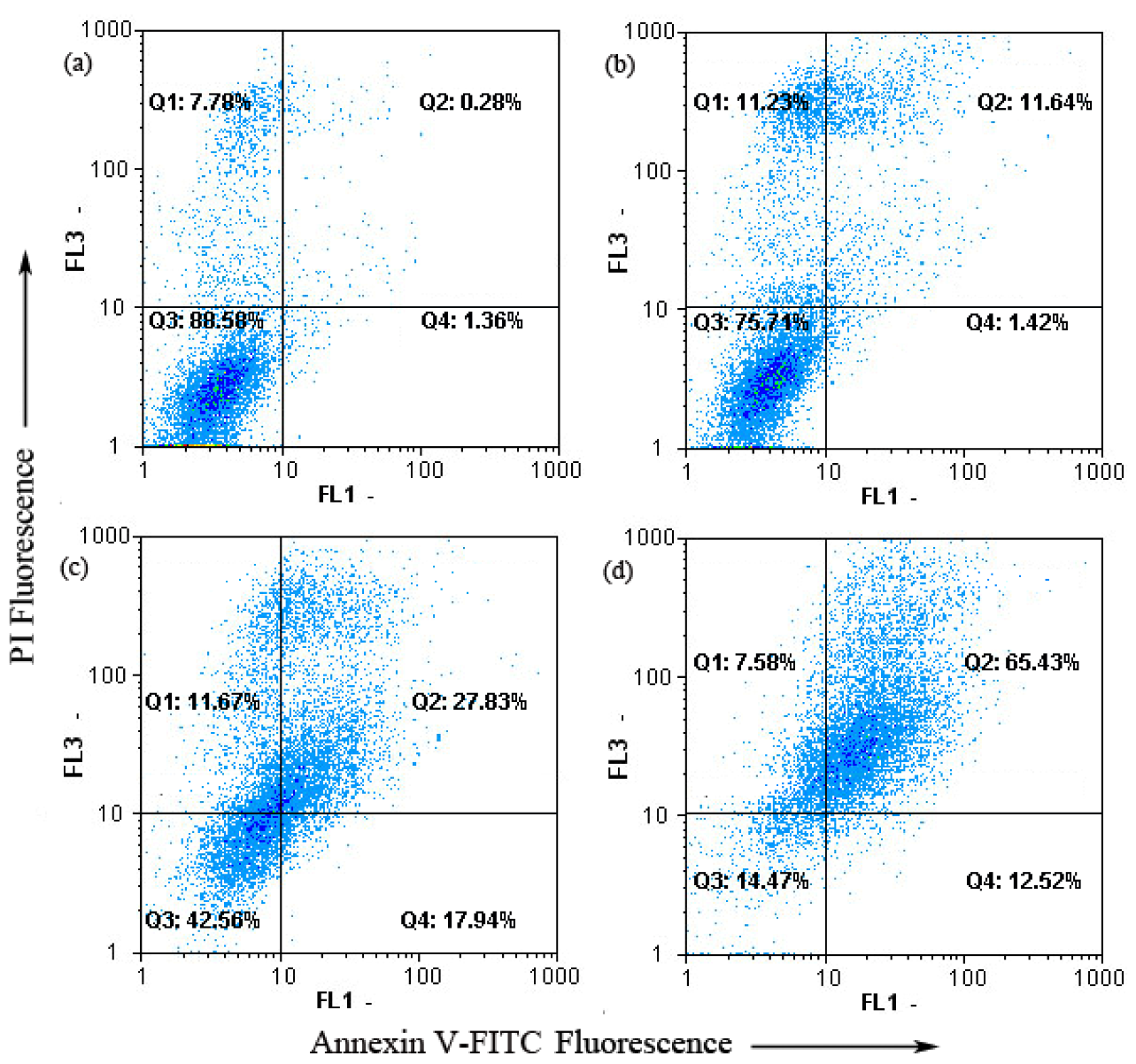
2.2. G2-M Phase Cell Cycle Arrest and Apoptosis by Chamaejasmine in MDA-MB-231 Cells

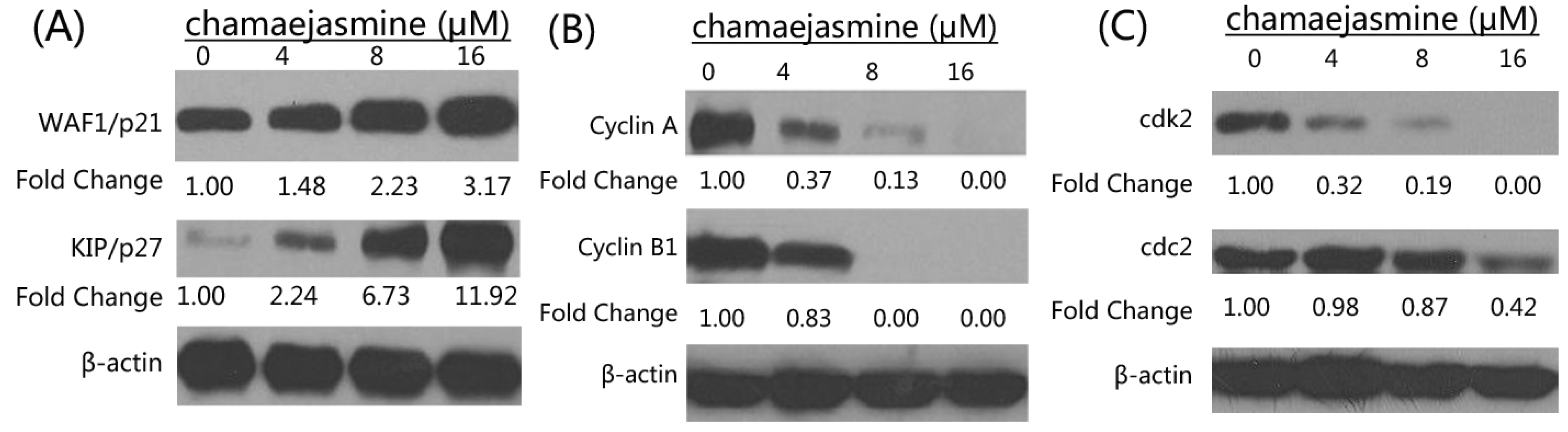
2.3. Inhibition of Cyclins, Cdk2, cdc2 and Induction of WAF1/p21 and KIP1/p27 by Chamaejasmine in MDA-MB-231 Cells


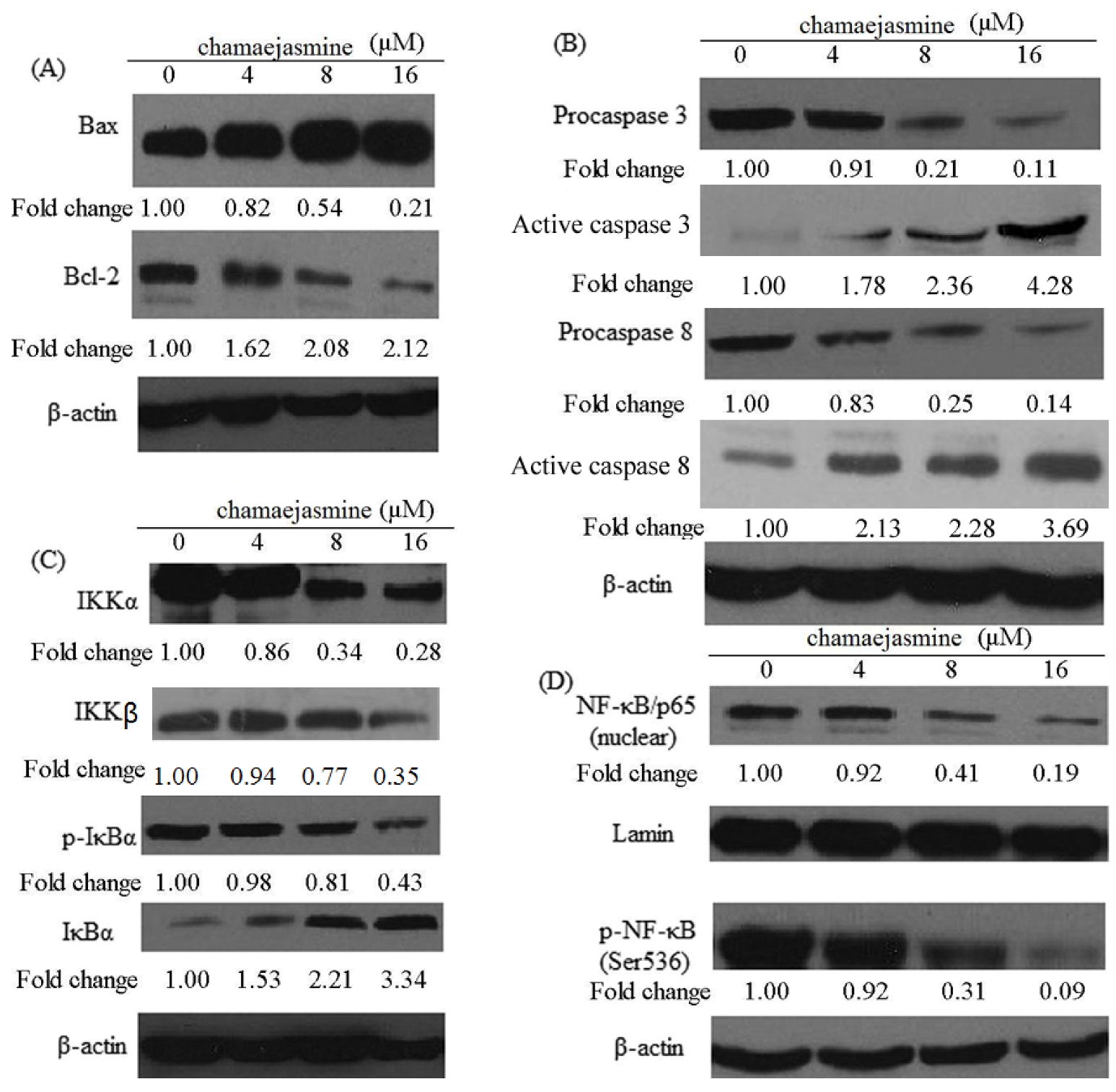
2.4. Induction of Bax and Inhibition of Bcl-2 and Procaspases in MDA-MB-231 Cells

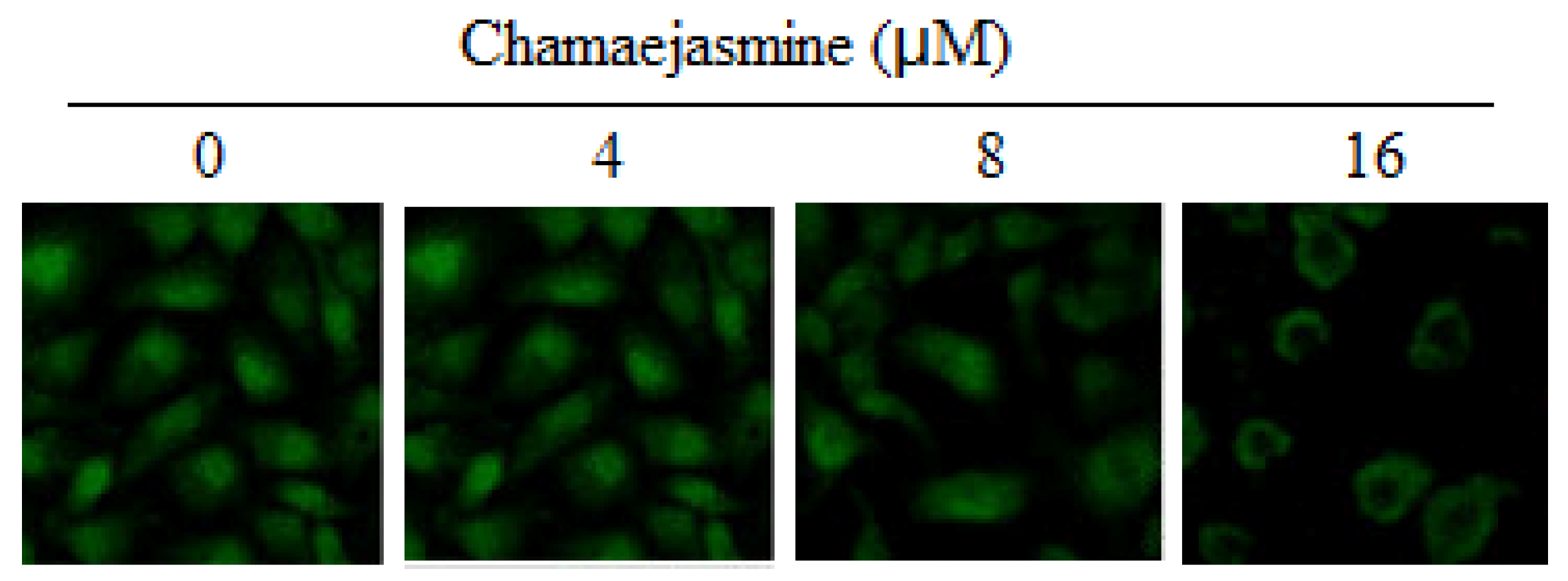
2.5. Inhibition of NF-κB Pathway by chamaejasmine in MDA-MB-231 Cells

3. Discussion
4. Experimental
4.1. Materials
4.2. Cell Culture and Chemicals
4.3. Cytotoxicity Assay
4.4. Flow Cytometric Analysis of Cell Cycle and Apoptosis
4.5. Preparation of Cytosolic and Nuclear Lysates
4.6. Western Blotting Assay
4.7. Immunofluorescence Assay
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
- Sample Availability: Samples of the compounds are available from the authors.
References
- Mohinta, S.; Mohinta, H.; Chaurasia, P.; Watabe, K. Wnt pathway and breast cancer. Front. Biosci. 2007, 12, 4020–4033. [Google Scholar] [CrossRef]
- Desantis, C.; Siegel, R.; Bandi, P.; Jemal, A. Breast cancer statistics, 2011. CA Cancer J. Clin. 2011, 61, 409–418. [Google Scholar]
- Mavrogenis, A.F.; Pala, E.; Romantini, M.; Guerra, G.; Romagnoli, C.; Maccauro, G.; Ruggieri, P. Side effects of radiation in musculoskeletal oncology: Clinical evaluation of radiationinduced fractures. Int. J. Immunopathol. Pharmacol. 2011, 24, 29–37. [Google Scholar]
- Mavrogenis, A.F.; Papagelopoulos, P.J.; Romantini, M.; Angelini, A.; Ruggieri, P. Side effects of chemotherapy in musculoskeletal oncology. J. Long Term Eff. Med. Implants 2010, 20, 1–12. [Google Scholar] [CrossRef]
- Koo, N.K.; Kim, Y.C. A Case of Paclitaxel-induced Maculopathy Treated with Methazolamide. Korean J. Ophthalmol. 2012, 26, 394–397. [Google Scholar] [CrossRef]
- Yim, D.; Singh, R.P.; Agarwal, C.; Lee, S.; Chi, H.; Agarwal, R. A Novel Anticancer Agent, Decursin, Induces G1 Arrest and Apoptosis in Human Prostate Carcinoma Cells. Cancer Res. 2005, 65, 1035–1044. [Google Scholar]
- Tomasini, R.; Mak, T.W.; Melino, G. The impact of p53 and p73 on aneuploidy and cancer. Trends Cell Biol. 2008, 18, 244–252. [Google Scholar] [CrossRef]
- Vinatier, D.; Dufour, P.; Subtil, D. Apoptosis: A programmed cell death involved in ovarian and uterine physiology. Eur. J. Obstet. Gyn. R. B. 1996, 67, 85–102. [Google Scholar] [CrossRef]
- Das, G.C.; Holiday, D.; Gallardo, R.; Haas, C. Taxol-induced cell cycle arrest and apoptosis: Dose-response relationship in lung cancer cells of different wild-type p53 status and under isogenic condition. Cancer Lett. 2001, 165, 147–153. [Google Scholar] [CrossRef]
- Debatin, K.-M. Activation of apoptosis pathways by anticancer treatment. Toxicol. Lett. 2000, 112, 41–48. [Google Scholar] [CrossRef]
- Da Silva, C.P.; de Oliveira, C.R.; de Lima, M.C.P. Apoptosis as a mechanism of cell death inducedby different chemotherapeutic drugs in human leukemic T-lymphocytes. Biochem. Pharmacol. 1996, 51, 1331–1340. [Google Scholar]
- Sarraf, C.E.; Bowen, I.D. Proportions of Mitotic and Apoptotic Cells In A Range of Untreated Experimental Tumours. Cell Prolif. 1988, 21, 45–49. [Google Scholar] [CrossRef]
- Carson, D.A.; Ribeiro, J.M. Apoptosis and disease. Lancet 1993, 341, 1251–1254. [Google Scholar]
- Newman, D.J.; Cragg, G.M.; Snader, K.M. Natural Products as Sources of New Drugs over the Period 1981–2002. J. Nat. Prod. 2003, 66, 1022–1037. [Google Scholar] [CrossRef]
- Cragg, G.M.; Newman, D.J. Plants as a source of anti-cancer agents. J. Ethnopharmacol. 2005, 100, 72–79. [Google Scholar] [CrossRef]
- Shi, Z.C. The Poisonous Plants in Chinese Pasture; China Agricultural Press: Beijing, China, 1997. [Google Scholar]
- Zhao, S.H.; Wan, S.Q. Progress of investigations and application of insecticide plants. Guangdong Agric. Sci. 1997, 1, 26–28. [Google Scholar]
- Feng, W.; Tetsuro, I.; Mitsuzi, Y. The antitumor activities of gnidimacrin isolated from Stellera chamaejasme L. Zhonghua Zhong Liu Za Zhi 1996, 8, 101–104. [Google Scholar]
- Jin, C.; Michetich, R.G.; Daneshtalab, M. Flavonoids from Stellera chamaejasme. Phytochemistry 1999, 50, 505–508. [Google Scholar] [CrossRef]
- Yang, G.; Chen, D. Biflavanones, Flavonoids, and Coumarins from the Roots of Stellera chamaejasme and Their Antiviral Effect on Hepatitis B Virus. Chem. Biodivers. 2008, 5, 1419–1424. [Google Scholar] [CrossRef]
- Malumbres, M.; Barbacid, M. Cell cycle, CDKs and cancer: A changing paradigm. Nat. Rev. Cancer 2009, 9, 153–166. [Google Scholar] [CrossRef]
- Herman, J.G.; Merlo, A.; Mao, L.; Lapidus, R.G.; Issa, J.P.; Davidson, N.E.; Sidransky, D.; Baylin, S.B. Inactivation of the CDKN2/p16/MTS1 Gene Is Frequently Associated with Aberrant DNA Methylation in All Common Human Cancers. Cancer Res. 1995, 55, 4525–4530. [Google Scholar]
- Wang, Y.; Zhao, Y.; Liu, Y.; Tian, L.; Jin, D. Chamaejasmine Inactivates Akt To Trigger Apoptosis in Human HEp-2 Larynx Carcinoma Cells. Molecules 2011, 16, 8152–8164. [Google Scholar] [CrossRef]
- Fang, W.; Liu, S.; Nie, Y. Anticancer Activity of Chamaejasmine: Effect on Tubulin Protein. Molecules 2011, 16, 6243–6254. [Google Scholar] [CrossRef]
- Gu, S.; He, J. Daphnoretin Induces Cell Cycle Arrest and Apoptosis in Human Osteosarcoma (HOS) Cells. Molecules 2012, 17, 598–612. [Google Scholar] [CrossRef]
- Hartwell, L.H.; Kastan, M.B. Cell cycle control and cancer. Science 1994, 266, 1821–1828. [Google Scholar]
- Vermeulen, K.; van Bockstaele, D.R.; Berneman, Z.N. The cell cycle: A review of regulation, deregulation and therapeutic targets in cancer. Cell Prolif. 2003, 36, 131–149. [Google Scholar] [CrossRef]
- Jeong, H.-W.; Han, D.C.; Son, K.-H.; Han, M.Y.; Lim, J.S.; Ha, J.H.; Lee, C.W.; Kim, H.M.; Kim, H.C.; Kwon, B.M. Antitumor effect of the cinnamaldehyde derivative CB403 through the arrest of cell cycle progression in the G2/M phase. Biochem. Pharmacol. 2003, 65, 1343–1350. [Google Scholar] [CrossRef]
- Buolamwini, J.K. Cell Cycle Molecular Targets in Novel Anticancer Drug Discovery. Curr. Pharm. Des. 2000, 14, 379–392. [Google Scholar] [CrossRef]
- McDonald, E.R., III; El-Deiry, W.S. Cell cycle control as a basis for cancer drug development. Int. J. Oncol. 2000, 16, 871–957. [Google Scholar]
- Devault, A.C.J.; Fesquet, D.; Labbé, J.C.; Lorca, T.; Picard, A.; Strausfeld, U.; Dorée, M. Concerted roles of cyclin A, cdc25+ mitotic inducer, and type 2A phosphatase in activating the cyclin B/cdc2 protein kinase at the G2/M phase transition. Cold Spring Harb. Symp. Quant. Biol. 1991, 56, 503–513. [Google Scholar] [CrossRef]
- Van den Heuvel, S.; Harlow, E. Distinct roles for cyclin-dependent kinases in cell cycle control. Science 1993, 262, 2050–2054. [Google Scholar]
- Nurse, P. Universal control mechanism regulating onset of M-phase. Nature 1990, 344, 503–508. [Google Scholar] [CrossRef]
- Piao, W.; Yoo, J.; Lee, K.D.; Hwang, H.J.; Kim, J.H. Induction of G2/M phase arrest and apoptosis by a new synthetic anti-cancer agent, DW2282, in promyelocytic leukemia (HL-60) cells. Biochem. Pharmacol. 2001, 62, 1439–1447. [Google Scholar] [CrossRef]
- Hockenbery, D.M.; Oltvai, Z.N.; Yin, X.-M.; Milliman, C.L.; Korsmeyer, S.J. Bcl-2 functions in an antioxidant pathway to prevent apoptosis. Cell 1993, 75, 241–251. [Google Scholar] [CrossRef]
- Reed, J.C. Regulation of apoptosis by bcl-2 family proteins and its role in cancer and chemoresistance. Curr. Opin. Oncol. 1995, 7, 541–546. [Google Scholar] [CrossRef]
- Revelos, K.; Petraki, C.; Gregorakis, A.; Scorilas, A.; Papanastasiou, P.; Koutsilieris, M. Immunohistochemical Expression of Bcl2 is an Independent Predictor of Time-to-biochemical Failure in Patients with Clinically Localized Prostate Cancer Following Radical Prostatectomy. Anticancer Res. 2005, 25, 3123–3133. [Google Scholar]
- Nicholson, D.W. Caspase structure, proteolytic substrates, and function during apoptotic cell death. Cell Death Differ. 1999, 6, 1028–1042. [Google Scholar]
- Grutter, M.G. Caspases: Key players in programmed cell death. Curr. Opin. Struct. Biol. 2000, 10, 649–655. [Google Scholar]
- Zimmermann, K.C.; Bonzon, C.; Green, D.R. The machinery of programmed cell death. Pharmacol. Ther. 2001, 92, 57–70. [Google Scholar]
- Islam, S.; Islam, N.; Kermode, T.; Johnstone, B.; Mukhtar, H.; Moskowitz, R.W.; Goldberg, V.M.; Malemud, C.J.; Haqqi, T.M. Involvement of Caspase-3 in Epigallocatechin-3-gallate-Mediated Apoptosis of Human Chondrosarcoma Cells. Biochem. Biophys. Res. Commun. 2000, 270, 793–797. [Google Scholar]
- Hayakawa, S.; Saeki, K.; Sazuka, M.; Suzuki, Y; Shoji, Y.; Ohta, T.; Kaji, K.; Yuo, A.; Isemura, M. Apoptosis Induction by Epigallocatechin Gallate Involves Its Binding to Fas. Biochem. Biophys. Res. Commun. 2001, 285, 1102–1106. [Google Scholar]
- Hsu, H.-F.; Houng, J.-Y.; Kuo, C.-F.; Tsao, N.; Wu, Y.-C. Glossogin, a novel phenylpropanoid from Glossogyne tenuifolia, induced apoptosis in A549 lung cancer cells. Food Chem. Toxicol. 2008, 46, 3785–3791. [Google Scholar]
- Lee, J.; Im, Y.-H.; Jung, H.H.; Kim, J.H.; Park, J.O.; Kim, K.; Kim, W.S.; Ahn, J.S.; Jung, C.W.; Park, Y.S.; et al. Curcumin inhibits interferon-α induced NF-κB and COX-2 in human A549 non-small cell lung cancer cells. Biochem. Biophys. Res. Commum. 2005, 334, 313–318. [Google Scholar]
- Gupta, S.; Hastak, K.; Afaq, F.; Ahmad, N.; Mukhtar, H. Essential role of caspases in epigallocatechin-3-gallate-mediated inhibition of nuclear factor kappaB and induction of apoptosis. Oncogene 2004, 23, 2507–2522. [Google Scholar]
- Thati, B.; Noble, A.; Creaven, B.S.; Walsh, M.; McCann, M.; Devereux, M.; Kavanagh, K.; Egan, D.A. Role of cell cycle events and apoptosis in mediating the anti-cancer activity of a silver(I) complex of 4-hydroxy-3-nitro-coumarin-bis(phenanthroline) in human malignant cancer cells. Eur. J. Pharmacol. 2009, 602, 203–214. [Google Scholar] [CrossRef]
- Cude, K.; Wang, Y.; Choi, H.J.; Hsuan, S.L.; Zhang, H.; Wang, C.Y.; Xia, Z. Regulation of the G2-M cell cycle progression by the ERK5-NFjB signaling pathway. J. Cell Biol. 2007, 177, 253–264. [Google Scholar] [CrossRef]
- Curry, C.L.; Reed, L.L.; Broude, E.; Golde, T.E.; Miele, L.; Foreman, K.E. Notch inhibition in Kaposi’s sarcoma tumor cells leads to mitotic catastrophe through nuclear factor-jB signaling. Mol. Cancer Ther. 2007, 6, 1983–1992. [Google Scholar] [CrossRef]
- Ryan, K.M.; Ernst, M.K.; Rice, N.R.; Vousden, K.H. Role of NF-kB in p53-mediated programmed cell death. Nature 2000, 404, 892–897. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, T.; Yu, H.; Dong, G.; Cai, L.; Bai, Y. Chamaejasmine Arrests Cell Cycle, Induces Apoptosis and Inhibits Nuclear NF-κB Translocation in the Human Breast Cancer Cell Line MDA-MB-231. Molecules 2013, 18, 845-858. https://doi.org/10.3390/molecules18010845
Zhang T, Yu H, Dong G, Cai L, Bai Y. Chamaejasmine Arrests Cell Cycle, Induces Apoptosis and Inhibits Nuclear NF-κB Translocation in the Human Breast Cancer Cell Line MDA-MB-231. Molecules. 2013; 18(1):845-858. https://doi.org/10.3390/molecules18010845
Chicago/Turabian StyleZhang, Tingting, Hongyang Yu, Guanglu Dong, Li Cai, and Yuxian Bai. 2013. "Chamaejasmine Arrests Cell Cycle, Induces Apoptosis and Inhibits Nuclear NF-κB Translocation in the Human Breast Cancer Cell Line MDA-MB-231" Molecules 18, no. 1: 845-858. https://doi.org/10.3390/molecules18010845
APA StyleZhang, T., Yu, H., Dong, G., Cai, L., & Bai, Y. (2013). Chamaejasmine Arrests Cell Cycle, Induces Apoptosis and Inhibits Nuclear NF-κB Translocation in the Human Breast Cancer Cell Line MDA-MB-231. Molecules, 18(1), 845-858. https://doi.org/10.3390/molecules18010845