Anti-Escherichia coli O157:H7 Properties of Purple Prairie Clover and Sainfoin Condensed Tannins


Abstract
:1. Introduction
2. Results
2.1. Protein-Precipitating Capacities of PPC and SF CT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteins | Parameters * | PPC | SF | SEM ǂ | P values |
|---|---|---|---|---|---|
| BSA | a0+a | 3.0 | 3.0 | 0.01 | 0.213 |
| b | 258 | 368 | 30.0 | 0.584 | |
| c | 176 | 316 | 9.9 | <0.001 | |
| PP | 233 | 364 | 9.4 | <0.001 | |
| Rubisco | a0+a | 3.0 | 2.2 | 0.02 | <0.001 |
| b | 335 | 863 | 22.1 | <0.001 | |
| c | 50 | 195 | 13.4 | 0.002 | |
| PP | 299 | 852 | 16.4 | <0.001 |

2.2. Reactivity of PPC and SF CT with Liposome
| Time | Source | CT concentration (µg/mL) | SEM * | P value (L ǂ) | |||
|---|---|---|---|---|---|---|---|
| 0 | 10 | 50 | 200 | ||||
| 10 min | Control | 286 | 5.7 | ||||
| PPC | 207 | 184 | 178 | <0.001 | |||
| SF | 203 | 195 | 168 | <0.001 | |||
| 30 min | Control | 257 | 7.8 | ||||
| PPC | 198 | 183 | 167 | <0.001 | |||
| SF | 191 | 190 | 166 | <0.001 | |||
2.3. Minimum Inhibitory Concentration (MIC) of the CT
| Species | Strains | MIC (CT µg/mL) | |
|---|---|---|---|
| PPC | SF | ||
| E. coli | ATCC 25922 | 20–40 | 100–150 |
| ATCC 35352 | 20 | 125 | |
| E. coli O157:H7 | 3081 | 20 | 100 |
| H4420N | 20–50 | 120 | |
| E318N | 20–30 | 110 | |
| EDL933 | 20 | 120 | |
| R508N2006 | 20 | 110 | |
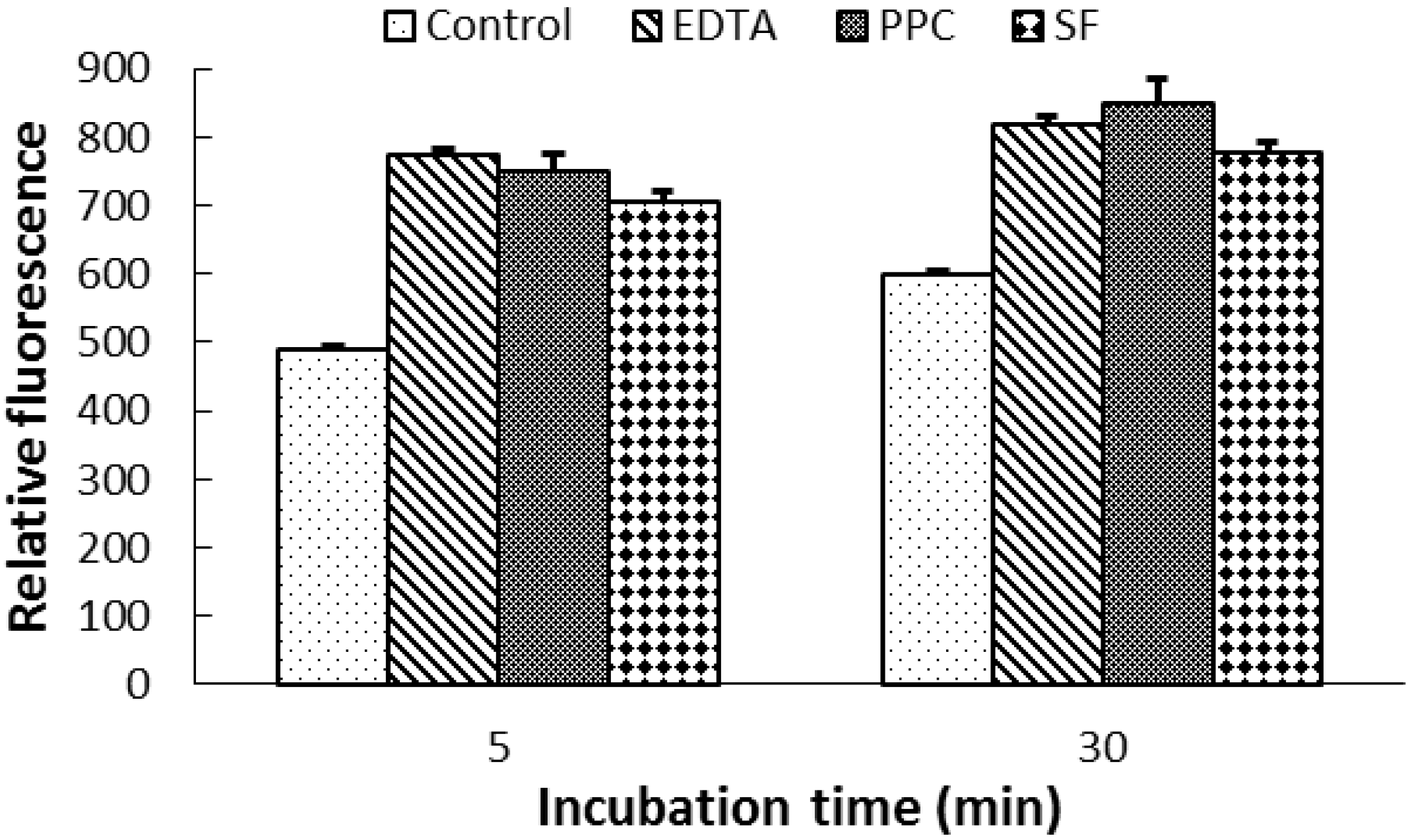
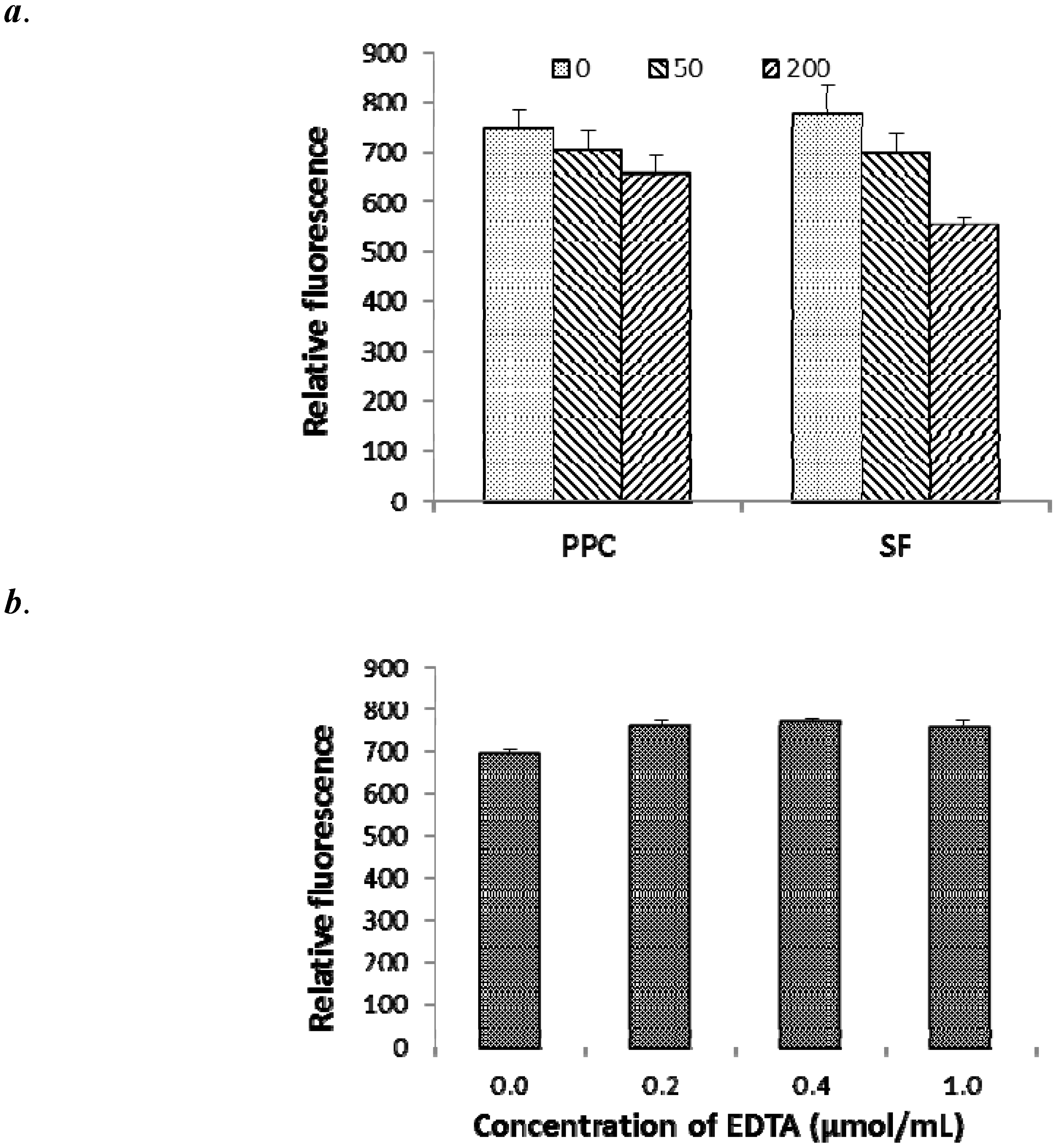
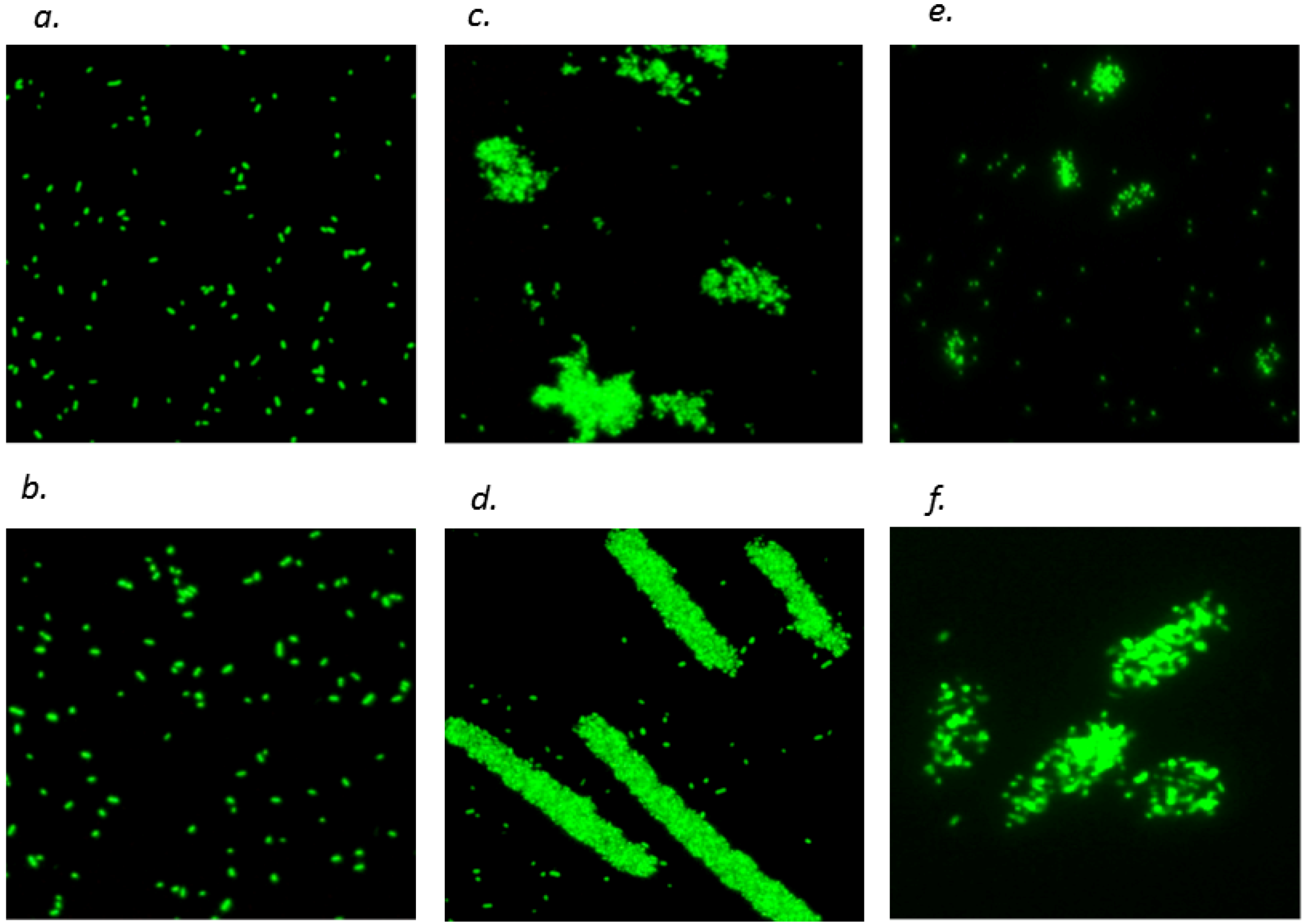
2.4. Effects of CT on OM Permeability and Cell Aggregation of E. coli


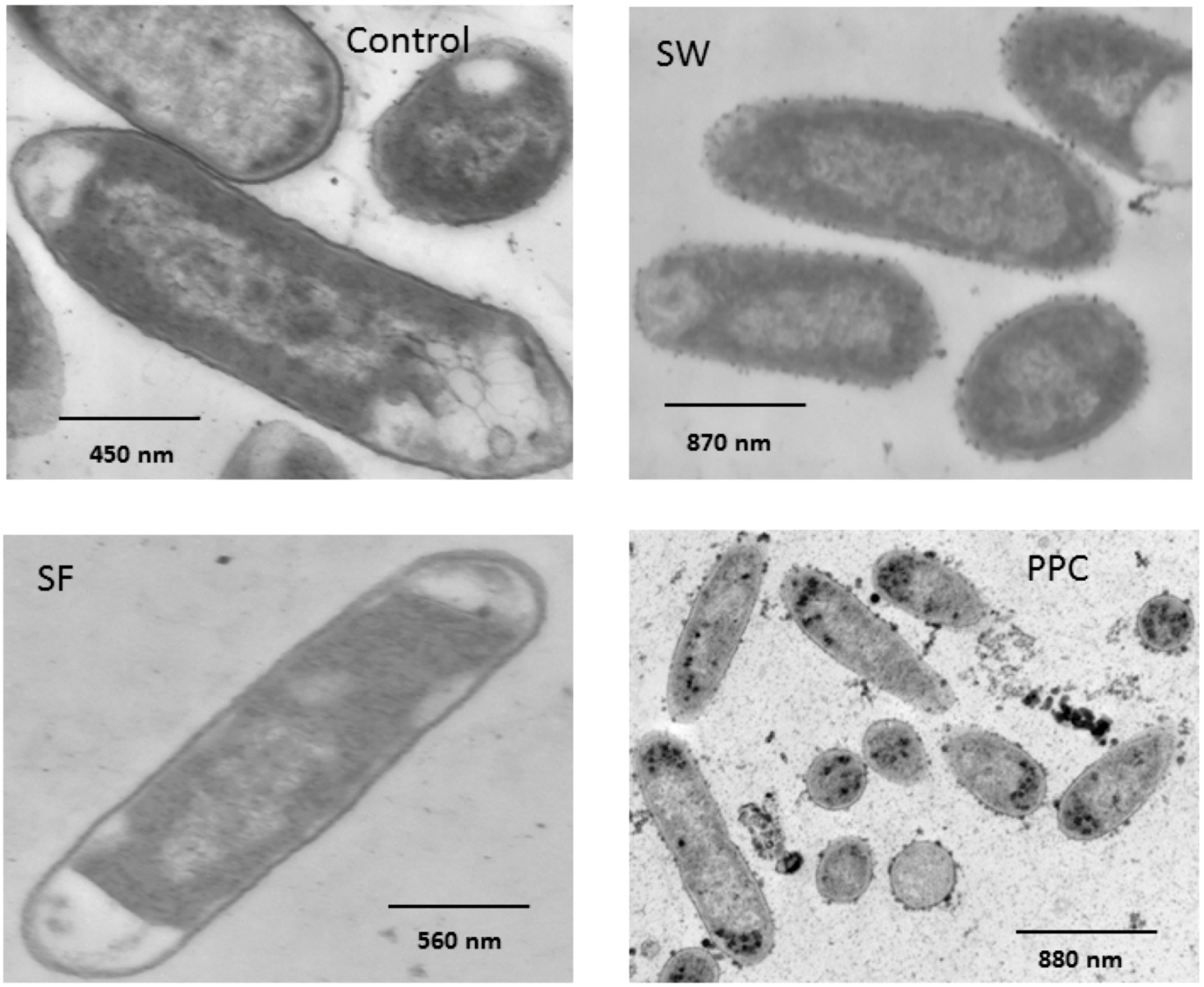
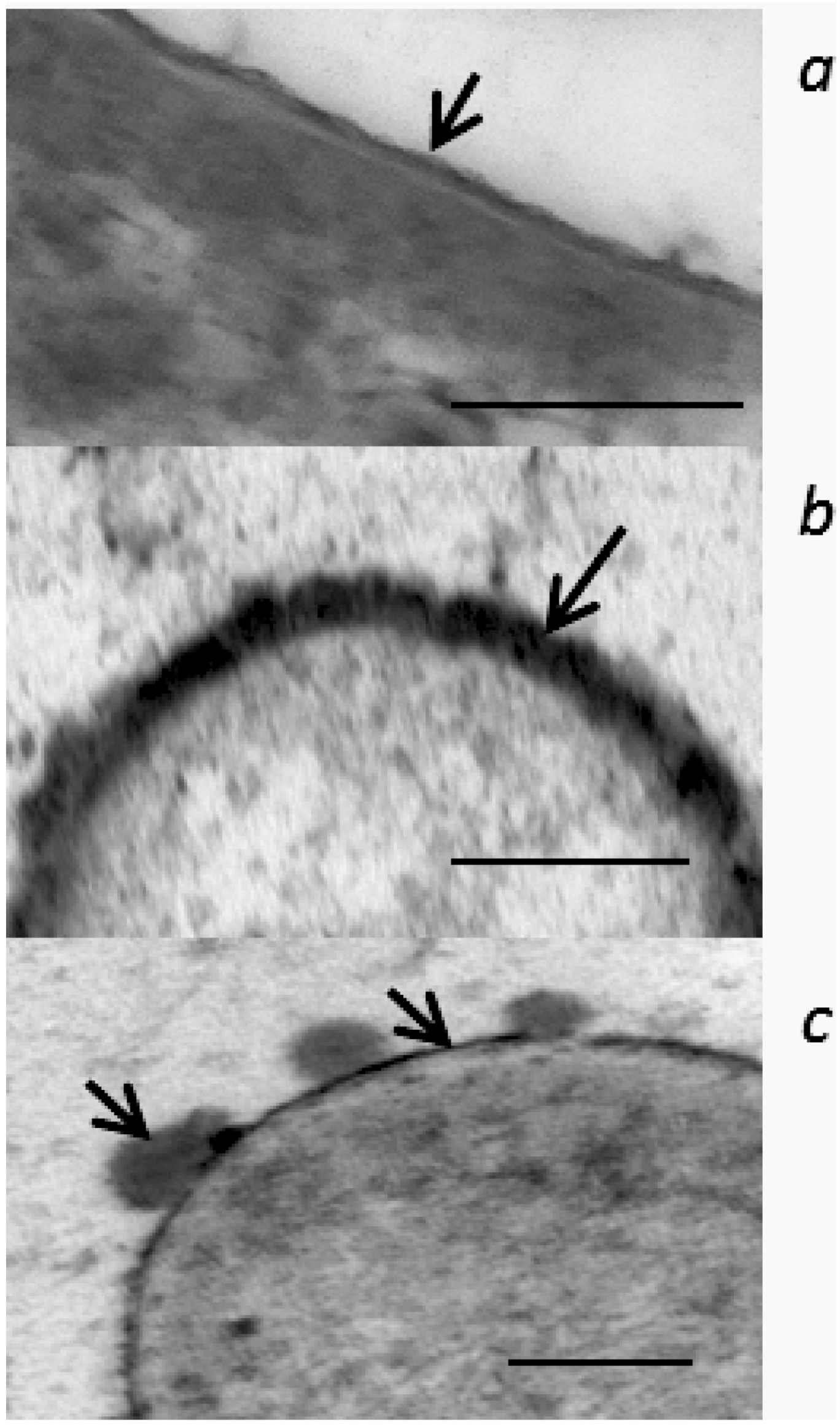
2.5. Morphological Alteration of Cell Membrane



3. Discussion
4. Experimental
4.1. Isolation and Purification of PPC and SF CT
4.2. Determination of the Protein-Precipitating Capacities of PPC and SF CT
4.3. Determination of Reactivity of PPC and SF CT with Liposome
4.4. Minimum Inhibitory Concentration (MIC) of CT
4.4.1. Pre-incubation of E. coli and E. coli O157:H7
4.4.2. Determination of MIC
4.5. Determination of the Effects of CT on OM Permeability and Cell Aggregation of E. coli
4.5.1. Pre-culturing and Preparation of Bacterial Suspension
4.5.2. OM Permeability Assays
4.5.3. Determination of Cell Aggregation by CT
4.5.3.1. NPN Fluorescence Quenching Assay
4.5.3.2. Fluorescence Imaging Analysis
4.6. Transmission Electron Microscopy Examination of Morphological Alteration of Cell Membrane
4.7. Data Calculations and Statistical Analysis
5. Conclusions
Acknowledgments
Conflicts of Interest
- Sample Availability: Samples of the PPC CT and SF CT are available from the authors.
References
- Elder, R.O.; Keen, J.E.; Siragusa, G.R.; Barkocy-Gallagher, G.A.; Koohmaraie, M.; Laegreid, W.W. Correlation of enterohemorrhagic Escherichia coli O157:H7 prevalence in feces, hides, and carcasses of beef cattle during processing. Proc. Natl. Acad. Sci. USA 2000, 97, 2999–3003. [Google Scholar]
- Stanford, K.; McAllister, T.A.; Niu, D.Y.; Stephens, T.P.; Mazzocco, A.; Waddell, T.E.; Johnson, R.P. Oral delivery systems for encapsulated bacteriophages targeted at Escherichia coli O157:H7 in feedlot cattle. J. Food Prot. 2010, 73, 1304–1312. [Google Scholar]
- Callaway, T.R.; Carr, M.A.; Edrington, T.S.; Anderson, R.C.; Nisbet, D.J. Diet, Escherichia coli O157:H7, and cattle: A review after 10 years. Curr. Issues Mol. Biol. 2009, 11, 67–80. [Google Scholar]
- Min, B.R.; Pinchak, W.E.; Anderson, R.C.; Callaway, T.R. Effect of tannins on the in vitro growth of Escherichia coli O157:H7 and in vivo growth of generic Escherichia coli excreted from steers. J. Food. Prot. 2007, 70, 543–550. [Google Scholar]
- Wang, Y.; Xu, Z.; Bach, S.B.; McAllister, T.A. Sensitivity of Escherichia coli O157:H7 to seaweed (Ascophyllum nodosum) phlorotannins and terrestrial tannins. Asian-Aust. J. Anim. Sci. 2009, 22, 238–245. [Google Scholar]
- Wang, Y.; Jin, L.; Ominski, K.H.; He, M.; Xu, Z.; Krause, D.O.; Acharya, S.N.; Wittenberg, K.M.; Liu, X.L.; Stanford, K.; McAllister, T.A. Screening of condensed tannins from Canadian Prairie forages for anti-Escherichia coli O157:H7 with an emphasis on purple prairie clover (Dalea purpurea Vent). J. Food Prot. 2013, in press. [Google Scholar]
- Berard, N.C.; Wang, Y.; Wittenberg, K.M.; Krause, D.O.; Coulman, D.E.; McAllister, T.A.; Ominski, K.H. Condensed tannin concentrations found in vegetative and mature forage legumes grown in western Canada. Can. J. Plant Sci. 2011, 91, 669–675. [Google Scholar] [CrossRef]
- Jin, L.; Wang, Y.; Iwaasa, A.D.; Xu, Z.; Schellenberg, M.P.; Zhang, Y.G.; Liu, X.L.; McAllister, T.A. Effect of condensed tannins on ruminal degradability of purple prairie clover (Dalea purpurea Vent.) harvested at two growth stages. Anim. Feed Sci. Technol. 2012, 176, 17–25. [Google Scholar]
- Haslam, E. Plant polyphenols (syn. vegetable tannins) and chemical defence-a reappraisal. J. Chem. Ecol. 1988, 14, 1789–1805. [Google Scholar] [CrossRef]
- Ikigai, H.; Nakae, T.; Hara, Y.; Shimamura, T. Bactericidal catechins damage the lipid bilayer. Biochem. Biophys. Acta 1993, 1147, 132–136. [Google Scholar] [CrossRef]
- Jones, G.A.; McAllister, T.A.; Muir, A.; Cheng, K.J. Effects of sainfoin (Onobrychis viciifolia scop) condensed tannins on growth and proteolysis by four strains of ruminal bacteria. Appl. Environ. Microbiol. 1994, 60, 1374–1378. [Google Scholar]
- Min, B.R.; Attwood, G.T.; McNabb, W.C.; Molan, A.L.; Barry, T.N. The effect of condensed tannins from Lotus corniculatus on the proteolytic activities and growth of rumen bacteria. Anim. Feed Sci. Technol. 2005, 121, 45–48. [Google Scholar] [CrossRef]
- Min, B.R.; Pinchak, W.E.; Merkel, R.; Walker, S.; Tomita, G.; Anderson, R.C. Comparative antimicrobial activity of tannin extracts from perennial plants on mastitis pathogens. Sci. Res. Essays 2008, 3, 66–73. [Google Scholar]
- Haslam, E. Plant Polyphenols, Vegetable Tannins Revisited; Cambridge University Press: Cambridge, UK, 1989. [Google Scholar]
- Tsuchiya, H. Stereospecificity in membrane effects of catechins. Chem-Biol. Interact. 2001, 134, 41–54. [Google Scholar] [CrossRef]
- Cox, S.D.; Mann, C.M.; Markham, J.L.; Gustafson, J.E.; Warmington, J.R.; Wyllie, S.G. Determining the antimicrobial actions of tea tree oil. Molecules 2001, 6, 87–91. [Google Scholar] [CrossRef]
- Nohynek, L.J.; Alakomi, H.L.; Kähkönen, M.P.; Heinonen, M.; Helander, I.M.; Oksman-Caldentey, K.M.; Puupponen-Pimiä, R.H. Berry phenolics, antimicrobial properties and mechanisms of action against severe human pathogens. Nutr. Cancer 2006, 54, 18–32. [Google Scholar] [CrossRef]
- Pasqua, R.D.; Betts, G.; Hoskins, N.; Edwards, M.; Ercolini, D.; Mauriello, G. Membrane toxicity of antimicrobial compounds from essential oils. J. Agric. Food Chem. 2007, 55, 4863–4870. [Google Scholar] [CrossRef]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef]
- Cohen, G.N. Microbial Biochemistry, 2nd ed; Dordrecht, Springer: New York, NY, USA, 2011. [Google Scholar]
- McAllister, T.A.; Martinez, T.; Bae, H.D.; Muir, A.D.; Yanke, L.J.; Jones, G.A. Characterization of condensed tannins purified from legume forages, chromophore production, protein precipitation, and inhibitory effects on cellulose digestion. J. Chem. Ecol. 2005, 31, 2049–2068. [Google Scholar] [CrossRef]
- Shimamura, T.; Zhao, W.H.; Hu, Z.Q. Mechanism of action and potential for use of tea catechin as an anti-infective agent. Anti-Infect. Agents Med. Chem. 2007, 6, 57–62. [Google Scholar] [CrossRef]
- Gea, A.; Stringano, E.; Brown, R.H.; Mueller-Harvey, I. In situ analysis and structural elucidation of sainfoin (Onobrychis viciifolia) tannins for high-throughput germplasm screening. J. Agric. Food Chem. 2011, 59, 495–503. [Google Scholar]
- Theodoridou, K.; Aufrère, J.; Andueza, D.; Le Morvan, A.; Picard, F.; Stringano, E.; Pourrat, J.; Mueller-Harvey, I.; Baumont, R. Effect of plant development during first and second growth cycle on chemical composition, condensed tannins and nutritive value of three sainfoin (Onobrychis viciifolia) varieties and lucerne. Grass Forage Sci. 2011, 66, 402–414. [Google Scholar] [CrossRef]
- Hayat, M.A. Principles and Techniques of Electron Microscopy, Biological Applications; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Ibrahim, H.R.; Sugimoto, Y.; Aoki, T. Ovotransferrin antimicrobial peptide (OTAT-92) kills bacteria through a membrane damage mechanism. Biochem. Biophys. Acta 2000, 1523, 196–205. [Google Scholar] [CrossRef]
- Helander, I.M.; Alakomi, H.L.; Latva-Kala, K.; Mattila-Sandholm, T.; Pol, I.; Smid, E.J.; Gorris, L.G.M.; Von Wright, A. Characterization of the action of selected essential oil components on gram-negative bacteria. J. Agric. Food Chem. 1998, 46, 3590–3595. [Google Scholar]
- Je, J.Y.; Kim, S.K. Chitosan derivatives killed bacteria by disrupting the outer and inner membrane. J. Agric. Food Chem. 2006, 54, 6629–6633. [Google Scholar] [CrossRef]
- Helander, I.M.; Mattila-Sandholm, T. Fluorometric assessment of gram-negative bacterial permeabilization. J. Appl. Microbiol. 2000, 88, 213–219. [Google Scholar] [CrossRef]
- Hancock, R.E.W. Alterations in outer membrane permeability. Ann. Rev. Microbiol. 1984, 38, 237–264. [Google Scholar] [CrossRef]
- Scalbert, A. Antimicrobial properties of tannins, review article number 63. Phytochemistry 1991, 30, 3875–3883. [Google Scholar] [CrossRef]
- Annuk, H.; Hirmo, S.; Türi, E.; Mikelsaar, M.; Arak, E.; Wadström, T. Effect on cell surface hydrophobicity and susceptibility of Helicobacter pylori to medicinal plant extracts. FEMS Microbiol. Lett. 1999, 172, 41–45. [Google Scholar] [CrossRef]
- Türi, M.; Türi, E.; Kõljalg, S.; Mikelsaar, M. Influence of aqueous extracts of medicinal plants on surface hydrophobicity of Escherichia coli strains of different origin. APMIS 1997, 105, 956–962. [Google Scholar] [CrossRef]
- Kuehn, M.J.; Kesty, N.C. Bacterial outer membrane vesicles and the host-pathogen interaction. Genes Dev. 2005, 19, 2645–2655. [Google Scholar] [CrossRef]
- Rahman, M.; Kühn, I.; Rahman, M.; Olsson-Liljequist, B.; Möllby, R. Evaluation of a scanner-assisted colorimetric MIC method for susceptibility testing of gram-negative fermentative bacteria. Appl. Environ. Microb. 2004, 70, 2398–2403. [Google Scholar]
- Chung, K.T.; Lu, Z.; Chou, M.W. Mechanism of inhibition of tannic acid and related compounds on the growth of intestinal bacteria. Food Chem. Toxicol. 1998, 36, 1053–1060. [Google Scholar] [CrossRef]
- Holowachuk, S.A.; Bal’a, M.F.; Buddington, R.K. A kinetic microplate method for quantifying the antibacterial properties of biological fluids. J. Microbiol. Meth. 2003, 55, 441–446. [Google Scholar] [CrossRef]
- Mustaffa, F.; Indurkar, J.; Ismail, S.; Shah, M.; Mansor, S.M. An antimicrobial compound isolated from Cinnamomum iners leaves with activity against methicillin-resistant Staphylococcus aureus. Molecules 2011, 16, 3037–3047. [Google Scholar]
- Bach, S.J.; Wang, Y.; McAllister, T.A. Effect of seaweed (Ascophyllum nodosum) on fecal shedding of Escherichia coli O157:H7 by feedlot cattle and on growth performance of lambs. Anim. Feed Sci. Technol. 2008, 142, 17–32. [Google Scholar] [CrossRef]
- Berney, M.; Hammes, F.; Bosshard, F.; Weilenmann, H.U.; Egli, T. Assessment and interpretation of bacterial viability by using the LIVE/DEAD BacLight Kit in combination with flow cytometry. Appl. Environ. Microbiol. 2007, 73, 3283–3290. [Google Scholar]
- Bae, H.D.; McAllister, T.A.; Yanke, L.J.; Cheng, K.-J.; Muir, A. D. Effects of condensed tannins on endoglucanase activity and filter paper digestion by Fibrobacter succinogenes S85. Appl. Environ. Microbiol. 1993, 59, 2131–2138. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, X.-L.; Hao, Y.-Q.; Jin, L.; Xu, Z.-J.; McAllister, T.A.; Wang, Y. Anti-Escherichia coli O157:H7 Properties of Purple Prairie Clover and Sainfoin Condensed Tannins. Molecules 2013, 18, 2183-2199. https://doi.org/10.3390/molecules18022183
Liu X-L, Hao Y-Q, Jin L, Xu Z-J, McAllister TA, Wang Y. Anti-Escherichia coli O157:H7 Properties of Purple Prairie Clover and Sainfoin Condensed Tannins. Molecules. 2013; 18(2):2183-2199. https://doi.org/10.3390/molecules18022183
Chicago/Turabian StyleLiu, Xiu-Li, Yong-Qing Hao, Long Jin, Zhong-Jun Xu, Tim A. McAllister, and Yuxi Wang. 2013. "Anti-Escherichia coli O157:H7 Properties of Purple Prairie Clover and Sainfoin Condensed Tannins" Molecules 18, no. 2: 2183-2199. https://doi.org/10.3390/molecules18022183