Gelam Honey Protects against Gamma-Irradiation Damage to Antioxidant Enzymes in Human Diploid Fibroblasts

Abstract
:1. Introduction
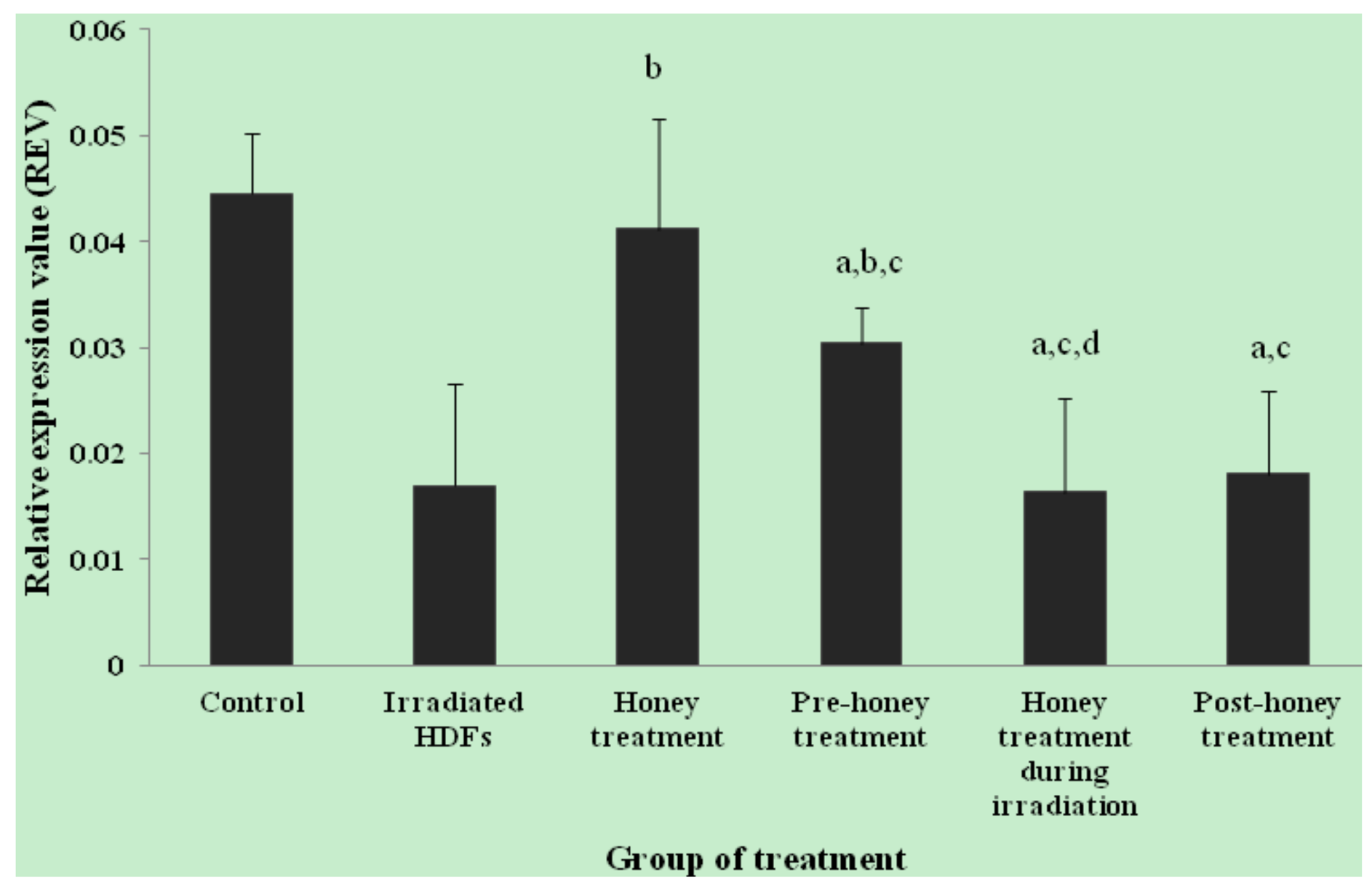
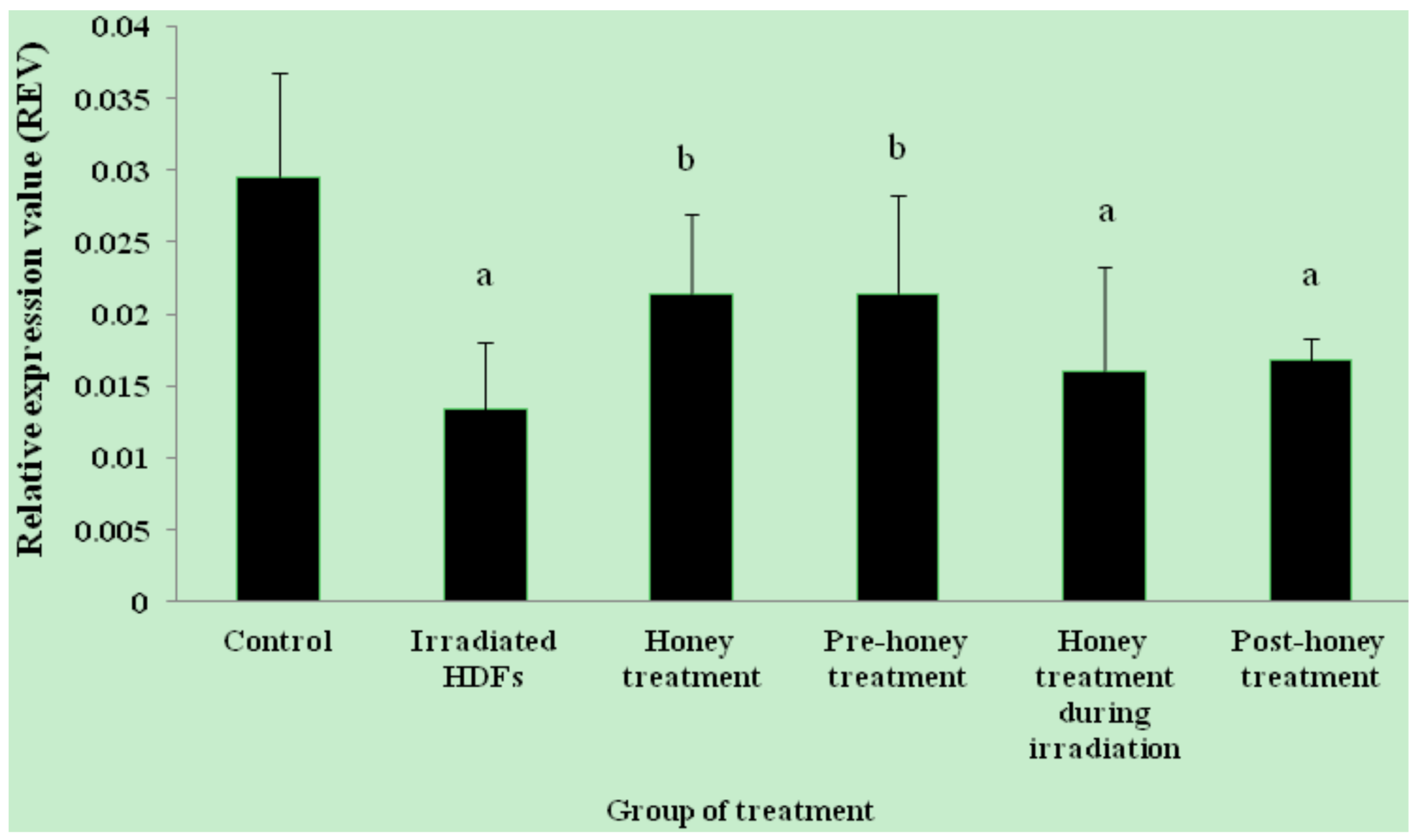
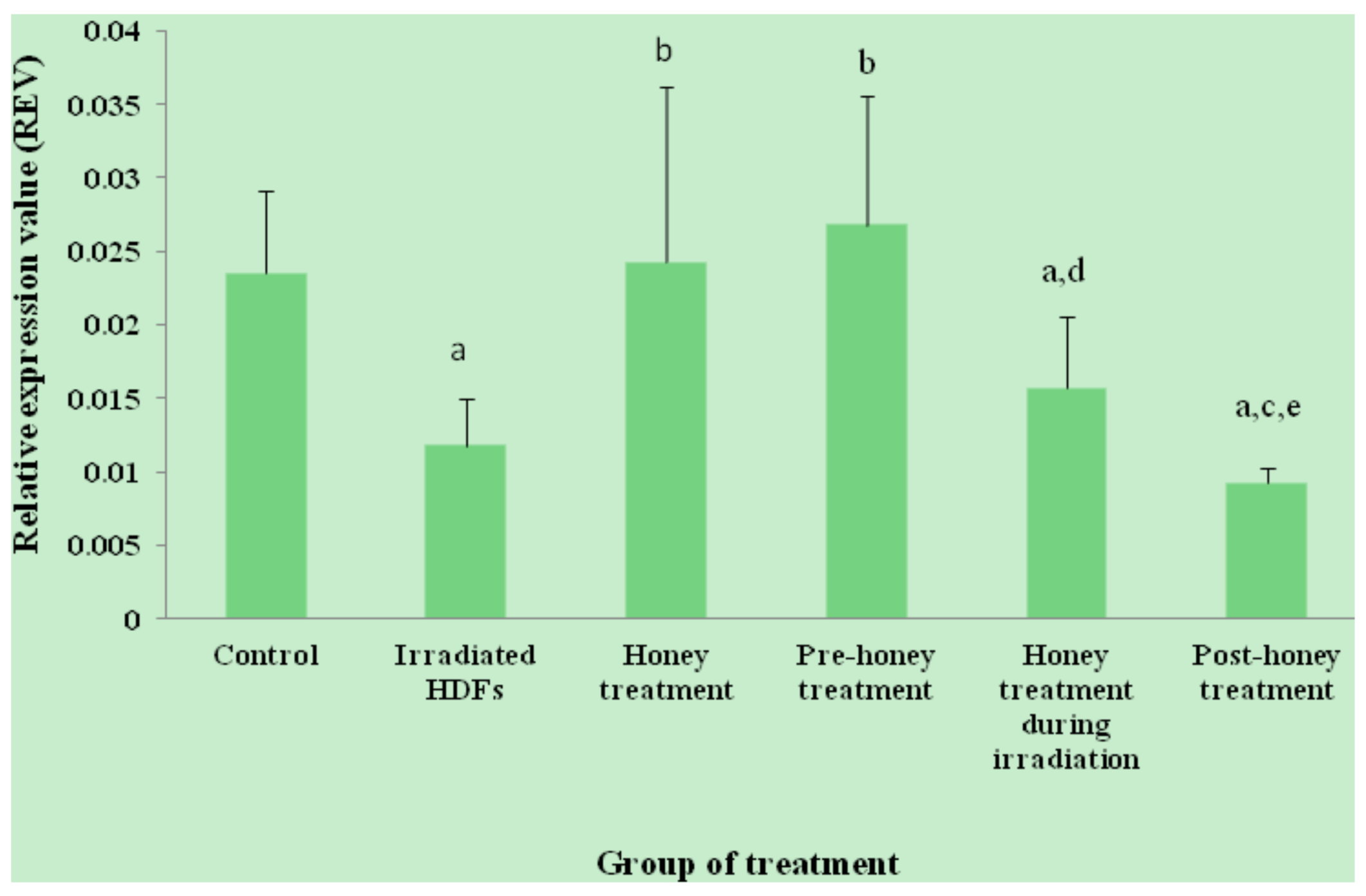
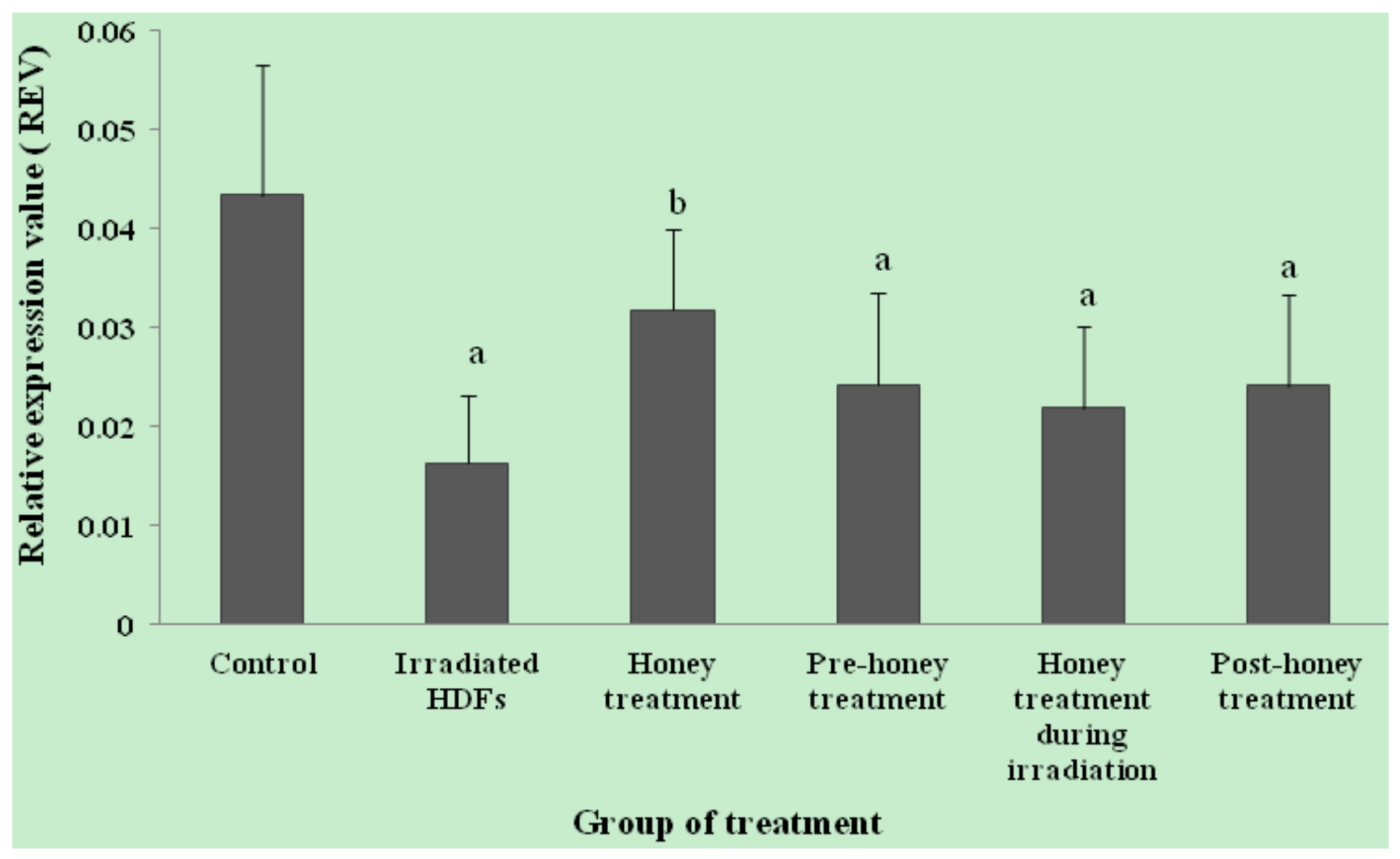
2. Results
2.1. SOD1, SOD2, CAT and GPx1 Gene Expression
2.2. Antioxidant Enzyme Activity
3. Discussion
4. Experimental
4.1. Sterilization of Gelam Honey
4.2. Cell Culture Protocol
4.3. Honey Treatment Protocol
4.4. Exposure to Gamma-Irradiation
4.5. Primer Design
4.6. RNA Extraction
4.7. Quantitative Real-Time RT-PCR
4.8. Protein Extraction
4.9. Enzyme Extraction
4.10. Superoxide Dismutase (SOD) Assay
4.11. Catalase (CAT) Assay
4.12. Glutathione Peroxidase (GPx) Assay
4.13. Statistical Analysis
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- U.S. Environmental Protection Agency (US EPA). Radiation protection; 2011. Available online: http://www.epa.gov/radiation/sources/#natural/ (accessed on 18 January 2013).
- Washington State Department of Health (WSDH). Background radiation natural versus man made; 2002. Available online: http://www.doh.wa.gov/Portals/1/Documents/Pubs/320-063_bkvsman_fs.pdf (accessed on 18 January 2013).
- World Health Organization (WHO). Human Exposure to Ionizing Radiation. 2011. http://www.who.int/hac/crises/jpn/faqs/en/index2.html/ (accessed on 18 January 2013).
- World Nuclear Association (WNA). Nuclear Power in the World Today. 2012. Available online: http://www.world-nuclear.org/ (accessed on 18 January 2013).
- Baskar, R. Emerging role of radiation induced bystander effects: Cell communications and carcinogenesis. Genome Integr. 2010, 1, 13. [Google Scholar] [CrossRef] [PubMed]
- Held, K.D.; Willer, H. Molecular and Cellular Basis of Radiation Injury. In Human Radiation Injury; Shrieve, D.C., Loeffler, J.S., Eds.; Lippincott Williams & Wilkins, Wolters Klumer: Philadelphia, PA, USA, 2011; Volume 1, pp. 1–13. [Google Scholar]
- Sun, J.; Chen, Y.; Li, M.; Ge, Z. Role of antioxidant enzymes on ionizing radiation Resistance. Free Radic. Biol. Med. 1998, 24, 586–593. [Google Scholar] [CrossRef]
- Limon-Pacheco, J.; Gonsebatt, M.E. The role of antioxidants and antioxidant-related enzymes in protective responses to environmentally induced oxidative stress. Mutat. Res. 2009, 674, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Hosseinimehr, S.J.; Nemati, A. Radioprotective effects of hesperidin against gamma irradiation in mouse bone marrow cells. Br. J. Radiol. 2006, 79, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Prouillac, C.; Célariès, B.; Vicendo, P.; Rima, G. Evaluation, in vitro, of the radioprotection of DNA from gamma-rays by naphazoline. C. R. Biol. 2006, 329, 196–199. [Google Scholar] [CrossRef] [PubMed]
- Weydert, C.J.; Cullen, J.J. Measurement of superoxide dismutase, catalase and glutathione peroxidase in cultures cells and tissue. Nat. Protoc. 2010, 5, 51–66. [Google Scholar] [CrossRef] [PubMed]
- Warters, R.L.; Packardb, A.T.; Kramera, G.F.; Gaffneya, D.K.; Moos, P.J. Differential gene expression in primary human skin keratinocytes and fibroblasts in response to ionizing radiation. Radiat. Res. 2009, 172, 82–95. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Naser, M.B.; Abdallah, M.; Almeida, H.L.D., Jr.; Wollin, U. Human skin cell culture and its impact on dermatology. Egypt. Dermatol. J. 2005, 1, 1–25. [Google Scholar]
- Cmielová, J.; Havele, R.; Jiroutová, A.; Kohlerová, R.; Seifrtová, M.; MUthná, D.; Vávrová, J.; Øezáèová, M. DNA damage caused by ionizing radiation in embryonic diploid fibroblasts WI-38 induces both apoptosis and senescence. Physiol. Res. 2005, 60, 667–677. [Google Scholar]
- Makpol, S.; Ahmad, T.A.F.A.; Jubri, Z.; Rajab, N.; Yusof, N.; Yusof, Y.A.M. Gelam honey acting as a radioprotectant agent in gamma-irradiated human diploid fibroblasts. J. Med. Plants Res. 2012, 6, 129–138. [Google Scholar]
- Westbury, C.B.; Sahlberg, K.K.; Borresen-Dale, A.L.; Isacke, C.M.; Yarnold, J.R. Gene expression profiling of human dermal fibroblasts exposed to bleomycin sulphate does not differentiate between radiation sensitive and control patients. Radiat. Oncol. 2011, 6, 42. [Google Scholar] [CrossRef] [PubMed]
- Zelko, I.N.; Mariani, T.J.; Folz, R.J. Superoxide dismutase multigene family: A comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2), AND EC-SOD (SOD3) gene structures, evolution and expression. Free Radic. Biol. Med. 2002, 33, 337–349. [Google Scholar] [CrossRef]
- Hrdanz, E.R.; Kahl, R. Alterations of antioxidant enzyme expression in response to hydrogen peroxide. Free Radic. Biol. Med. 1998, 24, 27–38. [Google Scholar] [CrossRef]
- Somosy, Z. Radiation response of cell organelles. Micron 2000, 31, 165–181. [Google Scholar] [CrossRef]
- Davis, G.D.J.; Masilamoni, J.G.; Arul, V.; Kumar, M.S.M.; Baraneedharan, U.; Paul, S.F.D.; Sakthivelu, I.V.; Jesudason, E.P.; Jayakumar, R. Radioprotective effect of DL-α-lipoic acid on mice skin fibroblasts. Cell Biol. Toxicol. 2008, 25, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.A.; Lee, I.K.; Zhang, R.; Piao, M.J.; Kim, K.C.; Kim, S.Y.; Shin, T.; Kim, B.J.; Lee, N.H.; Hyun, J.W. Radioprotective effect of geraniin via the inhibition of apoptosis triggered by γ-radiation-induced oxidative stress. Cell Biol. Toxicol. 2011, 27, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Borek, C. Antioxidants and radiation therapy. J. Nutr. 2004, 134, 3207S–3209S. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K.; Srinivasan, V.; Toles, R.; Karikari, P.; Seed, T.; Papas, V.K.; Hyatt, J.A.; Kumar, K.S. Radiation Protection by the Antioxidant Alpha-Tocopherol Succinate. NATO RTG-009. 2005. http://www.usuhs.mil/afrri/outreach/pdf/kumar_NATO_2005.pdf (accessed on 18 January 2013).
- Okunieff, P.; Swarts, S.; Keng, P.; Sun, W.; Wang, W.; Kim, J.; Yang, S.; Zhang, H.; Liu, C.; Williams, J.P.; et al. Antioxidants reduce consequences of radiation exposure. Adv. Exp. Med. Biol. 2008, 614, 165–178. [Google Scholar] [PubMed]
- Biswal, B.M.; Zakaria, A.; Ahmad, N.M. Topical application of honey in the management of radiation mucositis. A Preliminary study. Support. Care Cancer. 2003, 11, 242–248. [Google Scholar] [PubMed]
- Hussein, S.Z.; Yusoff, K.M.; Makpol, S.; Yusof, Y.A.M. Antioxidant capacities and total phenolic contents increase with gamma irradiation in two types of Malaysian honey. Molecules 2011, 16, 6378–6395. [Google Scholar] [CrossRef] [PubMed]
- Aljadi, A.M.; Kamaruddin, M.Y. Evaluation of the phenolic contents and antioxidant capacities of two Malaysian flora honeys. Food Chem. 2004, 85, 513–518. [Google Scholar] [CrossRef]
- Yao, L.K.; Razak, S.L.A.; Ismail, N.; Fai, N.C.; Asgar, M.H.A.M.; Sharif, N.M.; Aan, G.J.; Jubri, Z. Malaysian gelam honey reduces oxidative damage and modulates antioxidant enzyme activities in young and middle aged rats. J. Med. Plants Res. 2011, 5, 5618–5625. [Google Scholar]
- Park, E.M.; Ramnath, N.; Yang, G.Y.; Ahn, J.Y.; Park, Y.; Lee, T.Y.; Shin, H.S.; Yu, J.; Ip, C.; Park, Y.M. High SOD and low GPX activities in RBC predict susceptibility of lung cancer patients to radiation pneumonitis. Free Radic. Biol. Med. 2007, 42, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Cemeli, E.; Baumgartner, A.; Anderson, D. Antioxidants and the comet assay. Mutan. Res. 2005, 681, 51–67. [Google Scholar] [CrossRef] [PubMed]
- Beyer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Aebi, H. Calatase in vitro. Meth. Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Panglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar]
Sample Availability: Samples of Malaysian Gelam honey are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group of Treatment | Enzyme specific activity (mU/mg protein) | ||
|---|---|---|---|
| SOD | CAT | GPx | |
| Control | 1020.85 + 133.18 | 0.45 + 0.09 | 26.22 + 3.49 |
| Irradiated HDFs | 487.45 + 123.75 a | 0.32 + 0.05 a | 12.64 + 2.51 a |
| Honey treatment | 759.83 + 98.74 a,b | 0.47 + 0.13 b | 16.59 + 3.38 a,b |
| Pre-honey treatment | 772.06 + 206.36 a,b | 0.42 + 0.07 b | 13.12 + 2.92 a |
| Honey treatment during radiation | 665.04 + 114.80 a,b | 0.35 + 0.12 | 8.74 + 1.40 a,b,c,d |
| Post-honey treatment | 591.84 + 90.99 a,c | 0.36 + 0.08 | 6.16 + 0.50 a,b,c,e |
| Gene | Sequence (5'–3') | Size of PCR product (bp) |
|---|---|---|
| GAPDH (Forward) | TCCCTGAGCTGAACGGGAAG | 217 |
| GAPDH (Reverse) | GGAGGAGTGGGTGTCGCTGT | |
| SOD1 (Forward) | GAAGGTGTGGGGAAGCATTA | 174 |
| SOD1 (Reverse) | ACATTGCCCAAGTCTCCAAC | |
| SOD2 (Forward) | CGTCACCGAGGAGAAGTACC | 312 |
| SOD2 (Reverse) | CTGATTTGGACAAGCAGCAA | |
| CAT (Forward) | CGTGCTGAATGAGGAACAGA | 119 |
| CAT(Reverse) | AGTCAGGGTGGACCTCAGTG | |
| GPx1 (Forward) | CCAAGCTCATCACCTGGTCT | 127 |
| GPx1 (Reverse) | TCGATGTCAATGGTCTGGAA |
© 2013 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ahmad, T.A.F.T.; Jubri, Z.; Rajab, N.F.; Rahim, K.A.; Yusof, Y.A.M.; Makpol, S. Gelam Honey Protects against Gamma-Irradiation Damage to Antioxidant Enzymes in Human Diploid Fibroblasts. Molecules 2013, 18, 2200-2211. https://doi.org/10.3390/molecules18022200
Ahmad TAFT, Jubri Z, Rajab NF, Rahim KA, Yusof YAM, Makpol S. Gelam Honey Protects against Gamma-Irradiation Damage to Antioxidant Enzymes in Human Diploid Fibroblasts. Molecules. 2013; 18(2):2200-2211. https://doi.org/10.3390/molecules18022200
Chicago/Turabian StyleAhmad, Tengku Ahbrizal Farizal Tengku, Zakiah Jubri, Nor Fadilah Rajab, Khairuddin Abdul Rahim, Yasmin Anum Mohd Yusof, and Suzana Makpol. 2013. "Gelam Honey Protects against Gamma-Irradiation Damage to Antioxidant Enzymes in Human Diploid Fibroblasts" Molecules 18, no. 2: 2200-2211. https://doi.org/10.3390/molecules18022200