Brefeldin A Effectively Inhibits Cancer Stem Cell-Like Properties and MMP-9 Activity in Human Colorectal Cancer Colo 205 Cells

 , and
, and 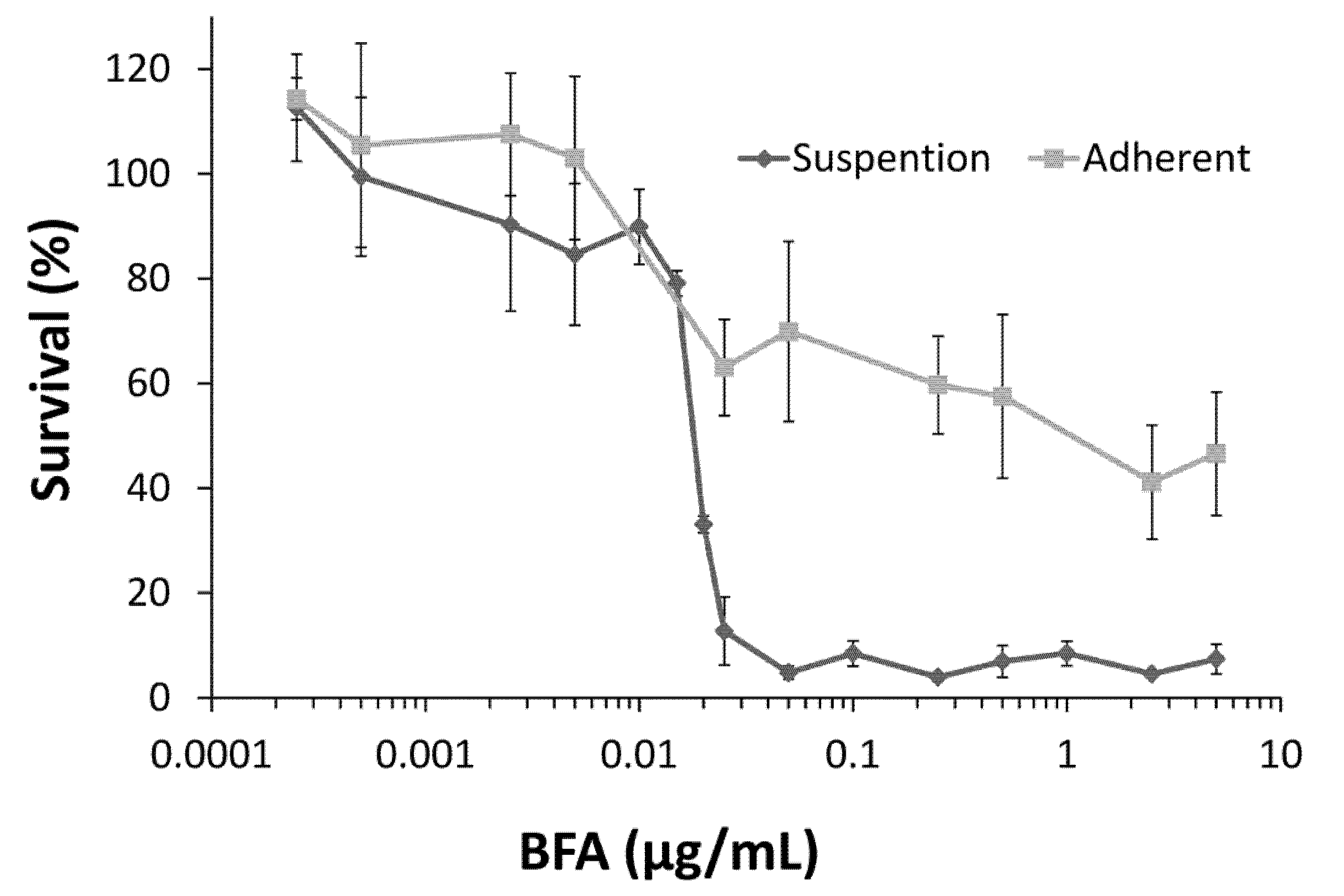{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Colo 205 Suspension Cells Were Sensitive to the Cytotoxic Effect of BFA
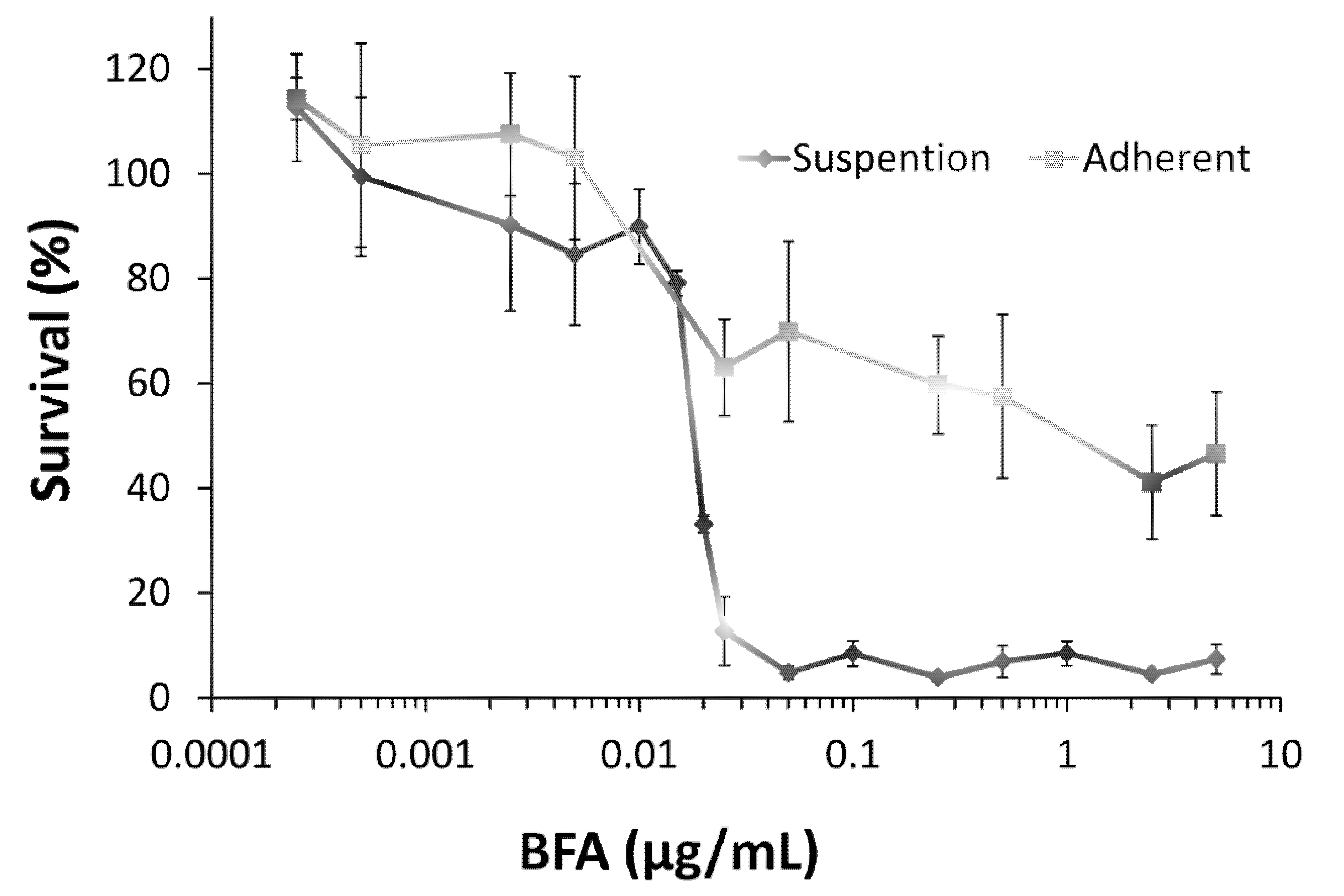
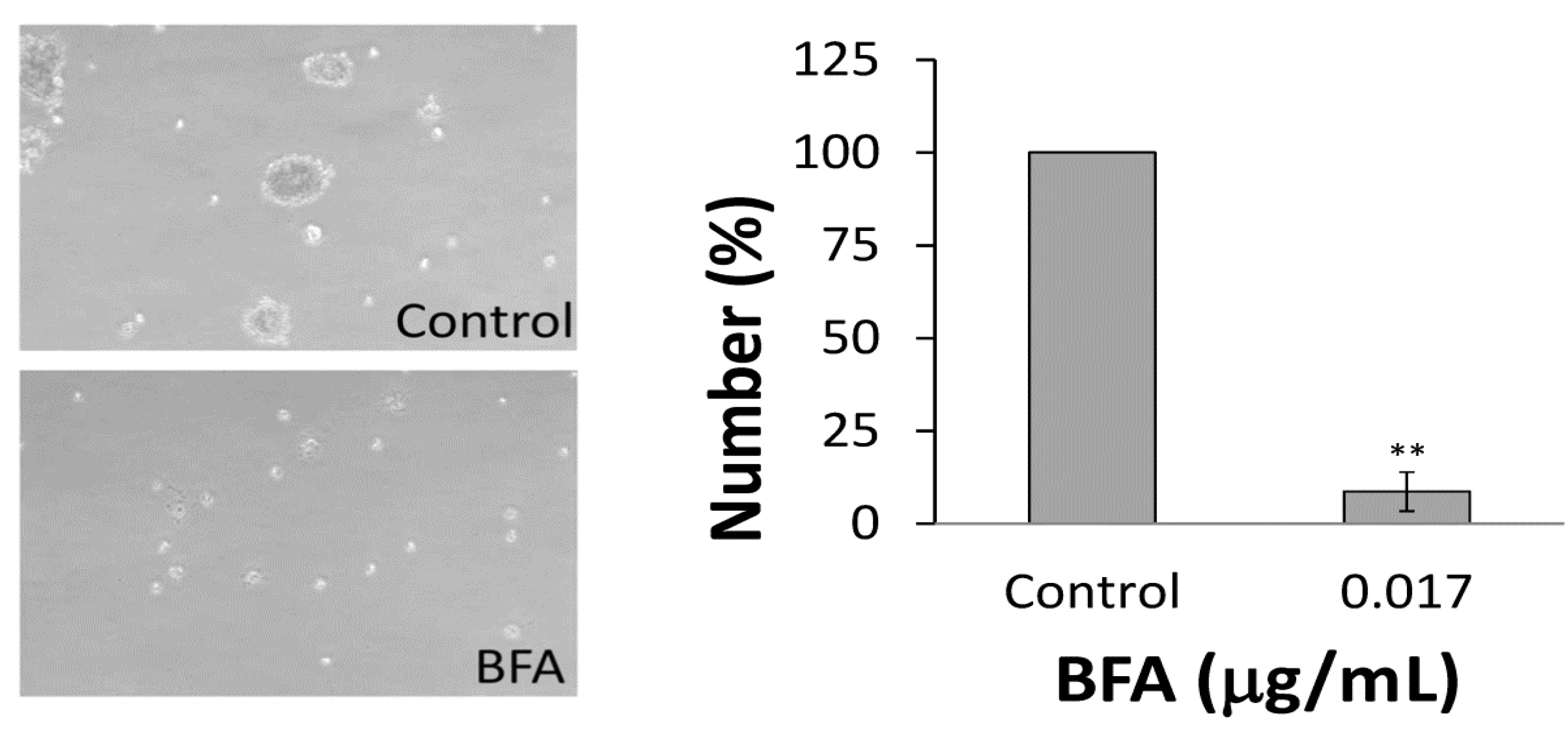
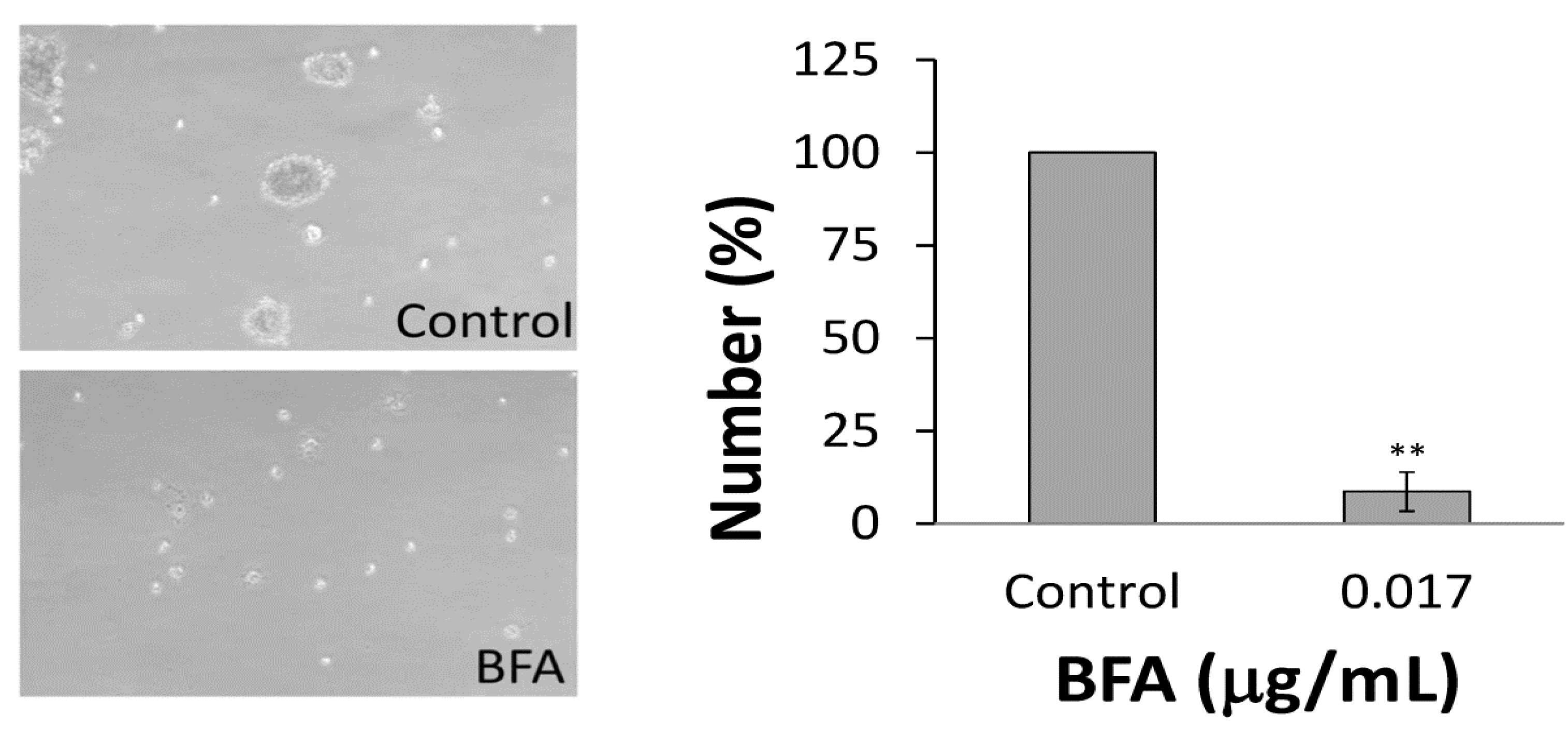
2.2. BFA Reduced the Clonogenicity of Colo 205 CSCs

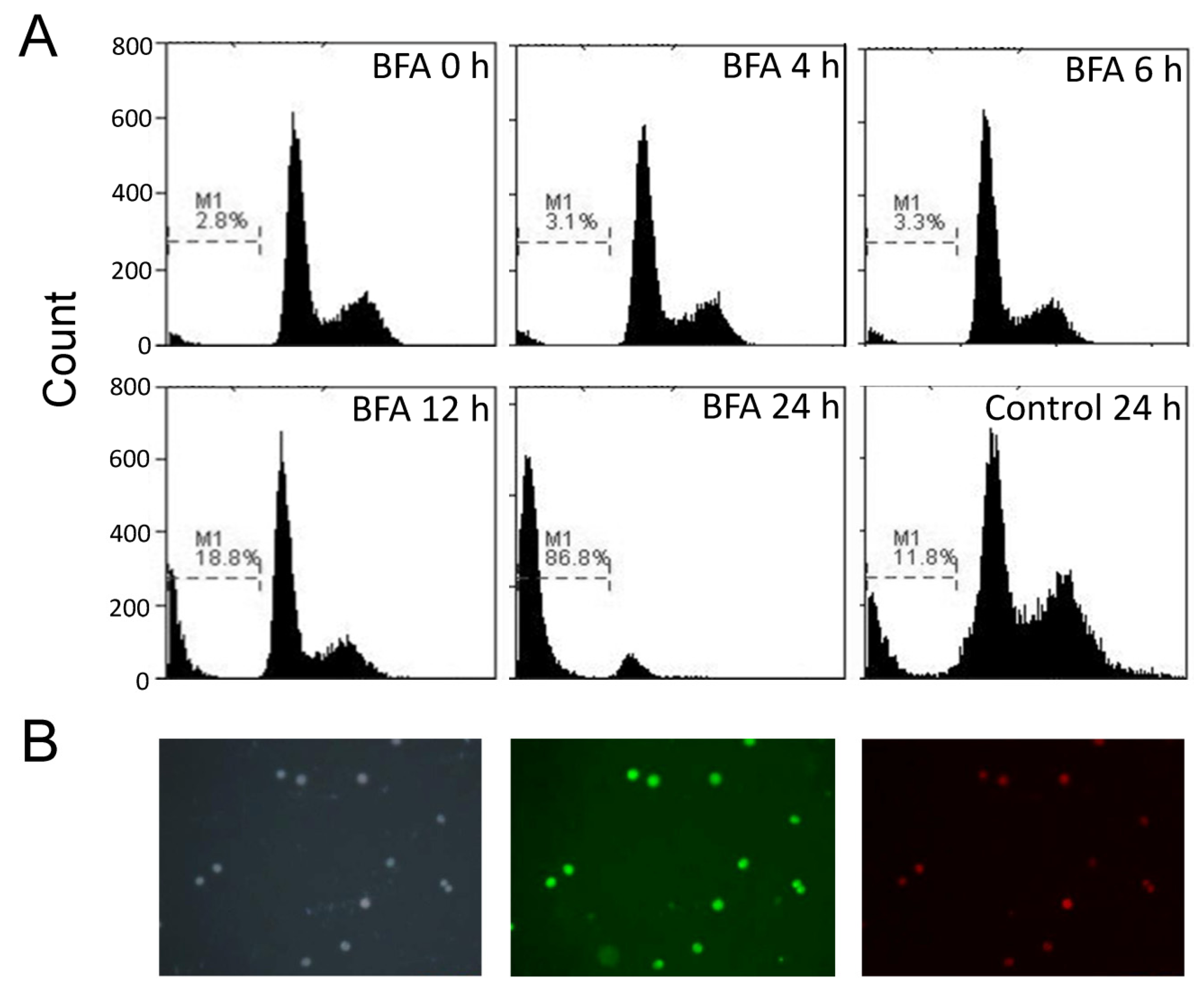
2.3. BFA Induced Apoptosis of Suspension Colo 205 cells
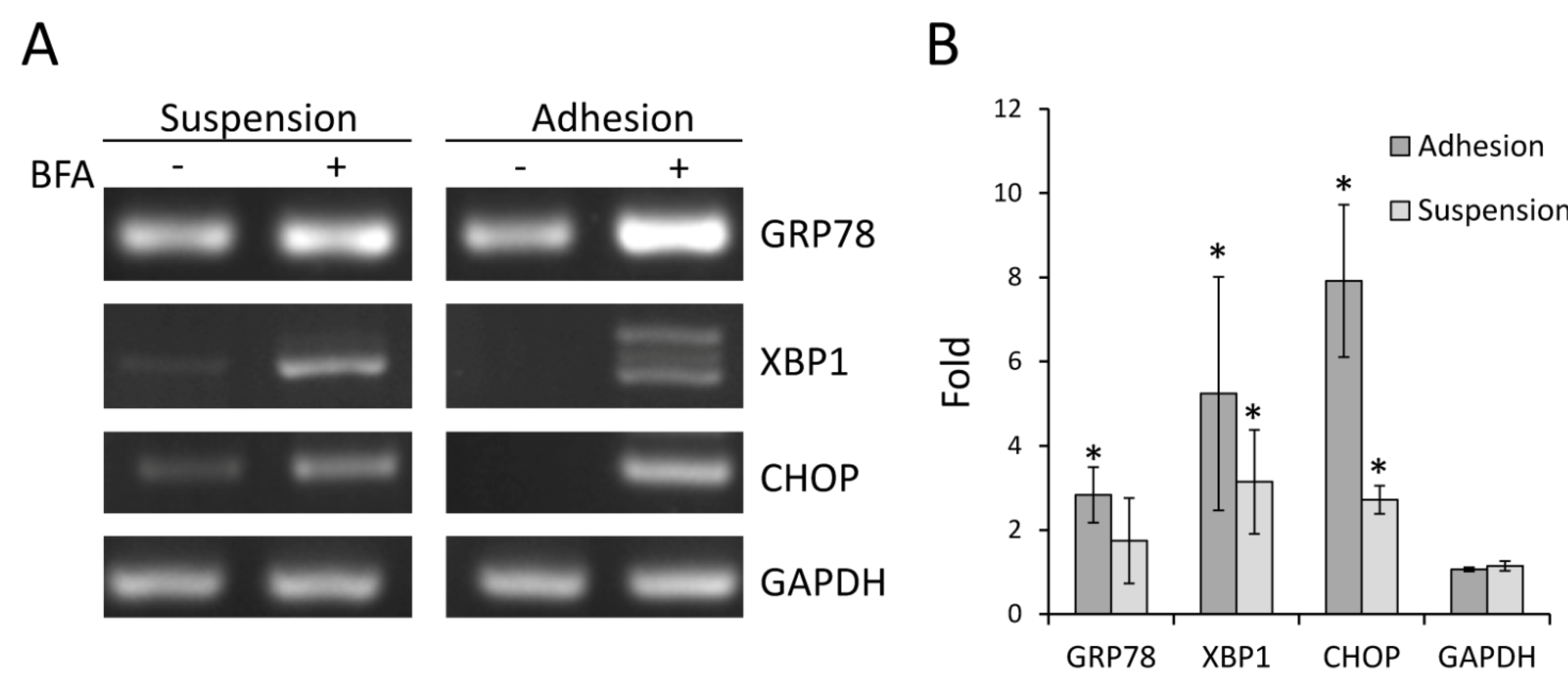
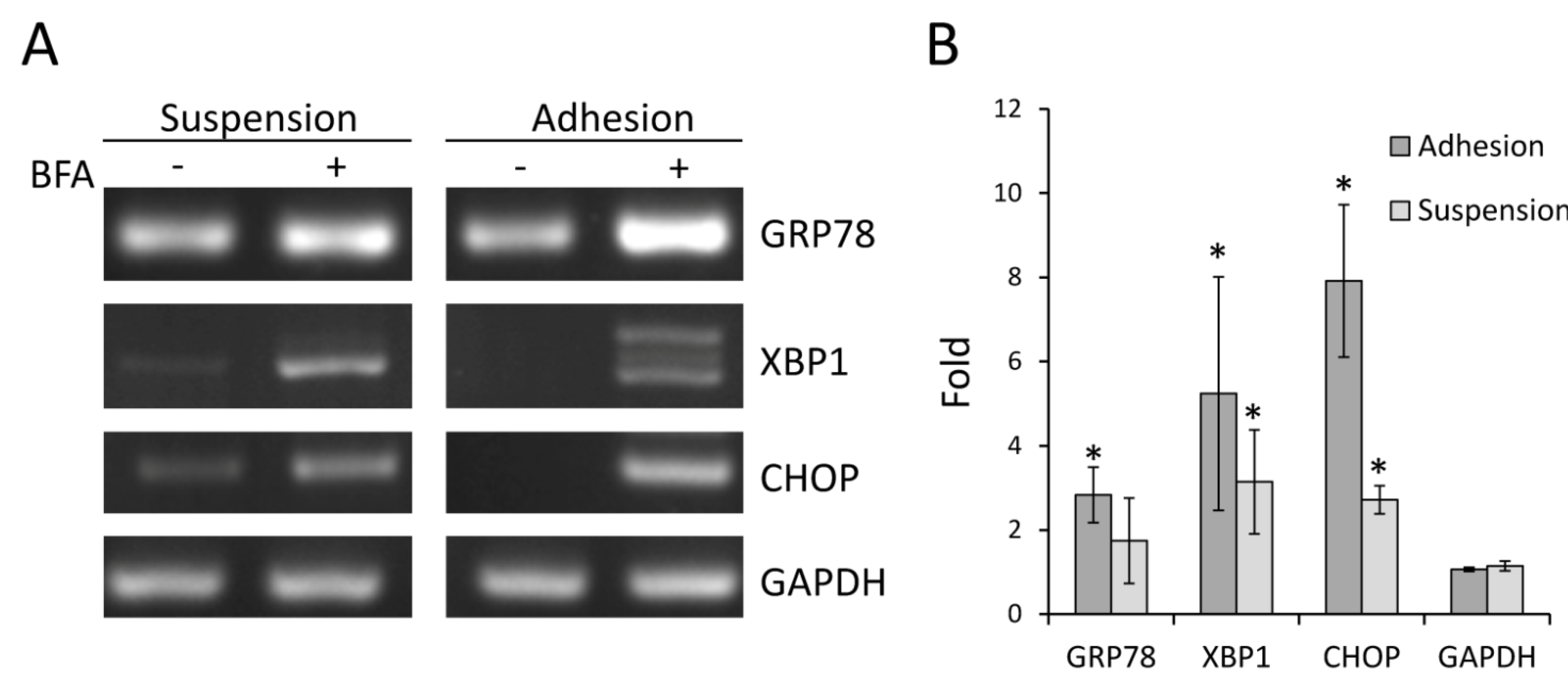
2.4. BFA Induced ER Stress Response in Suspension Colo 205 Cells at Nanogram per Milliliter Range
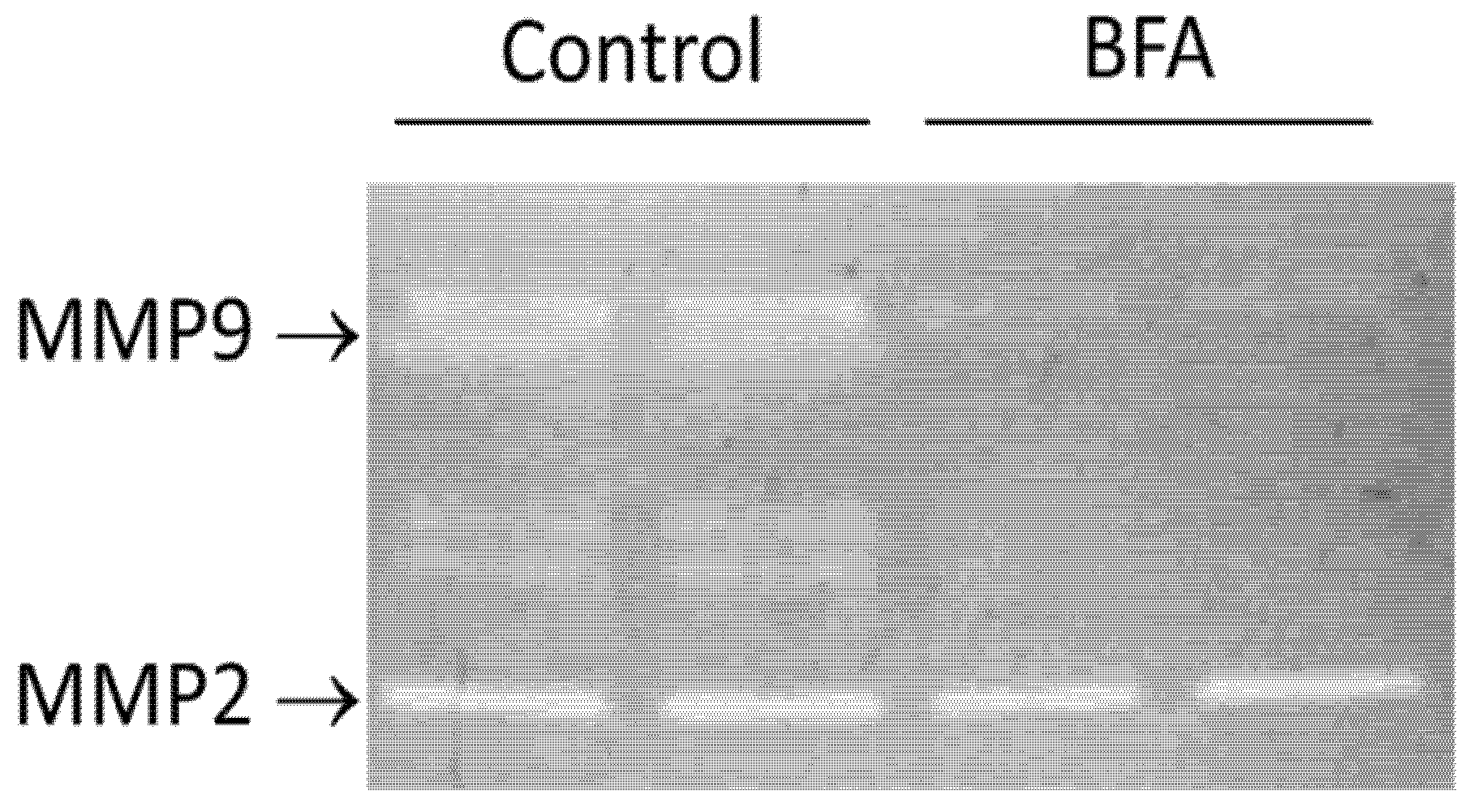
2.5. BFA Reduced the Activity of MMP9

2.6. Discussion
3. Experimental
3.1. Cell Culture
3.2. Cell Survival Assay
3.3. Soft Agar Colony Formation Assay
3.4. Determination of Sub-G1 Population and Annexin V/PI Staining
3.5. Reverse-Transcription Polymerase Chain Reaction (RT-PCR)
3.6. Gelatin Zymography
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA-Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef]
- Kriza, C.; Emmert, M.; Wahlster, P.; Niederlander, C.; Kolominsky-Rabas, P. Cost of Illness in Colorectal Cancer: An International Review. Pharmaco Economics 2013, 31, 577–588. [Google Scholar] [CrossRef]
- Chang, H.F.; Yang, L.L. Gamma-mangostin, a micronutrient of mangosteen fruit, Induces apoptosis in human colon cancer cells. Molecules 2012, 17, 8010–8021. [Google Scholar] [CrossRef]
- Kim, R.P.; Bihud, V.; Bin Mohamad, K.; Leong, K.H.; Bin Mohamad, J.; Bin Ahmad, F.; Hazni, H.; Kasim, N.; Halim, S.N.; Awang, K. Cytotoxic and antioxidant compounds from the stem bark of Goniothalamus tapisoides Mat Salleh. Molecules 2012, 18, 128–139. [Google Scholar] [CrossRef]
- Reya, T.; Morrison, S.J.; Clarke, M.F.; Weissman, I.L. Stem cells, Cancer, and cancer stem cells. Nature 2001, 414, 105–111. [Google Scholar] [CrossRef]
- Yamashita, T.; Wang, X.W. Cancer stem cells in the development of liver cancer. J. Clin. Invest. 2013, 123, 1911–1918. [Google Scholar] [CrossRef]
- Peng, C.Y.; Fong, P.C.; Yu, C.C.; Tsai, W.C.; Tzeng, Y.M.; Chang, W.W. Methyl Antcinate A suppresses the population of cancer stem-like cells in MCF7 human breast cancer cell line. Molecules 2013, 18, 2539–2348. [Google Scholar] [CrossRef]
- Dhawan, P.; Ahmad, R.; Srivastava, A.S.; Singh, A.B. Cancer stem cells and colorectal cancer: An overview. Curr. Top. Med. Chem. 2011, 11, 1592–1598. [Google Scholar] [CrossRef]
- Olszewski, U.; Liedauer, R.; Ausch, C.; Thalhammer, T.; Hamilton, G. Overexpression of CYP3A4 in a COLO 205 Colon Cancer Stem Cell Model in vitro. Cancers 2011, 3, 1467–1479. [Google Scholar] [CrossRef]
- Chen, S.F.; Chang, Y.C.; Nieh, S.; Liu, C.L.; Yang, C.Y.; Lin, Y.S. Nonadhesive culture system as a model of rapid sphere formation with cancer stem cell properties. PLoS One 2012, 7, e31864. [Google Scholar]
- Kwatra, D.; Subramaniam, D.; Ramamoorthy, P.; Standing, D.; Moran, E.; Velayutham, R.; Mitra, A.; Umar, S.; Anant, S. Methanolic extracts of bitter melon inhibit colon cancer stem cells by affecting energy homeostasis and autophagy. J. Evid. Based Complementary Altern. Med. 2013, 2013, 702869. [Google Scholar]
- Gupta, P.B.; Onder, T.T.; Jiang, G.; Tao, K.; Kuperwasser, C.; Weinberg, R.A.; Lander, E.S. Identification of selective inhibitors of cancer stem cells by high-throughput screening. Cell 2009, 138, 645–659. [Google Scholar] [CrossRef]
- Tamura, G.; Ando, K.; Suzuki, S.; Takatsuki, A.; Arima, K. Antiviral activity of brefeldin A and verrucarin A. J. Antibiot. 1968, 21, 160–161. [Google Scholar] [CrossRef]
- Chardin, P.; McCormick, F. Brefeldin A: the advantage of being uncompetitive. Cell 1999, 97, 153–155. [Google Scholar] [CrossRef]
- Natsume, Y.; Ito, S.; Satsu, H.; Shimizu, M. Protective effect of quercetin on ER stress caused by calcium dynamics dysregulation in intestinal epithelial cells. Toxicology 2009, 258, 164–175. [Google Scholar] [CrossRef]
- Lee, M.S.; Chao, J.; Yen, J.C.; Lin, L.W.; Tsai, F.S.; Hsieh, M.T.; Peng, W.H.; Cheng, H.Y. Schizandrin protects primary rat cortical cell cultures from glutamate-induced apoptosis by inhibiting activation of the MAPK family and the mitochondria dependent pathway. Molecules 2012, 18, 354–372. [Google Scholar] [CrossRef]
- Avila, M.F.; Cabezas, R.; Torrente, D.; Gonzalez, J.; Morales, L.; Alvarez, L.; Capani, F.; Barreto, G.E. Novel interactions of GRP78: UPR and estrogen responses in the brain. Cell Biol. Int. 2013, 37, 521–532. [Google Scholar] [CrossRef]
- Iwakoshi, N.N.; Lee, A.H.; Vallabhajosyula, P.; Otipoby, K.L.; Rajewsky, K.; Glimcher, L.H. Plasma cell differentiation and the unfolded protein response intersect at the transcription factor XBP-1. Nat. Immunol. 2003, 4, 321–329. [Google Scholar]
- Rajamahanty, S.; Alonzo, C.; Aynehchi, S.; Choudhury, M.; Konno, S. Growth inhibition of androgen-responsive prostate cancer cells with brefeldin A targeting cell cycle and androgen receptor. Int. J. Biomed. Sci. 2010, 17, 5. [Google Scholar] [CrossRef]
- Lin, L.; Fuchs, J.; Li, C.; Olson, V.; Bekaii-Saab, T.; Lin, J. STAT3 signaling pathway is necessary for cell survival and tumorsphere forming capacity in ALDH(+)/CD133(+) stem cell-like human colon cancer cells. Biochem. Biophys. Res. Commun. 2011, 416, 246–251. [Google Scholar] [CrossRef]
- Nicolis, S.K. Cancer stem cells and “stemness” genes in neuro-oncology. Neurobiol. Dis. 2007, 25, 217–229. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Wlodkowic, D.; Telford, W.; Skommer, J.; Darzynkiewicz, Z. Apoptosis and beyond: Cytometry in studies of programmed cell death. Methods Cell Biol. 2011, 103, 55–98. [Google Scholar] [CrossRef]
- Horibe, T.; Hoogenraad, N.J. The chop gene contains an element for the positive regulation of the mitochondrial unfolded protein response. PLoS One 2007, 2, e835. [Google Scholar] [CrossRef]
- Baumeister, P.; Luo, S.; Skarnes, W.C.; Sui, G.; Seto, E.; Shi, Y.; Lee, A.S. Endoplasmic reticulum stress induction of the Grp78/BiP promoter: Activating mechanisms mediated by YY1 and its interactive chromatin modifiers. Mol. Cell. Biol. 2005, 25, 4529–4540. [Google Scholar] [CrossRef]
- van Schadewijk, A.; van't Wout, E.F.; Stolk, J.; Hiemstra, P.S. A quantitative method for detection of spliced X-box binding protein-1 (XBP1) mRNA as a measure of endoplasmic reticulum (ER) stress. Cell Stress Chaperones 2012, 17, 275–279. [Google Scholar] [CrossRef]
- Deryugina, E.I.; Quigley, J.P. Matrix metalloproteinases and tumor metastasis. Cancer Metastasis Rev. 2006, 25, 9–34. [Google Scholar] [CrossRef]
- Masson, V.; de la Ballina, L.R.; Munaut, C.; Wielockx, B.; Jost, M.; Maillard, C.; Blacher, S.; Bajou, K.; Itoh, T.; Itohara, S.; et al. Contribution of host MMP-2 and MMP-9 to promote tumor vascularization and invasion of malignant keratinocytes. FASEB J. 2005, 19, 234–236. [Google Scholar]
- Hadler-Olsen, E.; Winberg, J.O.; Uhlin-Hansen, L. Matrix metalloproteinases in cancer: Their value as diagnostic and prognostic markers and therapeutic targets. Tumour Biol. 2013, 34, 2041–2051. [Google Scholar] [CrossRef]
- Liu, T.J.; Sun, B.C.; Zhao, X.L.; Zhao, X.M.; Sun, T.; Gu, Q.; Yao, Z.; Dong, X.Y.; Zhao, N.; Liu, N. CD133+ cells with cancer stem cell characteristics associates with vasculogenic mimicry in triple-negative breast cancer. Oncogene 2013, 32, 544–553. [Google Scholar] [CrossRef]
- Li, F.; Tiede, B.; Massague, J.; Kang, Y. Beyond tumorigenesis: Cancer stem cells in metastasis. Cell Res. 2007, 17, 3–14. [Google Scholar] [CrossRef]
- Anderson, E.C.; Hessman, C.; Levin, T.G.; Monroe, M.M.; Wong, M.H. The Role of Colorectal Cancer Stem Cells in Metastatic Disease and Therapeutic Response. Cancers 2011, 3, 319–339. [Google Scholar] [CrossRef]
- Yeh, C.C.; Yang, J.I.; Lee, J.C.; Tseng, C.N.; Chan, Y.C.; Hseu, Y.C.; Tang, J.Y.; Chuang, L.Y.; Huang, H.W.; Chang, F.R.; Chang, H.W. Anti-proliferative effect of methanolic extract of Gracilaria tenuistipitata on oral cancer cells involves apoptosis, DNA damage, and oxidative stress. BMC Complement Altern. Med. 2012, 12, 142. [Google Scholar] [CrossRef]
- Chang, Y.F.; Chou, H.J.; Yen, Y.C.; Chang, H.W.; Hong, Y.R.; Huang, H.W.; Tseng, C.N. Agrin induces association of Chrna1 mRNA and nicotinic acetylcholine receptor in C2C12 myotubes. FEBS Lett. 2012, 586, 3111–3116. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chiu, C.C.; Liu, P.L.; Huang, K.J.; Wang, H.M.; Chang, K.F.; Chou, C.K.; Chang, F.R.; Chong, I.W.; Fang, K.; Chen, J.S.; Chang, H.W.; et al. Goniothalamin inhibits growth of human lung cancer cells through DNA damage, apoptosis, and reduced migration ability. J. Agric. Food. Chem. 2011, 59, 4288–4293. [Google Scholar] [CrossRef]
- Sample Availability: Brefeldin A (B-8500) was purchased from LC Laboratories (Woburn, MA, USA).
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tseng, C.-N.; Huang, C.-F.; Cho, C.-L.; Chang, H.-W.; Huang, C.-W.; Chiu, C.-C.; Chang, Y.-F. Brefeldin A Effectively Inhibits Cancer Stem Cell-Like Properties and MMP-9 Activity in Human Colorectal Cancer Colo 205 Cells. Molecules 2013, 18, 10242-10253. https://doi.org/10.3390/molecules180910242
Tseng C-N, Huang C-F, Cho C-L, Chang H-W, Huang C-W, Chiu C-C, Chang Y-F. Brefeldin A Effectively Inhibits Cancer Stem Cell-Like Properties and MMP-9 Activity in Human Colorectal Cancer Colo 205 Cells. Molecules. 2013; 18(9):10242-10253. https://doi.org/10.3390/molecules180910242
Chicago/Turabian StyleTseng, Chao-Neng, Chien-Fu Huang, Chung-Lung Cho, Hsueh-Wei Chang, Chao-Wei Huang, Chien-Chih Chiu, and Yung-Fu Chang. 2013. "Brefeldin A Effectively Inhibits Cancer Stem Cell-Like Properties and MMP-9 Activity in Human Colorectal Cancer Colo 205 Cells" Molecules 18, no. 9: 10242-10253. https://doi.org/10.3390/molecules180910242
APA StyleTseng, C.-N., Huang, C.-F., Cho, C.-L., Chang, H.-W., Huang, C.-W., Chiu, C.-C., & Chang, Y.-F. (2013). Brefeldin A Effectively Inhibits Cancer Stem Cell-Like Properties and MMP-9 Activity in Human Colorectal Cancer Colo 205 Cells. Molecules, 18(9), 10242-10253. https://doi.org/10.3390/molecules180910242