Protective Mechanisms of Guanosine from Solanum lycopersicum on Agonist-Induced Platelet Activation: Role of sCD40L



Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
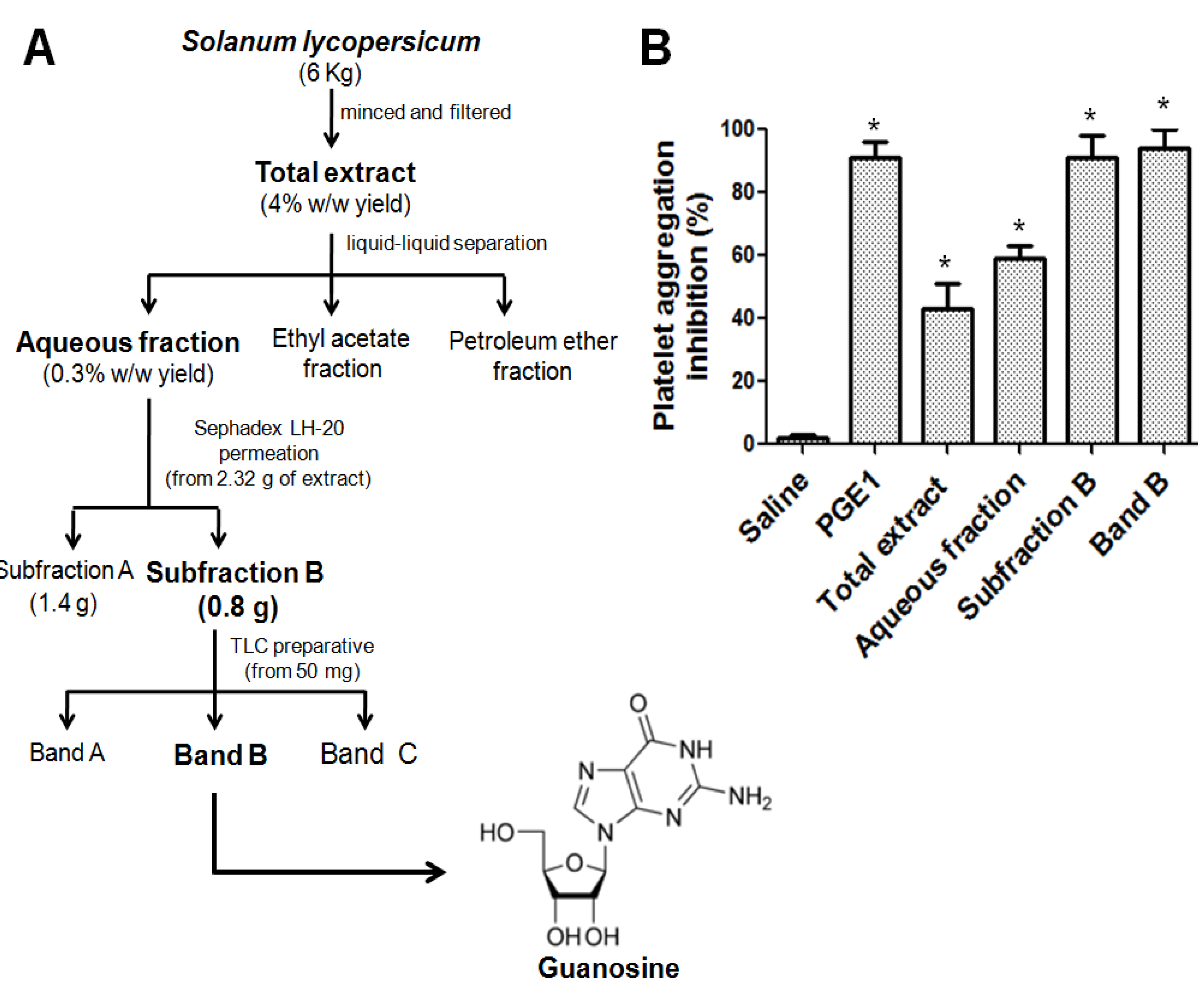
2.1. Bioassay-Guided Isolation of Antiplatelet Compound

2.2. Identification of the Antiplatelet Compound
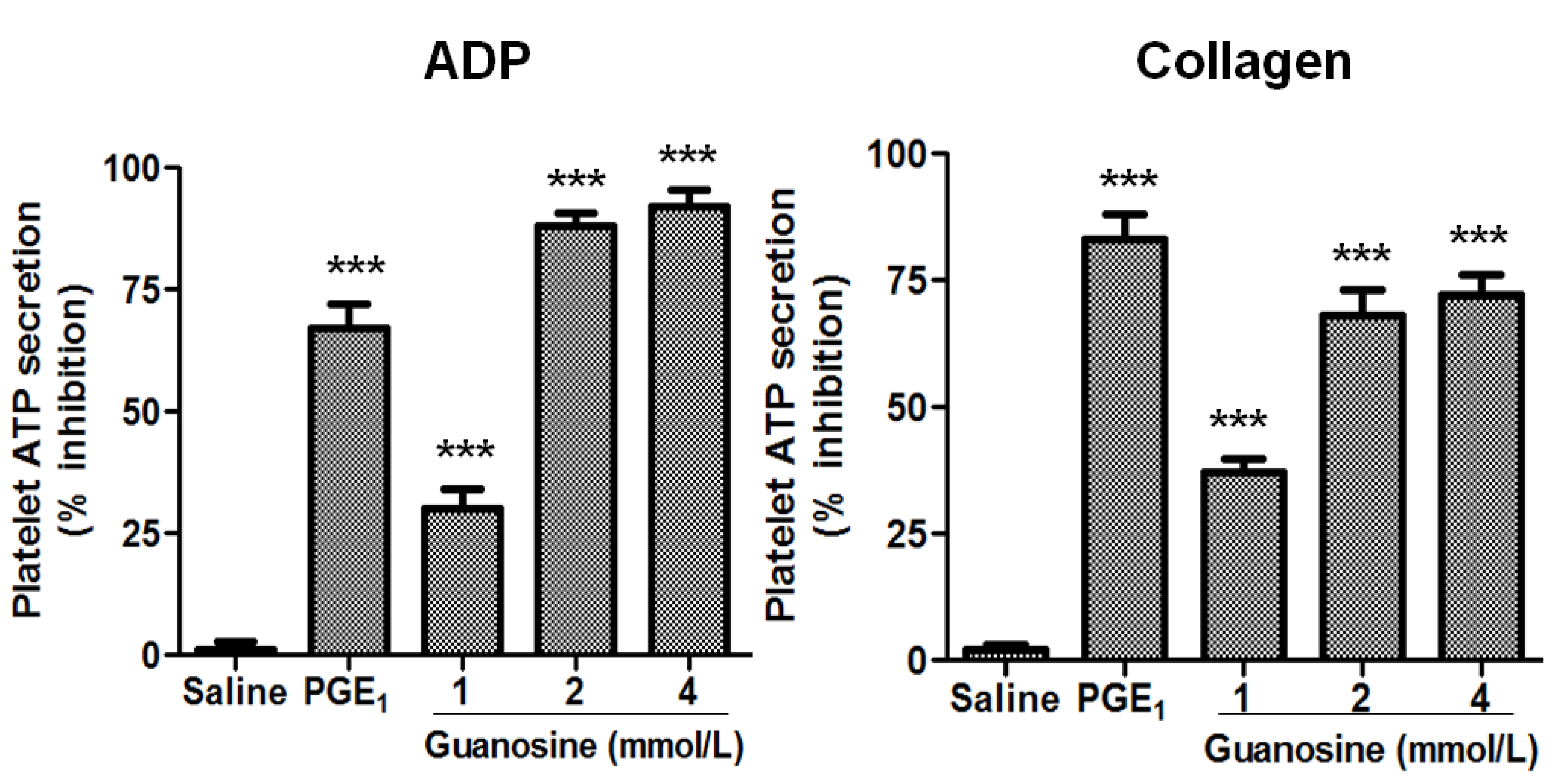
2.3. Effects of Guanosine on Platelet Function


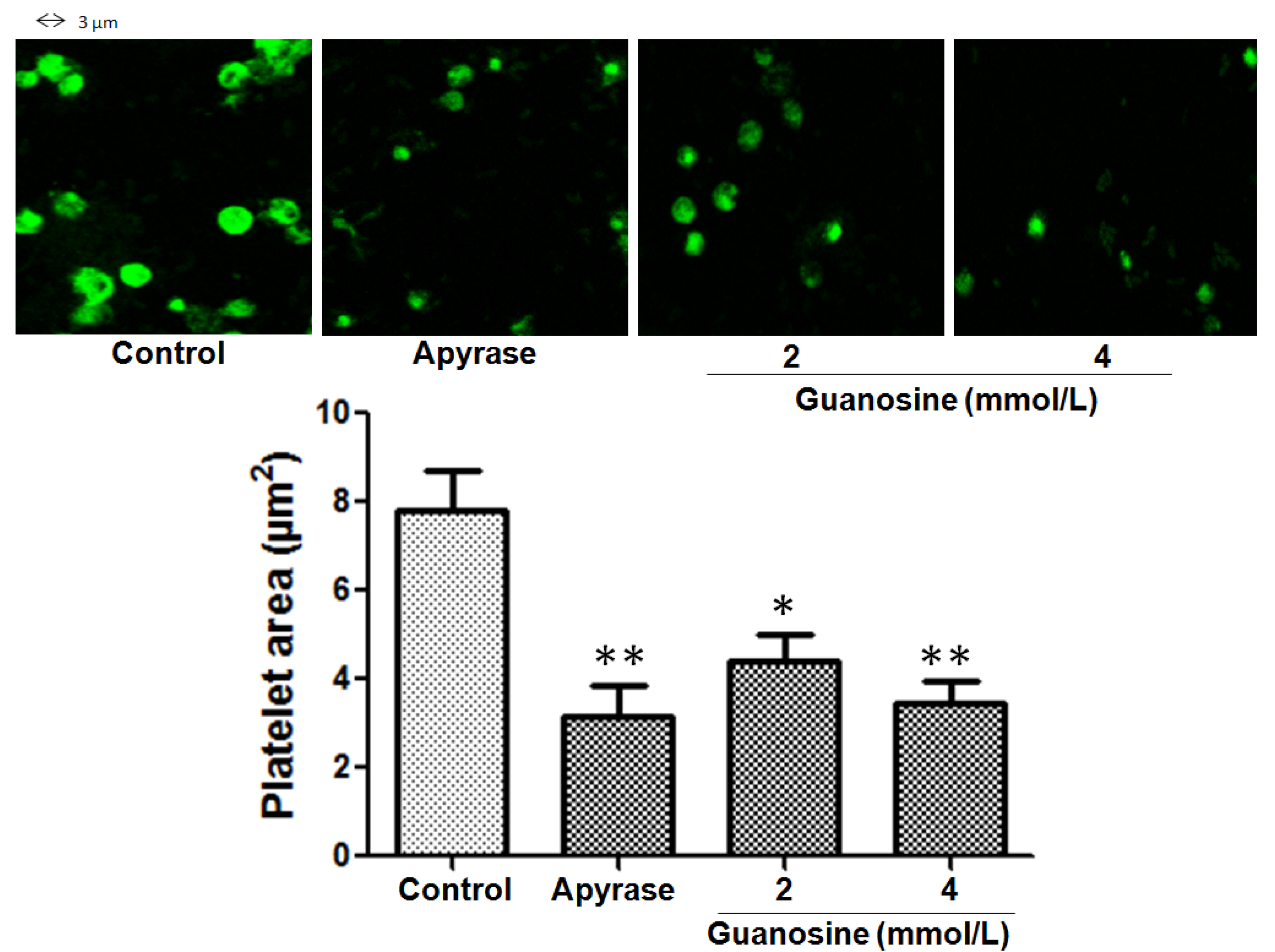
2.4. Inhibitory Effect of Guanosine on Platelet Spreading on Immobilized Collagen

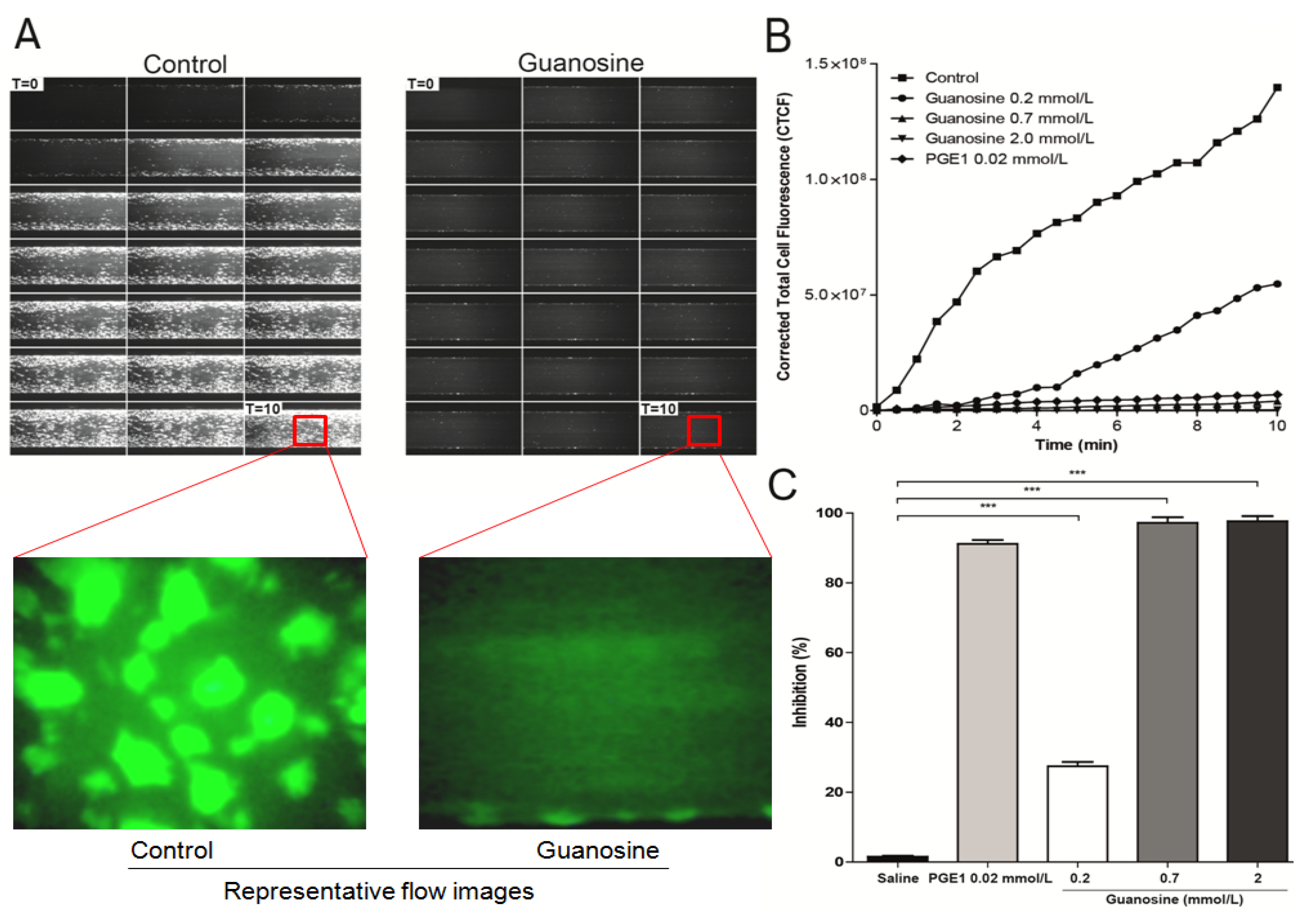
2.5. Guanosine Reduces Platelet Adhesion and Aggregation under Flow Conditions

2.6. Inhibitory Effect of Guanosine on Levels of sCD40L


3. Experimental
3.1. Chemicals and Reagents
3.2. Processing Material
3.3. Bioassay-Guided Isolation of Bioactive Compound
3.4. Chemical Identification of Antiplatelet Compound
3.4.1. Spectral Scanning
3.4.2. NMR Analysis
3.4.3. HPLC Analysis
3.5. Preparation of Human Platelet Suspensions
3.6. Measurement of Platelet Secretion
3.7. Platelet Spreading Assay
3.8. Measurement of Platelet Aggregation
3.9. Analysis of Platelet Adhesion and Aggregation under Controlled Flow
3.10. Measurement of sCD40L Levels
3.11. Statistical Analysis
4. Conclusion
Acknowledgments
Conflicts of Interest
References
- World Health Organization, WHO Publishes Definitive Atlas on Global Heart Disease and Stroke Epidemic (The atlas of heart disease and stroke); WHO: Geneva, Switzerland, 2004.
- Roger, V.L.; Go, A.S.; Lloyd-Jones, D.M.; Adams, R.J.; Berry, J.D.; Brown, T.M.; Carnethon, M.R.; Dai, S.; de Simone, G.; Ford, E.S.; et al. Heart disease and stroke statistics-2011 update: A report from the American Heart Association. Circulation 2011, 123, 18–209. [Google Scholar] [CrossRef]
- Reddy, K.S.; Yusuf, S. Emerging epidemic of cardiovascular disease in developing countries. Circulation 1998, 97, 596–601. [Google Scholar] [CrossRef]
- Jackson, C.F.; Wenger, N.K. Cardiovascular disease in the elderly. Rev. Esp. Cardiol. 2011, 64, 697–712. [Google Scholar] [CrossRef]
- Naghavi, M.; Libby, P.; Falk, E.; Casscells, S.W.; Litovsky, S.; Rumberger, J.; Badimon, J.J.; Stefanadis, C.; Moreno, P.; Gerard, P.; et al. From vulnerable plaque to vulnerable patient: a call for new definitions and risk assessment strategies: Part I. Circulation 2003, 108, 1664–1672. [Google Scholar] [CrossRef]
- Marenberg, M.E.; Risch, N.; Berkman, L.F.; Floderus, B.; de Faire, U. Genetic susceptibility to death from coronary heart disease in a study of twins. N. Engl. J. Med. 1994, 330, 1041–1046. [Google Scholar] [CrossRef]
- Palomo, I.; Toro, C.; Alarcon, M. The role of platelets in the pathophysiology of atherosclerosis (Review). Mol. Med. Report 2008, 1, 179–184. [Google Scholar]
- Davi, G.; Patrono, C. Platelet activation and atherothrombosis. N. Engl. J. Med. 2007, 357, 2482–2494. [Google Scholar] [CrossRef]
- Fuentes, Q.E.; Fuentes, Q.F.; Andres, V.; Pello, O.M.; de Mora, J.F.; Palomo, G.I. Role of plateletsas mediators that link inflammation and thrombosis in atherosclerosis. Platelets 2013, 24, 255–262. [Google Scholar] [CrossRef]
- Wagner, D.D.; Burger, P.C. Platelets in inflammation and thrombosis. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 2131–2137. [Google Scholar] [CrossRef]
- Missiou, A.; Wolf, D.; Platzer, I.; Ernst, S.; Walter, C.; Rudolf, P.; Zirlik, K.; Kostlin, N.; Willecke, F.K.; Munkel, C.; et al. CD40L induces inflammation and adipogenesis in adipose cells--a potential link between metabolic and cardiovascular disease. Thromb. Haemost. 2010, 103, 788–796. [Google Scholar] [CrossRef]
- Schonbeck, U.; Sukhova, G.K.; Shimizu, K.; Mach, F.; Libby, P. Inhibition of CD40 signaling limits evolution of established atherosclerosis in mice. Proc. Natl. Acad. Sci. USA 2000, 97, 7458–7463. [Google Scholar] [CrossRef]
- Zoungas, S.; McGrath, B.P.; Branley, P.; Kerr, P.G.; Muske, C.; Wolfe, R.; Atkins, R.C.; Nicholls, K.; Fraenkel, M.; Hutchison, B.G.; et al. Cardiovascular morbidity and mortality in the Atherosclerosis and Folic Acid Supplementation Trial (ASFAST) in chronic renal failure: A multicenter, randomized, controlled trial. J. Am. Coll. Cardiol. 2006, 47, 1108–1116. [Google Scholar] [CrossRef]
- Estruch, R.; Martinez-Gonzalez, M.A.; Corella, D.; Salas-Salvado, J.; Ruiz-Gutierrez, V.; Covas, M.I.; Fiol, M.; Gomez-Gracia, E.; Lopez-Sabater, M.C.; Vinyoles, E.; et al. Effects of a Mediterranean-style diet on cardiovascular risk factors: a randomized trial. Ann. Intern. Med. 2006, 145, 1–11. [Google Scholar] [CrossRef]
- Pearson, T.A.; Blair, S.N.; Daniels, S.R.; Eckel, R.H.; Fair, J.M.; Fortmann, S.P.; Franklin, B.A.; Goldstein, L.B.; Greenland, P.; Grundy, S.M.; et al. AHA guidelines for primary prevention of cardiovascular disease and stroke: 2002 Update: consensus panel guide to comprehensive risk reduction for adult patients without coronary or other atherosclerotic vascular diseases. american heart association science advisory and coordinating committee. Circulation 2002, 106, 388–391. [Google Scholar] [CrossRef]
- Palomo, I.; Leiva, E.; Vásquez, M. Dieta Mediterranea: Prevención De Las Enfermedades Cardiovasculares (in Spanish language); Universidad de Talca Press: Talca, Chile, 2007. [Google Scholar]
- Koleckar, V.; Brojerova, E.; Rehakova, Z.; Kubikova, K.; Cervenka, F.; Kuca, K.; Jun, D.; Hronek, M.; Opletalova, V.; Opletal, L. In vitro antiplatelet activity of flavonoids from Leuzea carthamoides. Drug Chem. Toxicol. 2008, 31, 27–35. [Google Scholar] [CrossRef]
- Khan, A.N.; Fatima, I.; Khaliq, U.A.; Malik, A.; Miana, G.A.; Qureshi, Z.U.; Rasheed, H. Potent anti-platelet constituents from Centaurea iberica. Molecules 2011, 16, 2053–2064. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef]
- Fuentes, E.; Astudillo, L.A.; Gutierrez, M.I.; Contreras, S.O.; Bustamante, L.O.; Rubio, P.I.; Moore-Carrasco, R.; Alarcon, M.A.; Fuentes, J.A.; Gonzalez, D.E.; et al. Fractions of aqueous and methanolic extracts from tomato (Solanum lycopersicum L.) present platelet antiaggregant activity. Blood Coagul. Fibrinolysis 2012, 23, 109–117. [Google Scholar] [CrossRef]
- Fuentes, E.; Castro, R.; Astudillo, L.; Carrasco, G.; Alarcon, M.; Gutierrez, M.; Palomo, I. Bioassay-Guided isolation and HPLC determination of bioactive compound that relate to the antiplatelet activity (Adhesion, Secretion, and Aggregation) from Solanum lycopersicu. Evid. Based Complement Alternat. Med. 2012, 2012, 147031. [Google Scholar]
- Palomo, I.; Fuentes, E.; Padró, T.; Badimon, L. Platelets and atherogenesis: Platelet antiaggregating activity and endothelial protection from tomatoes (Solanum lycopersicum L.). Exp. Ther. Med. 2012, 3, 577–584. [Google Scholar]
- Agarwal, A.; Shen, H.; Agarwal, S.; Rao, A.V. Lycopene content of tomato products: its stability, bioavailability and in vivo antioxidant properties. J. Med. Food 2001, 4, 9–15. [Google Scholar] [CrossRef]
- Barrett, N.E.; Holbrook, L.; Jones, S.; Kaiser, W.J.; Moraes, L.A.; Rana, R.; Sage, T.; Stanley, R.G.; Tucker, K.L.; Wright, B.; Gibbins, J.M. Future innovations in anti-platelet therapies. Br. J. Pharmacol. 2008, 154, 918–939. [Google Scholar] [CrossRef]
- Connor, W.E. Importance of n-3 fatty acids in health and disease. Am. J. Clin. Nutr. 2000, 71, 171–175. [Google Scholar]
- Fuentes, E.; Carle, R.; Astudillo, L.; Guzmán, L.; Gutiérrez, M.; Carrasco, G.; Palomo, I. Antioxidant and antiplatelet activities in extracts from green and fully ripe tomato fruits (Solanum lycopersicum) and pomace from industrial tomato processing. Evid.-Based Complement. Altern. Med. 2013, 2013, 1–9. [Google Scholar]
- Yang, D.; Chen, H.; Koupenova, M.; Carroll, S.H.; Eliades, A.; Freedman, J.E.; Toselli, P.; Ravid, K. A new role for the A2b adenosine receptor in regulating platelet function. J. Thromb. Haemost. 2010, 8, 817–827. [Google Scholar] [CrossRef]
- Loyau, S.; Dumont, B.; Ollivier, V.; Boulaftali, Y.; Feldman, L.; Ajzenberg, N.; Jandrot-Perrus, M. Platelet glycoprotein VI dimerization, an active process inducing receptor competence, is an indicator of platelet reactivity. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 778–785. [Google Scholar] [CrossRef]
- Inoue, O.; Suzuki-Inoue, K.; Dean, W.L.; Frampton, J.; Watson, S.P. Integrin alpha2beta1 mediates outside-in regulation of platelet spreading on collagen through activation of Src kinases and PLCgamma2. J. Cell Biol. 2003, 160, 769–780. [Google Scholar] [CrossRef]
- Ruggeri, Z.M.; Mendolicchio, G.L. Adhesion mechanisms in platelet function. Circ. Res. 2007, 100, 1673–1685. [Google Scholar] [CrossRef]
- Aukrust, P.; Muller, F.; Ueland, T.; Berget, T.; Aaser, E.; Brunsvig, A.; Solum, N.O.; Forfang, K.; Froland, S.S.; Gullestad, L. Enhanced levels of soluble and membrane-bound CD40 ligand in patients with unstable angina. Possible reflection of T lymphocyte and platelet involvement in the pathogenesis of acute coronary syndromes. Circulation 1999, 100, 614–620. [Google Scholar] [CrossRef]
- Nurden, A.T. Platelets, inflammation and tissue regeneration. Thromb. Haemost. 2011, 105, 13–33. [Google Scholar] [CrossRef]
- Andre, P.; Nannizzi-Alaimo, L.; Prasad, S.K.; Phillips, D.R. Platelet-derived CD40L: The switch-hitting player of cardiovascular disease. Circulation 2002, 106, 896–899. [Google Scholar] [CrossRef]
- Setianto, B.Y.; Hartopo, A.B.; Gharini, P.P.; Anggrahini, D.W.; Irawan, B. Circulating soluble CD40 ligand mediates the interaction between neutrophils and platelets in acute coronary syndrome. Heart Vessels 2010, 25, 282–287. [Google Scholar] [CrossRef]
- Lievens, D.; Zernecke, A.; Seijkens, T.; Soehnlein, O.; Beckers, L.; Munnix, I.C.; Wijnands, E.; Goossens, P.; van Kruchten, R.; Thevissen, L.; et al. Platelet CD40L mediates thrombotic and inflammatory processes in atherosclerosis. Blood 2010, 116, 4317–4327. [Google Scholar] [CrossRef]
- Lutgens, E.; Cleutjens, K.B.; Heeneman, S.; Koteliansky, V.E.; Burkly, L.C.; Daemen, M.J. Both early and delayed anti-CD40L antibody treatment induces a stable plaque phenotype. Proc. Natl. Acad. Sci. USA 2000, 97, 7464–7469. [Google Scholar]
- Boylan, B.; Gao, C.; Rathore, V.; Gill, J.C.; Newman, D.K.; Newman, P.J. Identification of FcgammaRIIa as the ITAM-bearing receptor mediating alphaIIbbeta3 outside-in integrin signaling in human platelets. Blood 2008, 112, 2780–2786. [Google Scholar] [CrossRef]
- Born, G.V.; Cross, M.J. The aggregation of blood platelets. J. Physiol. 1963, 168, 178–195. [Google Scholar]
- Conant, C.G.; Schwartz, M.A.; Nevill, T.; Ionescu-Zanetti, C. Platelet adhesion and aggregation under flow using microfluidic flow cells. J. Vis. Exp. 2009, 32, 1644. [Google Scholar]
- Worth, R.G.; Chien, C.D.; Chien, P.; Reilly, M.P.; McKenzie, S.E.; Schreiber, A.D. Platelet FcgammaRIIA binds and internalizes IgG-containing complexes. Exp. Hematol. 2006, 34, 1490–1495. [Google Scholar] [CrossRef]
- Antczak, A.J.; Singh, N.; Gay, S.R.; Worth, R.G. IgG-complex stimulated platelets: A source of sCD40L and RANTES in initiation of inflammatory cascade. Cell Immunol. 2010, 263, 129–133. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the guanosine is available from the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Fuentes, E.; Alarcón, M.; Astudillo, L.; Valenzuela, C.; Gutiérrez, M.; Palomo, I. Protective Mechanisms of Guanosine from Solanum lycopersicum on Agonist-Induced Platelet Activation: Role of sCD40L. Molecules 2013, 18, 8120-8135. https://doi.org/10.3390/molecules18078120
Fuentes E, Alarcón M, Astudillo L, Valenzuela C, Gutiérrez M, Palomo I. Protective Mechanisms of Guanosine from Solanum lycopersicum on Agonist-Induced Platelet Activation: Role of sCD40L. Molecules. 2013; 18(7):8120-8135. https://doi.org/10.3390/molecules18078120
Chicago/Turabian StyleFuentes, Eduardo, Marcelo Alarcón, Luis Astudillo, Claudio Valenzuela, Margarita Gutiérrez, and Iván Palomo. 2013. "Protective Mechanisms of Guanosine from Solanum lycopersicum on Agonist-Induced Platelet Activation: Role of sCD40L" Molecules 18, no. 7: 8120-8135. https://doi.org/10.3390/molecules18078120
APA StyleFuentes, E., Alarcón, M., Astudillo, L., Valenzuela, C., Gutiérrez, M., & Palomo, I. (2013). Protective Mechanisms of Guanosine from Solanum lycopersicum on Agonist-Induced Platelet Activation: Role of sCD40L. Molecules, 18(7), 8120-8135. https://doi.org/10.3390/molecules18078120