Metabolomics and Fetal-Neonatal Nutrition: Between “Not Enough” and “Too Much”


Abstract
:1. Introduction
2. How Big Is the Problem?
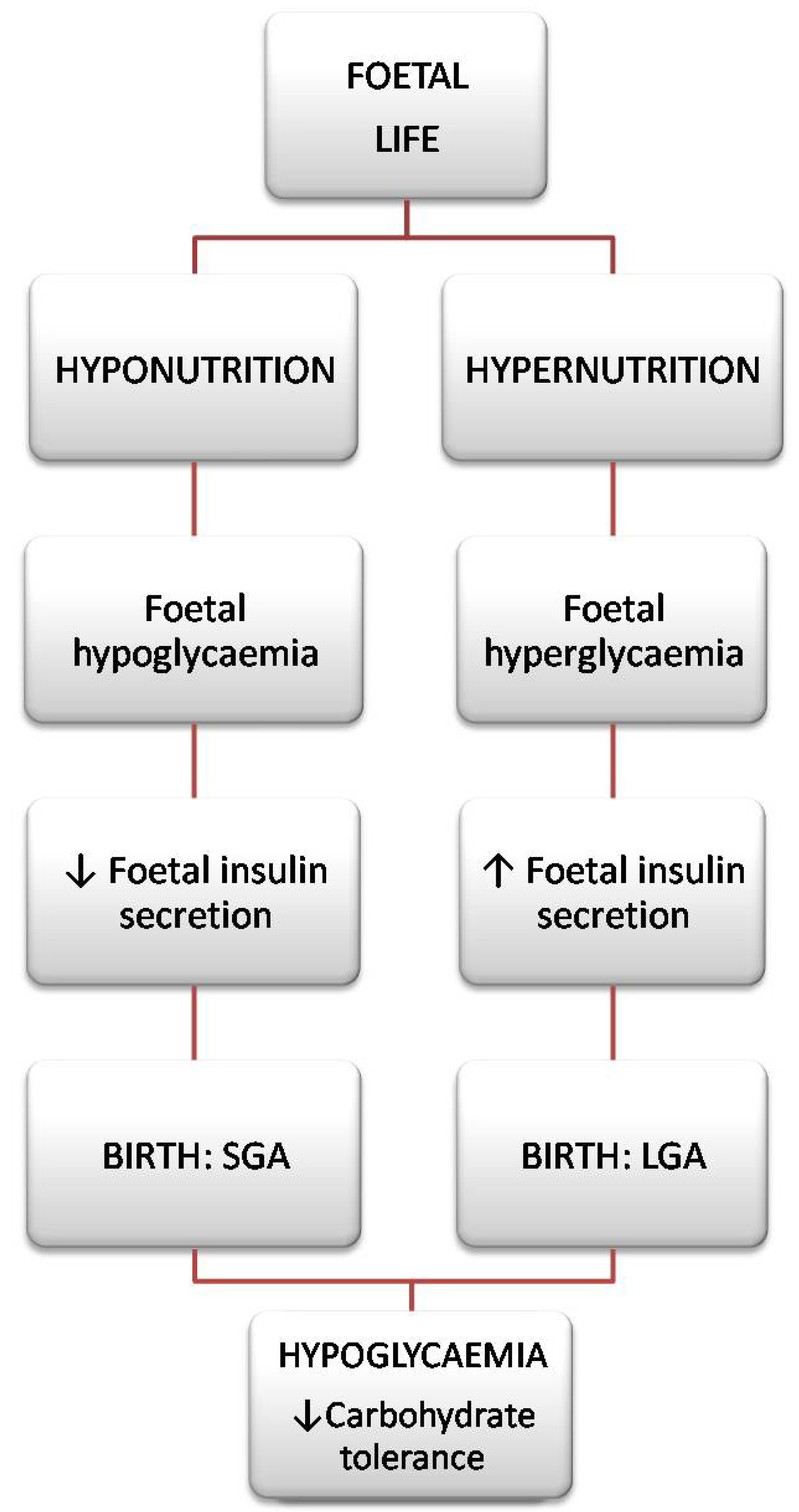
3. The Foetal Hypothesis: Everything Begins at the Beginning
4. SGA & LGA: Are They Different?

5. Metabolites and Metabolomics in SGA and LGA

{kind=link}
| Author | Population study | Sample | Metabolomic analysis | Metabolites results |
|---|---|---|---|---|
| van Vliet et al. 2013 [32] | 10 IUGR vs. 6 control rabbit fetuses | Brains | LC-QTOF-MS | Aspargine, ornithine, N-acetylaspartylglutamic acid, N-acetylaspartate and palmitoleic acid |
| Lin et al. 2012 [33] | 18 IUGR vs. 18 control pig fetuses | Umbilical vein plasma | Q-TOF MS | Pyroglutamic acid, carnitine and creatinine |
| Favretto et al. 2012 [34] | 22 IUGR vs. 21 control human neonates | Cord blood | LC-HRMS | Phenilalanine, tryptophan acid and glutamate |
| Logan et al. 2012 [28] | 18 IDM vs. 12 healthy term control infants | Urine | NMR | Glucose, formate, fumarate, succinate and citrate |
| Dessì et al. 2011 [35] | 26 IUGR vs. 30 control human neonates | Urine | NMR | Myo-inositol, sarcosine, carnitine and creatinine |
| Horgan et al. 2011 [36] | 8 IUGR vs. 6 control human neonates | Venous cord plasma | UPLC-MS | Phenylacetylglutamine, carnitine, hydroxybutyrate |
| Nissen et al. 2011 [37] | 12 IUGR vs. 12 control newborn piglets | Plasma | NMR | Myo-inositol and D-chiro-inositol |
6. Conclusions
Conflicts of Interest
References
- Nicholson, J.K.; Connelly, J.; Lindon, J.C.; Holmes, E. Metabonomics: A platform for studying drug toxicity and gene function. Nat. Rev. Drug. Discov. 2002, 1, 153–161. [Google Scholar] [CrossRef]
- Ellis, D.I.; Dunn, W.B.; Griffin, J.L.; Allwood, J.W.; Goodacre, R. Metabolic fingerprinting as a diagnostic tool. Pharmacogenomics 2007, 8, 1243–1266. [Google Scholar] [CrossRef]
- Kim, O.Y.; Lee, J.H.; Sweeney, G. Metabolomic profiling as a useful tool for diagnosis and treatment of chronic disease: Focus on obesity, diabetes and cardiovascular diseases. Expert. Rev. Cardiovasc. Ther. 2013, 11, 61–68. [Google Scholar] [CrossRef]
- Smith, S.C., Jr.; Collins, A.; Ferrari, R.; Holmes, D.R., Jr.; Logstrup, S.; McGhie, D.V.; Ralston, J.; Sacco, R.L.; Stam, H.; Taubert, K.; et al. Our time: A call to save preventable death from cardiovascular disease (heart disease and stroke). J. Am. Coll. Cardiol. 2012, 60, 2343–2348. [Google Scholar] [CrossRef]
- Bozzola, M.; Bozzola, E.; Abela, S.; Amato, S. Childhood obesity: Know it to prevent it. Ig. Sanita Pubbl. 2012, 68, 473–482. [Google Scholar]
- Chambers, J.C.; Elliott, P.; Zabaneh, D.; Zhang, W.; Li, Y.; Froguel, P.; Balding, D.; Scott, J.; Kooner, J.S. Common genetic variation near MC4R is associated with waist circumference and insulin resistance. Nat. Genet. 2008, 40, 716–718. [Google Scholar] [CrossRef]
- Hindorff, L.A.; Sethupathy, P.; Junkins, H.A.; Ramos, E.M.; Mehta, J.P.; Collins, F.S.; Manolio, T.A. Potential etiologic and functional implications of genome-wide association loci for human diseases and traits. Proc. Natl. Acad. Sci. USA 2009, 106, 9362–9367. [Google Scholar] [CrossRef]
- Cai, G.; Cole, S.A.; Butte, N.; Bacino, C.; Diego, V.; Tan, K.; Göring, H.H.; O’Rahilly, S.; Farooqi, I.S.; Comuzzie, A.G. A quantitative trait locus on chromosome 18q for physical activity and dietary intake in Hispanic children. Obesity (Silver Spring) 2006, 14, 1596–1604. [Google Scholar] [CrossRef]
- Meng, Q.; Mäkinen, V.P.; Luk, H.; Yang, X. Systems biology approaches and applications in obesity, diabetes, and cardiovascular diseases. Curr. Cardiovasc. Risk. Rep. 2013, 7, 73–83. [Google Scholar] [CrossRef]
- Barker, D.J. Fetal origins of coronary heart disease. BMJ 1995, 311, 171–174. [Google Scholar] [CrossRef]
- Hales, C.N.; Barker, D.J. Type 2 (non insulin dipendent) diabetes mellitus: The trifty phenotype hypothesis. Diabetologia 2003, 35, 395–601. [Google Scholar]
- Dessì, A.; Ottonello, G.; Fanos, V. Physiopathology of intrauterine growth retardation: From classic data to metabolomics. J. Matern. Fetal. Neonatal. Med. 2012, 25, 13–18. [Google Scholar] [CrossRef]
- Dyck, R.F.; Klomp, H.; Tan, L. From “thrifty genotype” to “hefty fetal phenotype”: The relationship between high birthweight and diabetes in Saskatchewan Registered Indians. Can. J. Public Health 2001, 92, 340–344. [Google Scholar]
- Harder, T.; Rodekamp, E.; Schellong, K.; Dudenhausen, J.W.; Plagemann, A. Birth weight and subsequent risk of type 2 diabetes: A meta-analysis. Am. J. Epidemiol. 2007, 165, 849–857. [Google Scholar] [CrossRef]
- Diderholm, B. Perinatal energy metabolism with reference to IUGR & SGA: Studies in pregnant women & newborn infants. Indian J. Med. Res. 2009, 130, 612–617. [Google Scholar]
- Committee on Practice Bulletins Gynecology; American College of Obstetricians and Gynecologists. Intrauterine growth restriction. Clinical management guidelines for obstetrician-gynecologists. Int. J.Gynaecol. Obstet. 2001, 72, 85–96. [Google Scholar] [CrossRef]
- Fanos, V.; Puddu, M.; Reali, A.; Atzei, A.; Zaffanello, M. Perinatal nutrient restriction reduces nephron endowment increasing renal morbidity in adulthood: A review. Early Hum. Dev. 2010, 86, 37–42. [Google Scholar] [CrossRef]
- Ju, H.; Chadha, Y.; Donovan, T.; O’Rourke, P. Fetal Macrosomia and pregnancy outcomes. Aust. N. Z. J. Obstet. Gynaecol. 2009, 49, 504–509. [Google Scholar] [CrossRef]
- King, J.C. Physiology of pregnancy and nutrient metabolism. Am. J. Clin. Nutr. 2000, 7, 1218–1225. [Google Scholar]
- Lam, Y.Y.; Hatzinikolas, G.; Weir, J.M.; Janovská, A.; McAinch, A.J.; Game, P.; Meikle, P.J.; Wittert, G.A. Insulin-stimulated glucose uptake and pathways regulating energy metabolism in skeletal muscle cells: The effects of subcutaneous and visceral fat, and long-chain saturated, n-3 and n-6 polyunsaturated fatty acids. Biochim. Biophys. Acta 2011, 1811, 468–475. [Google Scholar] [CrossRef]
- Holemans, K.; Aerts, L.; van Assche, F.A. Lifetime consequences of abnormal fetal pancreatic development. J. Physiol. 2003, 547, 11–20. [Google Scholar] [CrossRef]
- Fernandez-Twinn, D.S.; Ozanne, S.E. Mechanisms by which poor early growth programs type-2 diabetes, obesity and the metabolic syndrome. Physiol. Behav. 2006, 88, 234–243. [Google Scholar] [CrossRef]
- Plagemann, A.; Harder, T.; Rake, A.; Voits, M.; Fink, H.; Rohde, W.; Dörner, G. Perinatal elevation of hypothalamic insulin, acquired malformation of hypothalamic galaninergic neurons, and syndrome X-like alterations in adulthood of neonatally overfed rats. Brain Res. 1999, 836, 146–155. [Google Scholar] [CrossRef]
- Pantaleão, L.C.; Teodoro, G.F.; Torres-Leal, F.L.; Vianna, D.; de Paula, T.D.; de Matos-Neto, E.M.; Trindade, M.C.; Rogero, M.M.; Bueno, C.R., Jr.; Tirapegui, J. Maternal postnatal high-fat diet, rather than gestational diet, affects morphology and mTOR pathway in skeletal muscle of weaning rat. J. Nutr. Biochem. 2013, 24, 1340–1348. [Google Scholar] [CrossRef]
- Persson, B.; Westgren, M.; Celsi, G.; Nord, E.; Ortqvist, E. Leptin concentrations in cord blood in normal newborn infants and offspring of diabetic mothers. Horm. Metab. Res. 1999, 3, 467–471. [Google Scholar]
- Ibáñez, L.; Sebastiani, G.; Diaz, M.; Gómez-Roig, M.D.; Lopez-Bermejo, A.; de Zegher, F. Low body adiposity and high leptinemia in breast-fed infants born small-for-gestational-age. J. Pediatr. 2010, 156, 145–147. [Google Scholar] [CrossRef]
- Heltemes, A.; Gingery, A.; Soldner, E.L.; Bozadjieva, N.; Jahr, K.N.; Johnson, B.K.; Gilbert, J.S. Chronic placental ischemia alters amniotic fluid milieu and results in impaired glucose tolerance, insulin resistance and hyperleptinemia in young rats. Exp. Biol. Med. (Maywood) 2010, 235, 892–899. [Google Scholar] [CrossRef]
- Logan, K.M.; Wijeyesekera, A.D.; Perez, I.G.; Hyde, M.J.; Romero, M.G.; Jeffries, S.; Andreas, N.; Gale, C.; Holmes, E.; Modi, N. Infants of mothers with diabetes have altered urinary metabolic profile at birth. Available online: http://www.neonatalsociety.ac.uk/abstracts/logank_2012_diabeticurinaryprofile.shtml/ (accessed on 1 July 2013).
- Oresic, M.; Simell, S.; Sysi-Aho, M.; Näntö-Salonen, K.; Seppänen-Laakso, T.; Parikka, V.; Katajamaa, M.; Hekkala, A.; Mattila, I.; Keskinen, P.; et al. Dysregulation of lipid and amino acid metabolism precedes islet autoimmunity in children who later progress to type 1 diabetes. J. Exp. Med. 2008, 205, 2975–2984. [Google Scholar] [CrossRef]
- Sysi-Aho, M.; Ermolov, A.; Gopalacharyulu, P.V.; Tripathi, A.; Seppänen-Laakso, T.; Maukonen, J.; Mattila, I.; Ruohonen, S.T.; Vähätalo, L.; Yetukuri, L.; et al. Metabolic regulation in progression to autoimmune diabetes. PLoS Comput. Biol. 2011, 7, e1002257. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, Y.; Hao, F.; Zhou, X.; Han, X.; Tang, H.; Ji, L. Human serum metabonomic analysis reveals progression axes for glucose intolerance and insulin resistance statuses. J. Proteome Res. 2009, 8, 5188–5195. [Google Scholar] [CrossRef]
- Van Vliet, E.; Eixarch, E.; Illa, M.; Arbat-Plana, A.; González-Tendero, A.; Hogberg, H.T.; Zhao, L.; Hartung, T.; Gratacos, E. Metabolomics reveals metabolic alterations by intrauterine growth restriction in the fetal rabbit brain. PLoS One 2013, 8, e64545. [Google Scholar] [CrossRef]
- Lin, G.; Liu, C.; Feng, C.; Fan, Z.; Dai, Z.; Lai, C.; Li, Z.; Wu, G.; Wang, J. Metabolomic analysis reveals differences in umbilical vein plasma metabolites between normal and growth-restricted fetal pigs during late gestation. J. Nutr. 2012, 142, 990–998. [Google Scholar] [CrossRef]
- Favretto, D.; Cosmi, E.; Ragazzi, E.; Visentin, S.; Tucci, M.; Fais, P.; Cecchetto, G.; Zanardo, V.; Viel, G.; Ferrara, S.D. Cord blood metabolomic profiling in intrauterine growth restriction. Anal. Bioanal. Chem. 2012, 402, 1109–1121. [Google Scholar] [CrossRef]
- Dessì, A.; Atzori, L.; Noto, A.; Visser, G.H.; Gazzolo, D.; Zanardo, V.; Barberini, L.; Puddu, M.; Ottonello, G.; Atzei, A.; et al. Metabolomics in newborns with intrauterine growth retardation (IUGR): Urine reveals markers of metabolic syndrome. J. Matern. Fetal. Neonatal. Med. 2011, 2, 35–39. [Google Scholar]
- Horgan, R.P.; Broadhurst, D.I.; Dunn, W.B.; Brown, M.; Heazell, A.E.; Kell, D.B.; Baker, P.N.; Kenny, L.C. Changes in the metabolic footprint of placental explant-conditioned medium cultured in different oxygen tensions from placentas of small for gestational age and normal pregnancies. Placenta 2010, 31, 893–901. [Google Scholar] [CrossRef]
- Nissen, P.M.; Nebel, C.; Oksbjerg, N.; Bertram, H.C. Metabolomics reveals relationship between plasma inositols and birth weight: Possible markers for fetal programming of type 2 diabetes. J. Biomed. Biotechnol. 2011, 2011, 1147–1155. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Dessì, A.; Puddu, M.; Ottonello, G.; Fanos, V. Metabolomics and Fetal-Neonatal Nutrition: Between “Not Enough” and “Too Much”. Molecules 2013, 18, 11724-11732. https://doi.org/10.3390/molecules181011724
Dessì A, Puddu M, Ottonello G, Fanos V. Metabolomics and Fetal-Neonatal Nutrition: Between “Not Enough” and “Too Much”. Molecules. 2013; 18(10):11724-11732. https://doi.org/10.3390/molecules181011724
Chicago/Turabian StyleDessì, Angelica, Melania Puddu, Giovanni Ottonello, and Vassilios Fanos. 2013. "Metabolomics and Fetal-Neonatal Nutrition: Between “Not Enough” and “Too Much”" Molecules 18, no. 10: 11724-11732. https://doi.org/10.3390/molecules181011724
APA StyleDessì, A., Puddu, M., Ottonello, G., & Fanos, V. (2013). Metabolomics and Fetal-Neonatal Nutrition: Between “Not Enough” and “Too Much”. Molecules, 18(10), 11724-11732. https://doi.org/10.3390/molecules181011724