Metabolomic Analysis of Methyl Jasmonate-Induced Triterpenoid Production in the Medicinal Herb Centella asiatica (L.) Urban


Abstract
:1. Introduction
2. Results and Discussion
2.1. Partial Characterization and Fractionation of the Plant Extracts by TLC Analysis
2.2. Dynamic Changes in the Metabolome of MeJa-treated C. asiatica Cells



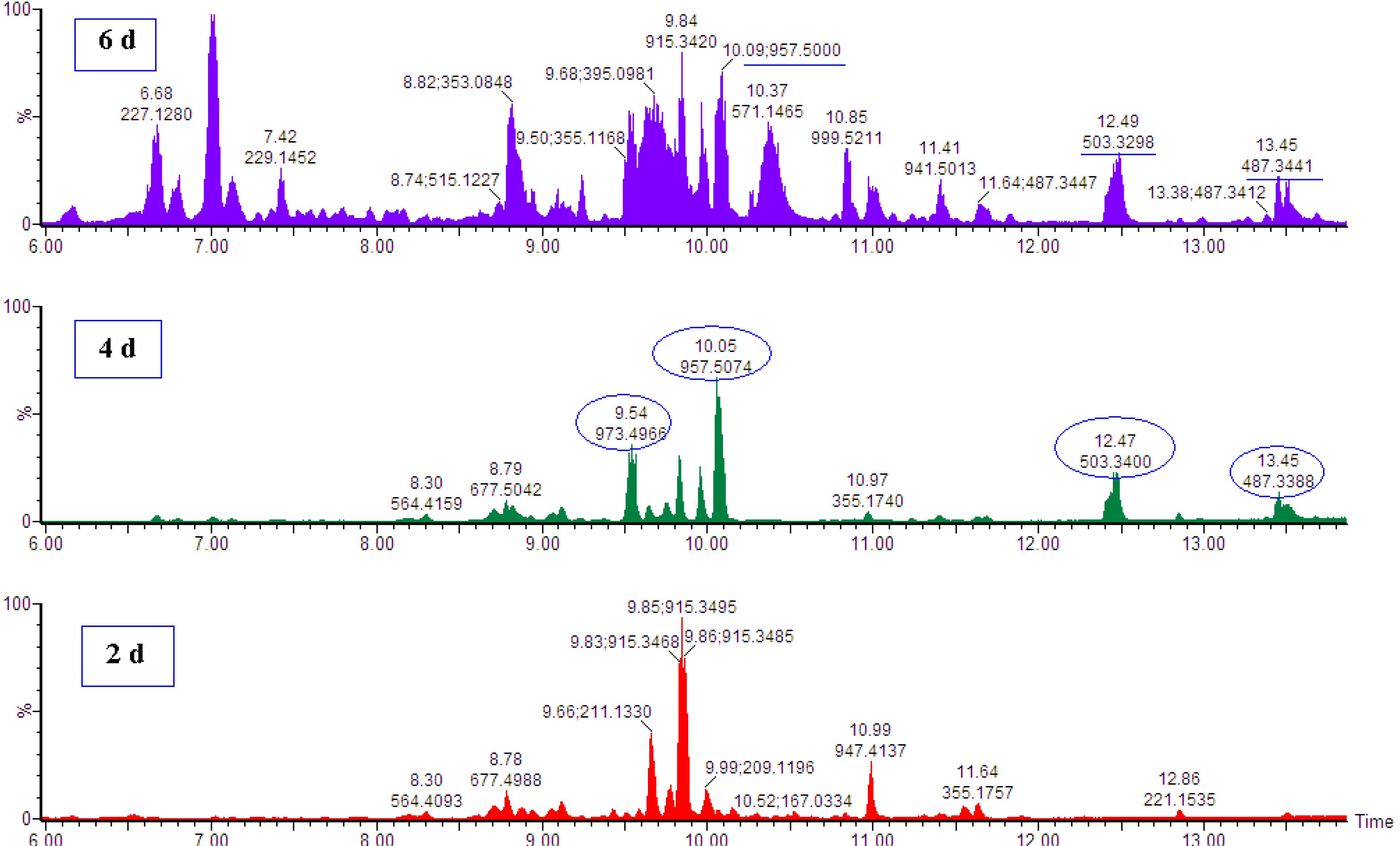{kind=link}
{kind=link}
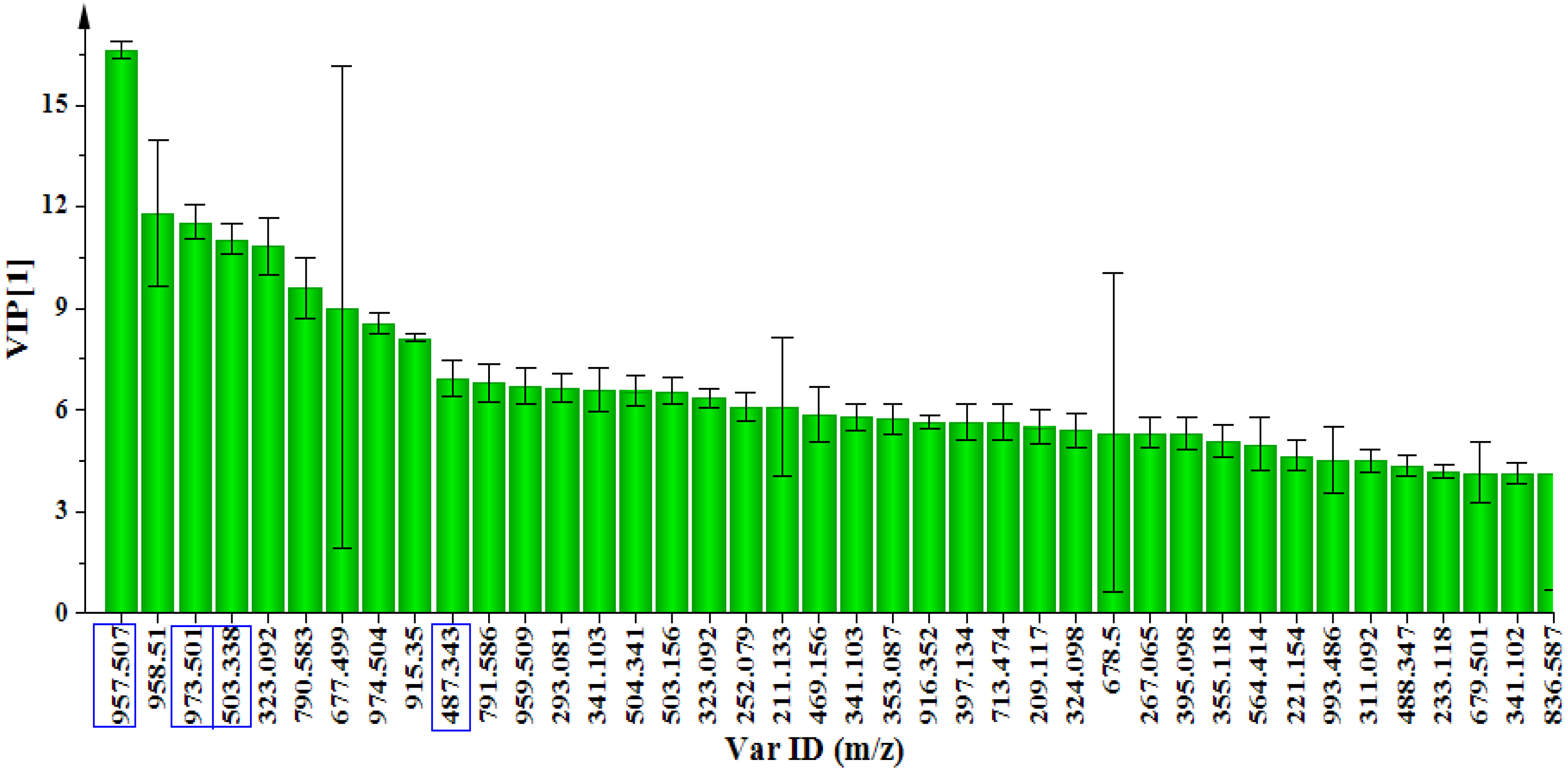{kind=link}
| Rt (min) | Observed mass (m/z) | Calculated mass (m/z) | Ion | Identification | Molecular Formula | Molecular Weight (g/mol) | Correlation with MeJa effect |
|---|---|---|---|---|---|---|---|
| 10.16 | 533.1980 | 533.1970 | [M+Na−H]− | Gibberellin 2-O-beta-d-glucoside | C25H34O11 | 509.53 | Negative |
| 10.05 | 957.5070 | 957.5059 | [M−H]− | Asiaticoside * | C48H78O19 | 959.12 | Positive |
| 9.54 | 973.5010 | 973.5008 | [M−H]− | Madecassoside * | C48H78O20 | 975.12 | Positive |
| 12.47 | 503.3380 | 503.3373 | [M−H]− | Madecassic acid * | C30H48O6 | 504.70 | Positive |
| 9.79 | 207.1380 | 207.1385 | [M−H]− | 3-Hydroxy-9-apo-delta-caroten-9-one | C13H20O2 | 208.29 | Positive |
| 8.94 | 323.0920 | 323.0919 | [M+FA+Na−H]− | 7,4'-Dihydroxy-8-methylflavan | C16H16O3 | 256.29 | Negative |
| 13.50 | 487.3430 | 487.3424 | [M−H]− | Asiatic acid * | C30H48O5 | 488.70 | Positive |
| 9.23 | 341.1030 | 341.1025 | [M−H]− | 4',5,6,7-Tetramethoxyflavone | C19H18O6 | 342.34 | Negative |
| 8.83 | 353.0870 | 353.0873 | [M−H]− | Chlorogenate | C16H18O9 | 354.31 | Positive |
| 7.09 | 227.1280 | 227.1283 | [M+FA-H]− | Geranyl formate | C11H18O2 | 182.26 | Positive |
| 9.69 | 395.0980 | 395.0970 | [M+Na_Na-H]− | Feruloylserotonin | C20H20N2O4 | 352.38 | Positive |
| 9.52 | 355.1180 | 355.1182 | [M−H]− | Kievitone | C20H20O6 | 356.37 | Negative |
| 14.74 | 221.1540 | 221.1542 | [M−H]− | Rishitin | C14H22O2 | 222.32 | Positive |
| 10.06 | 993.4860 | 993.4848 | [M+NaCl_HCOONa−H]− | Pheophytin a | C55H72N4O5 | 869.18 | Positive |
| 9.07 | 311.0920 | 311.0909 | [M−H]− | Baicalein 5,6,7-trimethyl ether | C18H16O5 | 312.31 | Negative |
| 9.87 | 251.1280 | 251.1283 | [M−H]− | Ubiquinol | C14H20O4 | 252.30 | Positive |
| 10.13 | 251.0700 | 251.0783 | [M−H]− | 4'-O-Methylisoflavone | C16H12O3 | 252.26 | Negative |
| 10.65 | 531.1500 | 531.1495 | [M−H]− | Flavonol 3-O-d-xylosylglucoside | C26H28O12 | 532.49 | Positive |
| 9.86 | 263.1290 | 263.1279 | [M−H]− | Abscisate | C15H20O4 | 264.31 | Positive |
| 10.99 | 265.1440 | 265.1433 | [M+FA−H]− | Glutinosone | C14H20O2 | 220.30 | Positive |
| 9.63 | 293.0820 | 293.0814 | [M−H]− | Dehydrocycloguanandin | C18H14O4 | 294.30 | Negative |
| 7.25 | 345.1340 | 345.1338 | [M−H]− | Gibberellin A3/6/29/34-catabolite | C19H22O6 | 346.37 | Negative |
| 8.84 | 351.0710 | 351.0716 | [M−H]− | 4-Methylumbelliferone glucuronide | C16H16O9 | 352.29 | Positive |
| 8.98 | 383.1130 | 383.1113 | [M−H]− | Anthocyanin 3'-O-beta-d-glucoside | C21H20O7 | 384.38 | Negative |
| 11.69 | 942.5160 | 941.5110 | [M−H]− | Soyasaponin I | C48H78O18 | 943.12 | Positive |
| 14.53 | 476.2780 | 476.2777 | [M−H]− | LysoPE(0:0/18:2(9Z,12Z)) | C23H44NO7P | 477.57 | Positive |
| 8.95 | 209.1170 | 209.1145 | [M−H]− | Jasmonic acid | C12H18O3 | 210.27 | Positive |
| 13.37 | 485.3280 | 485.3297 | [M−H]− | Gypsogenate | C30H46O5 | 486.68 | Positive |
| 11.40 | 488.3470 | 488.3435 | [M−H]− | Bayogenin | C30H48O5 | 488.70 | Positive |

2.3. MeJa Treatment of C. asiatica Cells Enhances the Production of Triterpenoids
| Triterpenoid | Rt (min) | m/z | Time point (day) | Concentration (μg/g wet weight) Control Treated | |
|---|---|---|---|---|---|
| Madecassoside | 9.49 | 973.51 | 2 | 0.388 ± 0.022 | 0.416 ± 0.024 |
| 4 | 0.426 ± 0.044 | 10.580 ± 0.634 | |||
| 6 | 0.424 ± 0.008 | 9.869 ± 9.235 | |||
| Asiaticoside | 10.05 | 957.51 | 2 | 0.669 ± 0.051 | 0.480 ± 0.146 |
| 4 | 0.542 ± 0.093 | 20.934 ± 6.089 | |||
| 6 | 0.727 ± 0.087 | 20.819 ± 5.685 | |||
| Madecassic acid | 12.41 | 503.33 | 2 | 0.515 ± 0.459 | 0.472 ± 0.489 |
| 4 | 0.473 ± 0.450 | 8.065 ± 9.435 | |||
| 6 | 0.539 ± 0.514 | 5.854 ± 6.381 | |||
| Asiatic acid | 13.47 | 487.34 | 2 | 0.258 ± 0.019 | 0.266 ± 0.011 |
| 4 | 0.205 ± 0.010 | 4.859 ± 0.342 | |||
| 6 | 0.317 ± 0.047 | 2.595 ± 0.096 | |||
3. Experimental
3.1. Preparation and Elicitation of Cell Cultures
3.2. Cell Viability Assessment
3.3. Metabolite Extraction
3.4. TLC Analysis of Ethanolic Extracts
3.5. Ultra-High Performance Liquid Chromatography–High Definition Mass Spectrometry (UHPLC-HDMS) Analysis
3.6. Data Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
References
- Hall, R.D. Plant metabolomics: From holistic hope to hype, to hot topic. New Phytol. 2005, 169, 453–468. [Google Scholar] [CrossRef]
- Shulaev, V.; Cortes, D.; Miller, G.; Mittler, R. Metabolomics for plant stress response. Physiol. Plant. 2008, 132, 199–208. [Google Scholar] [CrossRef]
- Suzuki, H.; Srinivasa-Reddy, M.S.; Naoumkina, M.; Aziz, N.; May, G.D.; Huhman, D.V.; Sumner, L.W.; Blount, J.W.; Mendes, P.; Dixon, R.A. Methyl jasmonate and yeast elicitor induce differential transcriptional and metabolic re-programming in cell suspension cultures of the model legume Medicago truncatula. Planta 2005, 220, 696–707. [Google Scholar] [CrossRef]
- James, J.T.; Dubery, I.A. Pentacyclic triterpenoids from the medicinal herb, Centella asiatica (L.) Urban. Molecules 2009, 14, 3922–3941. [Google Scholar] [CrossRef]
- Mangas, S.; Bonfill, M.; Osuna, L.; Moyano, E.; Tortoriello, J.; Cusido, R.M.; Pinol, M.T.; Palaźon, J. The effect of methyl jasmonate on triterpene and sterol metabolisms of Centella asiatica, Ruscus aculeatus and Galphimia glauca cultured plants. Phytochemistry 2006, 67, 2041–2049. [Google Scholar] [CrossRef]
- Hernandez-Vazquez, L.; Bonfill, M.; Moyana, E.; Cusido, R.M.; Navarro-Ocańa, A.; Palazon, J. Conversion of α-amyrin into centellosides by plant cell cultures of Centella asiatica. Biotechnol. Lett. 2010, 32, 315–319. [Google Scholar]
- Kim, O.T.; Ahn, J.C.; Hwang, S.J.; Hwang, B. Cloning and expression of a farnesyl diphosphate synthase in Centella asiatica (L.) Urban. Mol. Cell 2005, 19, 294–299. [Google Scholar] [CrossRef]
- Kim, O.T.; Seong, N.S.; Kim, M.Y.; Hwang, B. Isolation and characterisation of squalene synthase cDNA from Centella asiatica (L.) Urban. J. Plant Biol. 2005, 48, 263–269. [Google Scholar] [CrossRef]
- Kim, O.T.; Kim, M.Y.; Huh, S.M.; Bai, D.G.; Ahn, J.C.; Hwang, B. Cloning of a cDNA probably encoding oxidosqualene cyclase associated with asiaticoside biosynthesis from Centella asiatica (L.) Urban. Plant Cell Rep. 2005, 24, 304–311. [Google Scholar] [CrossRef]
- Cheong, J.-J.; Choi, Y.D. Methyl jasmonate as a vital substance in plants. Trends Genet. 2003, 19, 409–413. [Google Scholar] [CrossRef]
- Matsuda, H.; Morikawa, T.; Ueda, H.; Yokhikawa, M. Medicinal foodstuffs XXVII. Saponin constituents of Gotu Kola (2): Structures of new ursane- and oleanane-type triterpenene oligoglycerides, Centellasaponins B, C, and D, from Centella asiatica cultivated in Sri Lanka. Chem. Pharm. Bull. 2001, 49, 1368–1371. [Google Scholar] [CrossRef]
- Nath, S.; Buragohain, A.K. Establishment of callus and cell suspension cultures of Centella asiatica. Plant Biol. 2005, 49, 411–413. [Google Scholar] [CrossRef]
- Kim, Y.; Wyslouzil, B.E.; Weathers, P.J. Secondary metabolism of hairy root cultures in bioreactors. In Vitro Cell. Dev. Plant 2002, 38, 1–10. [Google Scholar]
- Bonfill, M.; Mangas, S.; Cusido, R.M.; Osuna, L.; Pinol, M.T.; Palazon, J. Identification of triterpenoid compounds of Centella asiatica. Plant Cell Tiss. Org. Cult. 2005, 104, 61–67. [Google Scholar]
- Bonfill, M.; Mangas, S.; Moyano, E.; Cusido, R.M.; Palazon, J. Production of centellosides and phytosterols in cell suspension cultures of Centella asiatica by thin-layer chromatography and mass spectrometry. Biomed. Chromatogr. 2011, 20, 151–153. [Google Scholar]
- Faria, J.M.S.; Nunes, I.S.; Figueiredo, A.C.; Pedro, L.G.; Trindale, H.; Barroso, J.G. Biotransformation of menthol and geraniol by hairy root cultures of Anetum greveolens: Effect on growth and volatile components. Biotechnol. Lett. 2009, 31, 897–903. [Google Scholar]
- Kiong, A.L.P.; Mahmood, M.; Fadzillah, N.M.; Daud, S.K. Effects of precursor supplementation on the production of triterpenes by Centella asiatica callus cultures. Pak. J. Biol. Sci. 2005, 8, 1160–1169. [Google Scholar] [CrossRef]
- Verpoorte, R.; Choi, Y.H.; Kim, H.K. NMR-based metabolomics at work in phytochemistry. Phytochem. Rev. 2007, 6, 3–14. [Google Scholar] [CrossRef]
- Kim, O.T.; Kim, S.H.; Ohyama, K.; Muranaka, T.; Choi, Y.E.; Lee, H.Y.; Kim, M.Y.; Hwang, B. Upregulation of phytosterol and triterpene biosynthesis in Centella asiatica hairy roots overexpressed ginseng farnesyl diphosphate synthase. Plant Cell Rep. 2010, 29, 403–411. [Google Scholar] [CrossRef]
- Cusidó, R.M.; Palazón, J.; Bonfill, M.; Navia-Osorio, A.; Morales, C.; Pińol, T. Improved paclitaxel and baccatin III production in cell suspension cultures of Taxus medias. Biotechnol. Prog. 2002, 18, 418–423. [Google Scholar] [CrossRef]
- Palazón, J.; Bonfill, R.M.C.M.; Mallol, A.; Moyano, E.; Morales, C.; Pinol, M.T. Elicitation of different Panax ginseng transformed root phenotypes for an improved ginsenoside production. Plant Physiol. Biochem. 2003, 41, 1019–1025. [Google Scholar] [CrossRef]
- Fritz, V.A.; Justen, V.L.; Bode, A.M.; Schuster, T.; Wang, M. Glucosinolate enhancement in cabbage induced by jasmonic acid application. HortScience 2010, 45, 1188–1191. [Google Scholar]
- James, J.T.; Meyer, R.; Dubery, I.A. Characterisation of two phenotypes of Centella asiatica in Southern Africa through the composition of four triterpenoids in callus, cell suspensions and leaves. Plant Cell Tiss. Org. Cult. 2008, 94, 91–99. [Google Scholar] [CrossRef]
- James, J.T.; Dubery, I.A. Identification and quantification of triterpenoid saponins and sapogenins in Centella asiatica (L.) Urban by densitometric TLC. J. Plan. Chromatogr. 2011, 24, 82–87. [Google Scholar] [CrossRef]
- Li, B.; Abliz, Z.; Tang, M.; Fu, G.; Yu, S. Rapid structural characterization of triterpenoid saponins in crude extract from Symplocos chinensis using liquid chromatography combined with electrospray ionization tandem mass spectrometry. J. Chromatogr. A 2005, 1101, 53–62. [Google Scholar]
- Hanisa, H.; Mohd Azmi, M.L.; Suhaila, M.; Somchit, M.N. Liquid chromatography-mass spectrometry-electrospray ionisation analysis of Centella asiatica I., Curcuma longa L. and Strobilanthes crispus L. methanol extracts. J. Med. Plant. Res. 2012, 6, 3908–3918. [Google Scholar]
- Goodacre, R.; Vaidyanathan, S.; Dunn, W.B.; Harrigan, G.G.; Kell, D.B. Metabolomics by numbers: Acquiring and understanding global metabolite data. Trends Biotechnol. 2004, 22, 245–252. [Google Scholar] [CrossRef]
- Trygg, J.; Holmes, E.; Lundstedt, T. Chemometrics in metabonomics. J. Proteome Res. 2007, 6, 469–479. [Google Scholar] [CrossRef]
- Boccard, J.; Grata, E.; Thiocone, A.; Gauvrit, J.Y.; Lanteri, P.; Carrupt, P.A.; Wolfender, J.L.; Rudaz, S. Multivariate data analysis of rapid LCTOF/MS experiments from Arabidopsis thaliana stressed by wounding. Chemometr. Intell. Lab. 2007, 86, 189–197. [Google Scholar]
- Tugizimana, F.; Piater, L.A.; Dubery, I.A. Plant metabolomics: A new frontier in phytochemical analysis. S. Afr. J. Sci. 2013, 109. in press. [Google Scholar]
- Trivedi, K.D.; Iles, K.R. The application of SIMCA P+ in shotgun metabolomics analysis of ZIC®HILIC-MS spectra of human urine - experience with the Shimadzu IT-TOF and profiling solutions data extraction software. J. Chromatogr. Sep. Tech. 2012, 3. [Google Scholar] [CrossRef]
- Dai, Y.; Li, Z.; Xue, L.; Dou, C.; Zhou, Y.; Zhang, L.; Qin, X. Metabolomics study on the anti-depression effect of xiaoyaosan on rat model of chronic unpredictable mild stress. J. Ethnopharmacol. 2010, 128, 482–489. [Google Scholar] [CrossRef]
- Chong, I.-G.; Jun, C.-H. Performance of some variable selection methods when multicollinearity is present. Chemometr. Intell. Lab. 2005, 78, 103–112. [Google Scholar] [CrossRef]
- Mehmood, T.; Liland, K.H.; Snipen, L.; Sæbø, S. A review of variable selection methods in partial least squares regression. Chemometr. Intell. Lab. 2012, 118, 62–69. [Google Scholar]
- Brown, M.; Dunn, W.B.; Dobson, P.; Patel, Y.; Winder, C.L.; Francis-McIntyre, S.; Begley, P.; Carroll, K.; Broadhurst, D.; Tseng, A.; et al. Mass spectrometry tools and metabolite-specific databases for molecular identification in metabolomics. Analyst 2009, 134, 1322–1332. [Google Scholar] [CrossRef]
- Taverna Workflow Management System Home Page. Available online: www.taverna.org.uk (accessed on 5 April 2013).
- Brown, M.; Wedge, D.C.; Goodacre, R.; Kell, D.B.; Baker, P.N.; Kenny, L.C.; Mamas, M.A.; Neyses, L.; Dunn, W.B. Automated workflows for accurate mass-based putative metabolite identification in LC/MS-derived metabolomic datasets. Bioinformatics 2011, 27, 1108–1112. [Google Scholar] [CrossRef]
- Chappell, J. The biochemistry and molecular biology of isoprenoid metabolism. Plant Physiol. 1995, 107, 1–6. [Google Scholar]
- Tholl, D. Terpene synthases and the regulation, diversity and biological roles of terpene metabolism. Curr. Opin. Plant Biol. 2006, 9, 297–304. [Google Scholar] [CrossRef]
- Ashour, M.; Wink, M.; Gershenzon, J. Biochemistry of Terpenoids: Monoterpenes, Sesquiterpenes and Diterpenes. In Annual Plant Reviews: Biochemistry of Plant Secondary Metabolism; Wink, M., Ed.; Wiley-Blackwell: Chichester, UK, 2010; pp. 258–303. [Google Scholar]
- Verpoorte, R. Secondary metabolism. In Metabolic Engineering of Plant Secondary Metabolism; Verpoorte, R., Alfermann, A.W., Eds.; Kluwer Academics Publishers: Dordrecht, The Netherlands, 2000; pp. 1–29. [Google Scholar]
- Chen, F.; Tholl, D.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef]
- Facchini, P.J.; Chappell, J. Gene family for an elicitor-induced sesquiterpene cyclase in tobacco. Proc. Natl. Acad. Sci. USA 1992, 89, 11088–11092. [Google Scholar] [CrossRef]
- Thaler, J.S.; Humphrey, P.T.; Whiteman, N.K. Evolution of jasmonate and salicylate signal crosstalk. Trends Plant Sci. 2012, 17, 260–270. [Google Scholar] [CrossRef]
- Rouillard-Guellec, F.; Robin, J.R.; Ratsimamanga, A.R.; Ratsimamanga, S.; Rasaoanaivo, R. Comparative study of Centella asiatica of Madagascar origin and Indian origin. Acta Bot. Gallica 1997, 144, 489–493. [Google Scholar] [CrossRef]
- Devkota, A.; Dall’Acqua, S.; Comai, S.; Innocenti, G. Centella asiatica (L.) Urban from Nepal: Quali-quantitative analysis of samples from several sites, and selection of high terpene containing populations for cultivation. Biochem. Syst. Ecol. 2010, 38, 12–22. [Google Scholar] [CrossRef]
- Byth, H.A.; Mchunu, B.I.; Dubery, I.A.; Bornman, L. Assessment of a simple, non-toxic Alamar blue cell survival assay to monitor tomato cell viability. Phytochem. Anal. 2001, 12, 340–346. [Google Scholar] [CrossRef]
- Jansen, J.J.; Smit, S.; Hoefsloot, H.C.J.; Smilde, A.K. The photographer and the greenhouse: How to analyse plant metabolomics data. Phytochem. Anal. 2010, 21, 48–60. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
James, J.T.; Tugizimana, F.; Steenkamp, P.A.; Dubery, I.A. Metabolomic Analysis of Methyl Jasmonate-Induced Triterpenoid Production in the Medicinal Herb Centella asiatica (L.) Urban. Molecules 2013, 18, 4267-4281. https://doi.org/10.3390/molecules18044267
James JT, Tugizimana F, Steenkamp PA, Dubery IA. Metabolomic Analysis of Methyl Jasmonate-Induced Triterpenoid Production in the Medicinal Herb Centella asiatica (L.) Urban. Molecules. 2013; 18(4):4267-4281. https://doi.org/10.3390/molecules18044267
Chicago/Turabian StyleJames, Jacinda T., Fidele Tugizimana, Paul A. Steenkamp, and Ian A. Dubery. 2013. "Metabolomic Analysis of Methyl Jasmonate-Induced Triterpenoid Production in the Medicinal Herb Centella asiatica (L.) Urban" Molecules 18, no. 4: 4267-4281. https://doi.org/10.3390/molecules18044267