Telmisartan Exerts Anti-Tumor Effects by Activating Peroxisome Proliferator-Activated Receptor-γ in Human Lung Adenocarcinoma A549 Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
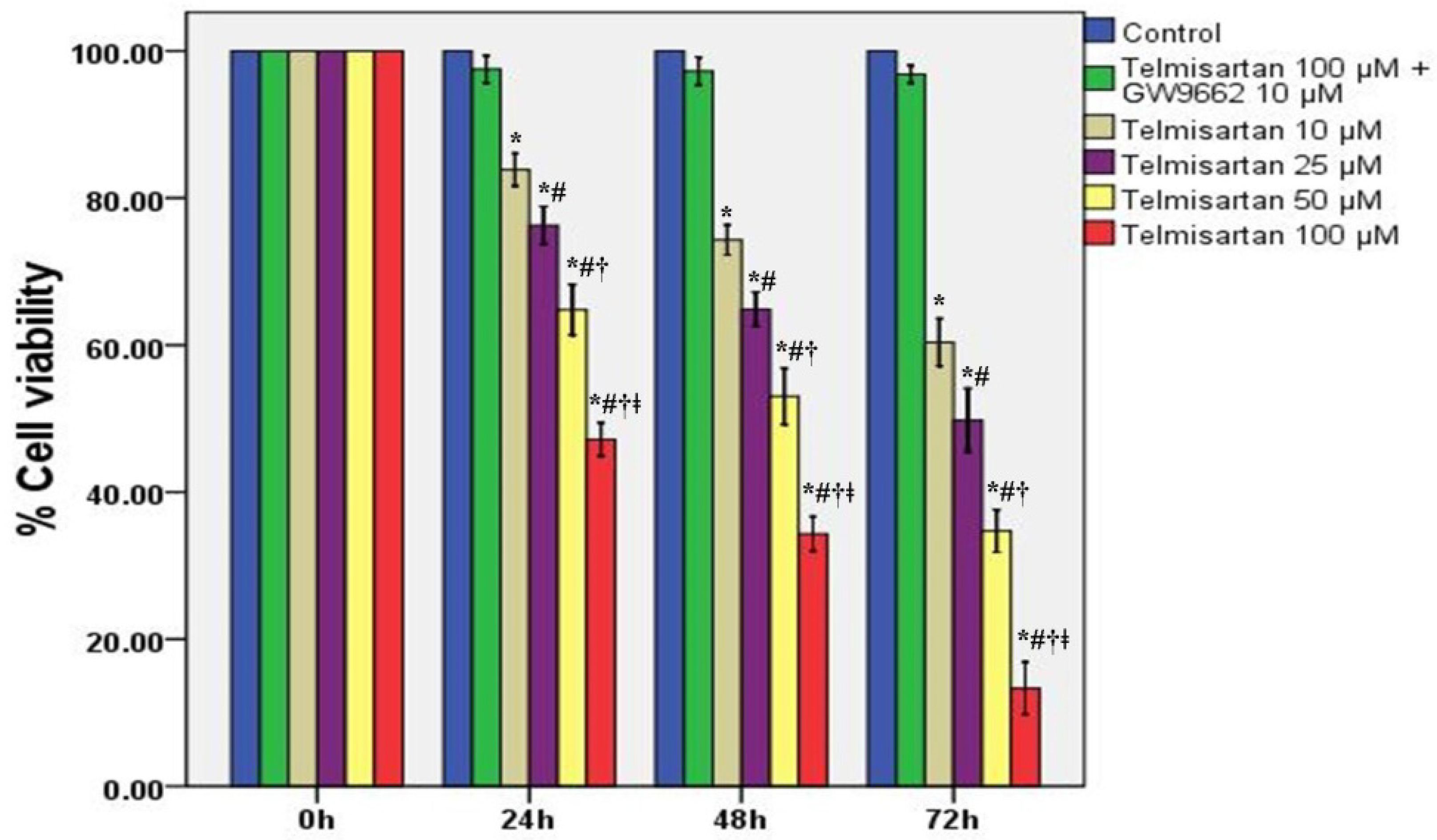
2.1. Telmisartan Inhibits the Cell Survival Rates and Cell Viabilities of A549 Cells


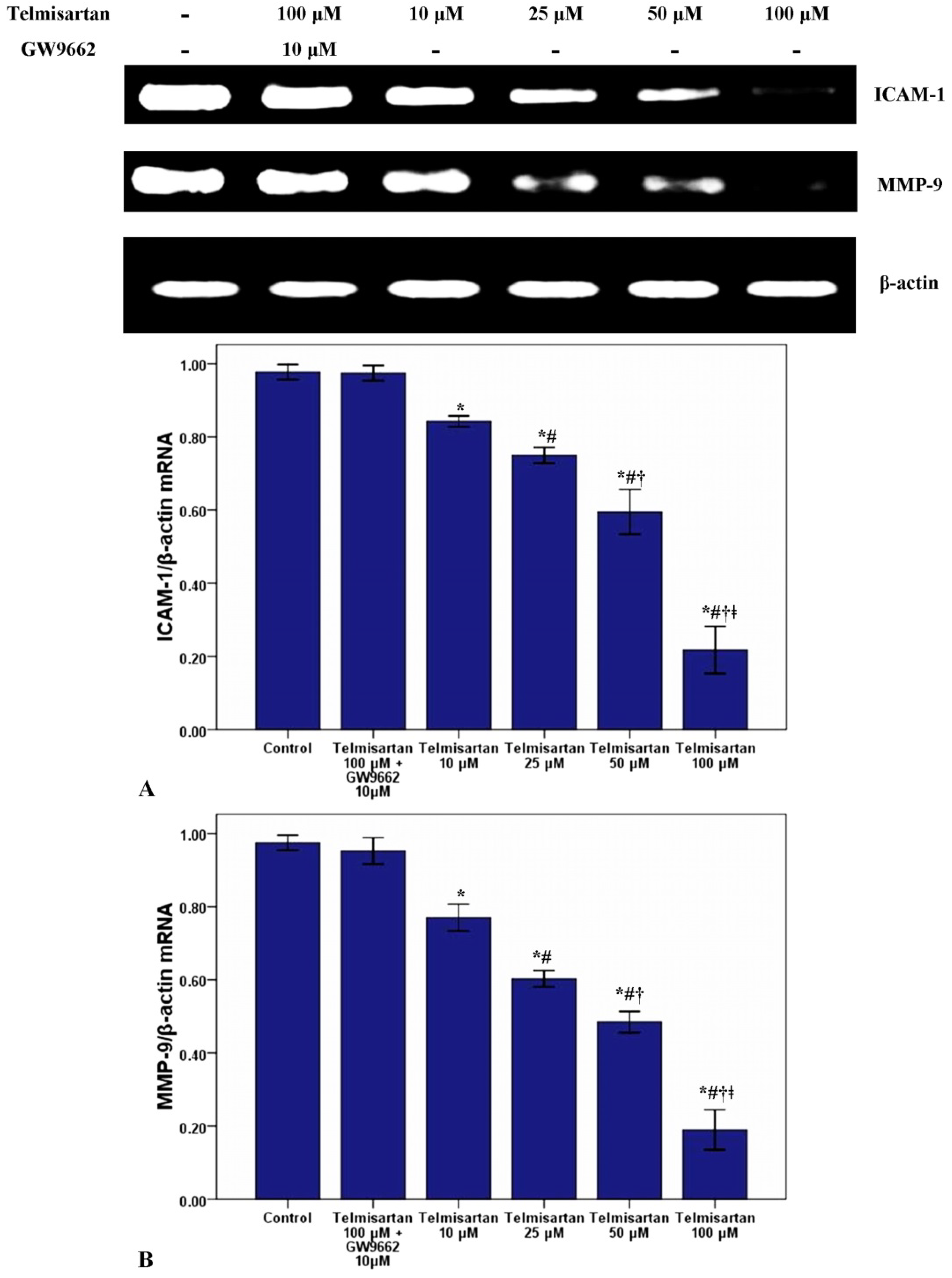
2.2. Telmisartan Dose-Dependently Reduces the mRNA and Protein Expression of ICAM-1 and MMP-9 in A549 Cells
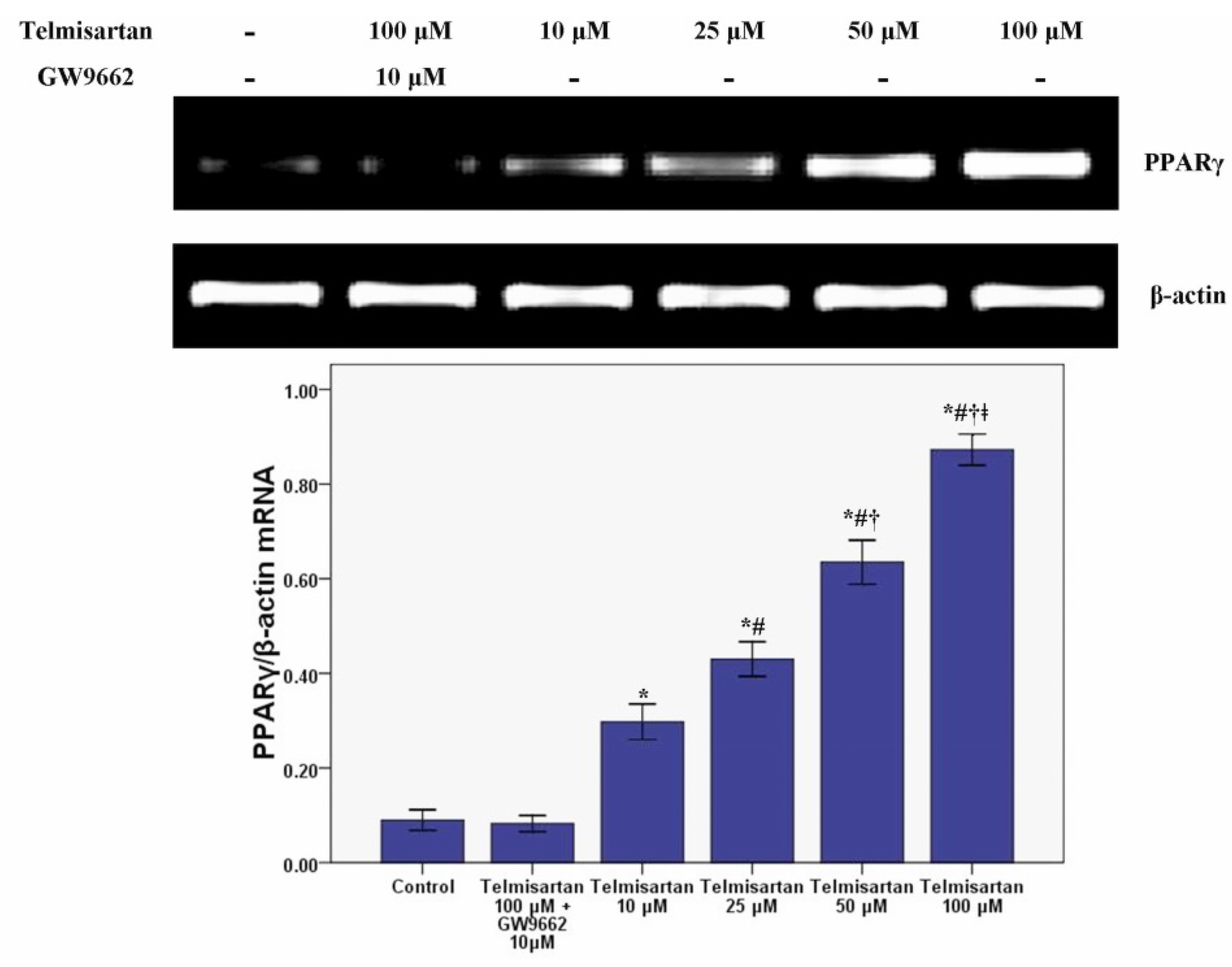
2.3. Telmisartan Dose-Dependently Enhances the mRNA and Protein Expression of PPARγ in A549 Cells
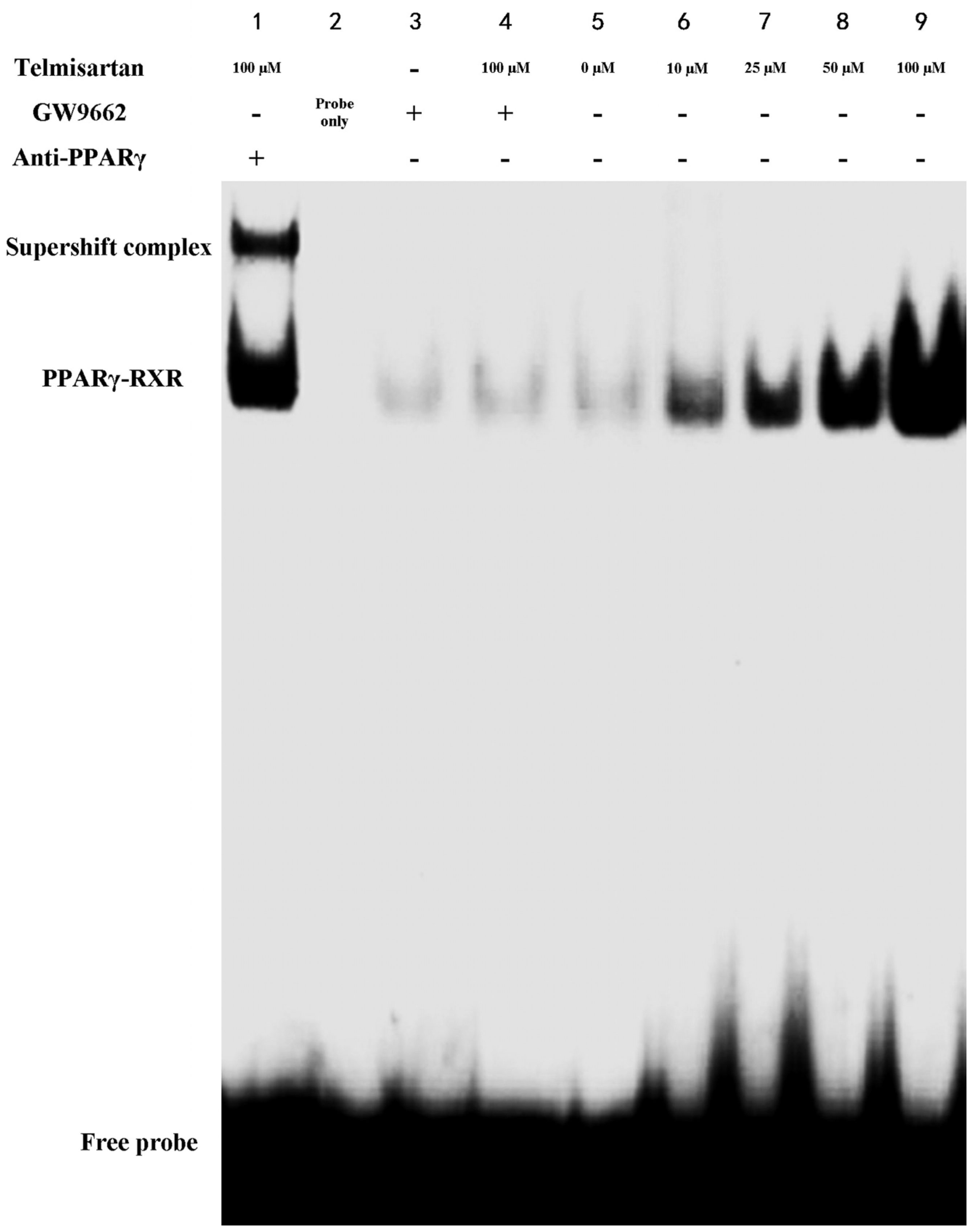
2.4. Telmisartan Increases DNA Binding Activity of PPARγ in A549 Cells





2.5. Discussion
3. Experimental
3.1. Reagents
3.2. Cell Culture, MTT Assay and Trypan Blue Exclusion
3.3. RT-PCR (Reverse Transcription-Polymerase Chain Reaction)
3.4. Western Blotting
3.5. Electrophoretic Mobility Shift Assay (EMSA)
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Conflictts of Interest
References
- Rosenson, R.S.; Wright, R.S.; Farkouh, M.; Plutzky, J. Modulating peroxisome proliferator-activated receptors for therapeutic benefit? Biology, clinical experience, and future prospects. Am. Heart J. 2012, 164, 672–680. [Google Scholar] [CrossRef]
- Choi, J.M.; Bothwell, A.L. The nuclear receptor PPARs as important regulators of T-cell functions and autoimmune diseases. Mol. Cell 2012, 33, 217–222. [Google Scholar]
- Sugawara, A.; Uruno, A.; Matsuda, K.; Saito-Ito, T.; Funato, T.; Saito-Hakoda, A.; Kudo, M.; Ito, S. Effects of PPARγ agonists against vascular and renal dysfunction. Curr. Mol. Pharmacol. 2012, 5, 248–254. [Google Scholar] [CrossRef]
- Bishop-Bailey, D. PPARs and angiogenesis. Biochem. Soc. Trans. 2011, 39, 1601–1605. [Google Scholar] [CrossRef]
- Xiao, M.; Zhu, T.; Wang, T.; Wen, F.Q. Diammonium glycyrrhizinate inhibits airway smooth muscle proliferation in a murine model of chronic asthma. Nan Fang. Yi Ke Da Xue Xue Bao 2013, 33, 1416–1420. [Google Scholar]
- Deng, Y.L.; Xiong, X.Z.; Cheng, N.S. Organ fibrosis inhibited by blocking transforming growth factor-β signaling via peroxisome proliferator-activated receptor γ agonists. Hepatobiliary Pancreat. Dis. Int. 2012, 11, 467–478. [Google Scholar] [CrossRef]
- Yeh, S.L.; Yeh, C.L.; Chan, S.T.; Chuang, C.H. Plasma rich in quercetin metabolites induces G2/M arrest by upregulating PPAR-γ expression in human A549 lung cancer cells. Planta Med. 2011, 77, 992–998. [Google Scholar] [CrossRef]
- Girnun, G.D. The diverse role of the PPARγ coactivator 1 family of transcriptional coactivators in cancer. Semin. Cell Dev. Biol. 2012, 23, 381–388. [Google Scholar] [CrossRef]
- Peters, J.M.; Shah, Y.M.; Gonzalez, F.J. The role of peroxisome proliferator-activated receptors in carcinogenesis and chemoprevention. Nat. Rev. Cancer 2012, 12, 181–195. [Google Scholar]
- Thompson, E.A. PPARgamma physiology and pathology in gastrointestinal epithelial cells. Mol. Cell 2007, 24, 167–176. [Google Scholar]
- Wang, D.; Dubois, R.N. Peroxisome proliferator-activated receptors and progression of colorectal cancer. PPAR Res. 2008, 931074. [Google Scholar]
- Robbins, G.T.; Nie, D. PPAR gamma, bioactive lipids, and cancer progression. Front. Biosci. 2012, 17, 1816–1834. [Google Scholar] [CrossRef]
- Yates, C.M.; McGettrick, H.M.; Nash, G.B.; Rainger, G.E. Adhesion of tumor cells to matrices and endothelium. Methods Mol. Biol. 2014, 1070, 57–75. [Google Scholar]
- Hung, P.F.; Hong, T.M.; Hsu, Y.C.; Chen, H.Y.; Chang, Y.L.; Wu, C.T.; Chang, G.C.; Jou, Y.S.; Pan, S.H.; Yang, P.C. The motor protein KIF14 inhibits tumor growth and cancer metastasis in lung adenocarcinoma. PLoS One 2013, 8, e61664. [Google Scholar] [CrossRef]
- Jia, W.; Martin, T.A.; Zhang, G.; Jiang, W.G. Junctional adhesion molecules in cerebral endothelial tight junction and brain metastasis. Anticancer Res. 2013, 33, 2353–2359. [Google Scholar]
- Liu, S.Q.; Su, Y.J.; Qin, M.B.; Mao, Y.B.; Huang, J.A.; Tang, G.D. Sphingosine kinase 1 promotes tumor progression and confers malignancy phenotypes of colon cancer by regulating the focal adhesion kinase pathway and adhesion molecules. Int. J. Oncol. 2013, 42, 617–626. [Google Scholar]
- Zheng, F.J.; Shi, L.; Yang, J.; Deng, X.H.; Wu, Y.Q.; Yan, X.Q.; Huang, N. Effect of tea polyphenols on the adhesion of highly metastatic human lung carcinoma cell lines to endothelial cells in vitro. Asian Pac. J. Cancer Prev. 2012, 13, 3751–3755. [Google Scholar] [CrossRef]
- Traub, S.; Nikonova, A.; Carruthers, A.; Dunmore, R.; Vousden, K.A.; Gogsadze, L.; Hao, W.; Zhu, Q.; Bernard, K.; Zhu, J.; et al. An anti-human ICAM-1 antibody inhibits rhinovirus-induced exacerbations of lung inflammation. PLoS Pathog. 2013, 9, e1003520. [Google Scholar] [CrossRef] [Green Version]
- Zumwalde, N.A.; Domae, E.; Mescher, M.F.; Shimizu, Y. ICAM-1-dependent homotypic aggregates regulate CD8 T cell effector function and differentiation during T cell activation. J. Immunol. 2013, 191, 3681–3693. [Google Scholar] [CrossRef]
- Dorecka, M.; Siemianowicz, K.; Francuz, T.; Garczorz, W.; Chyra, A.; Klych, A.; Romaniuk, W. Exendin-4 and GLP-1 decreases induced expression of ICAM-1, VCAM-1 and RAGE in human retinal pigment epithelial cells. Pharmacol. Rep. 2013, 65, 884–890. [Google Scholar]
- Yu, J.A.; Sadaria, M.R.; Meng, X.; Mitra, S.; Ao, L.; Fullerton, D.A.; Weyant, M.J. Lung cancer cell invasion and expression of intercellular adhesion molecule-1 (ICAM-1) are attenuated by secretory phospholipase A2 inhibition. J. Thorac. Cardiovasc. Surg. 2012, 143, 405–411. [Google Scholar] [CrossRef]
- Lin, Y.C.; Shun, C.T.; Wu, M.S.; Chen, C.C. A novel anticancer effect of thalidomide: Inhibition of intercellular adhesion molecule-1-mediated cell invasion and metastasis through suppression of nuclear factor-kappaB. Clin. Cancer Res. 2006, 12, 7165–7173. [Google Scholar] [CrossRef]
- Kotteas, E.A.; Gkiozos, I.; Tsagkouli, S.; Bastas, A.; Ntanos, I.; Saif, M.W.; Syrigos, K.N. Soluble ICAM-1 levels in small-cell lung cancer: Prognostic value for survival and predictive significance for response during chemotherapy. Med. Oncol. 2013, 30, 662. [Google Scholar] [CrossRef]
- Cianchetti, S.; del Fiorentino, A.; Colognato, R.; di Stefano, R.; Franzoni, F.; Pedrinelli, R. Anti-inflammatory and anti-oxidant properties of telmisartan in cultured human umbilical vein endothelial cells. Atherosclerosis 2008, 198, 22–28. [Google Scholar] [CrossRef]
- Celiński, K.; Madro, A.; Prozorow-Król, B.; Korolczuk, A.; Cichoz-Lach, H.; Słomka, M.; Korobowicz, E. Rosiglitazone, a peroxisome proliferator-activated receptor gamma (PPARgamma)-specific agonist, as a modulator in experimental acute pancreatitis. Med. Sci. Monit. 2009, 15, BR21–BR29. [Google Scholar]
- Arnold, R.; Neumann, M.; König, W. Peroxisome proliferator-activated receptor-gamma agonists inhibit respiratory syncytial virus-induced expression of intercellular adhesion molecule-1 in human lung epithelial cells. Immunology 2007, 121, 71–81. [Google Scholar] [CrossRef]
- Verma, R.P.; Hansch, C. Matrix metalloproteinases (MMPs): Chemical-biological functions and (Q)SARs. Bioorg. Med. Chem. 2007, 15, 2223–2268. [Google Scholar] [CrossRef]
- Gao, X.H.; Yang, X.Q.; Wang, B.C.; Liu, S.P.; Wang, F.B. Overexpression of twist and matrix metalloproteinase-9 with metastasis and prognosis in gastric cancer. Asian Pac. J. Cancer Prev. 2013, 14, 5055–5060. [Google Scholar]
- Ramanujum, R.; Lin, Y.L.; Liu, J.K.; He, S. Regulatory expression of MMP-8/MMP-9 and inhibition of proliferation, migration and invasion in human lung cancer A549 cells in the presence of HGF variants. Kaohsiung J. Med. Sci. 2013, 29, 530–539. [Google Scholar] [CrossRef]
- Xue, J.; Hua, Y.N.; Xie, M.L.; Gu, Z.L. Aspirin inhibits MMP-9 mRNA expression and release via the PPARalpha/gamma and COX-2/mPGES-1-mediated pathways in macrophages derived from THP-1 cells. Biomed. Pharmacother. 2010, 64, 118–123. [Google Scholar] [CrossRef]
- Marx, N.; Froehlich, J.; Siam, L.; Ittner, J.; Wierse, G.; Schmidt, A.; Scharnagl, H.; Hombach, V.; Koenig, W. Antidiabetic PPAR gamma-activator rosiglitazone reduces MMP-9 serum levels in type 2 diabetic patients with coronary artery disease. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 283–288. [Google Scholar] [CrossRef]
- Liu, J.; Lu, H.; Huang, R.; Lin, D.; Wu, X.; Lin, Q.; Wu, X.; Zheng, J.; Pan, X.; Peng, J.; et al. Peroxisome proliferator activated receptor-gamma ligands induced cell growth inhibition and its influence on matrix metalloproteinase activity in human myeloid leukemia cells. Cancer Chemother. Pharmacol. 2005, 56, 400–408. [Google Scholar] [CrossRef]
- Chaykovska, L.; Alter, M.L.; von Websky, K.; Hohmann, M.; Tsuprykov, O.; Reichetzeder, C.; Kutil, B.; Kraft, R.; Klein, T.; Hocher, B. Effects of telmisartan and linagliptin when used in combination on blood pressure and oxidative stress in rats with 2-kidney-1-clip hypertension. J. Hypertens. 2013, 31, 2290–2299. [Google Scholar] [CrossRef]
- Siragusa, M.; Sessa, W.C. Telmisartan exerts pleiotropic effects in endothelial cells and promotes endothelial cell quiescence and survival. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1852–1860. [Google Scholar] [CrossRef]
- Chen, Y.; Luo, Q.; Xiong, Z.; Liang, W.; Chen, L.; Xiong, Z. Telmisartan counteracts TGF-β1 induced epithelial-to-mesenchymal transition via PPAR-γ in human proximal tubule epithelial cells. Int. J. Clin. Exp. Pathol. 2012, 5, 522–529. [Google Scholar]
- Shigeto, T.; Yokoyama, Y.; Xin, B.; Mizunuma, H. Peroxisome proliferator-activated receptor alpha and gamma ligands inhibit the growth of human ovarian cancer. Oncol. Rep. 2007, 18, 833–840. [Google Scholar]
- Wei, D.; Peng, J.J.; Gao, H.; Li, H.; Li, D.; Tan, Y.; Zhang, T. Digoxin Downregulates NDRG1 and VEGF through the Inhibition of HIF-1α under Hypoxic Conditions in Human Lung Adenocarcinoma A549 Cells. Int. J. Mol. Sci. 2013, 14, 7273–7285. [Google Scholar] [CrossRef]
- Tang, Z.P.; Cui, Q.Z.; Dong, Q.Z.; Xu, K.; Wang, E.H. Ataxia-telangiectasia group D complementing gene (ATDC) upregulates matrix metalloproteinase 9 (MMP-9) to promote lung cancer cell invasion by activating ERK and JNK pathways. Tumour Biol. 2013, 34, 2835–2842. [Google Scholar] [CrossRef]
- Karami, H.; Baradaran, B.; Esfahani, A.; Estiar, M.A.; Naghavi-Behzad, M.; Sakhinia, M.; Sakhinia, E. siRNA-Mediated silencing of survivin inhibits proliferation and enhances etoposide chemosensitivity in acute myeloid leukemia cells. Asian Pac. J. Cancer Prev. 2013, 14, 7719–7724. [Google Scholar]
- Sample Availability: Samples of telmisartan and GW9662 are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, J.; Chen, L.; Yu, P.; Liu, B.; Zhu, J.; Yang, Y. Telmisartan Exerts Anti-Tumor Effects by Activating Peroxisome Proliferator-Activated Receptor-γ in Human Lung Adenocarcinoma A549 Cells. Molecules 2014, 19, 2862-2876. https://doi.org/10.3390/molecules19032862
Li J, Chen L, Yu P, Liu B, Zhu J, Yang Y. Telmisartan Exerts Anti-Tumor Effects by Activating Peroxisome Proliferator-Activated Receptor-γ in Human Lung Adenocarcinoma A549 Cells. Molecules. 2014; 19(3):2862-2876. https://doi.org/10.3390/molecules19032862
Chicago/Turabian StyleLi, Juan, Lin Chen, Ping Yu, Bin Liu, Jiang Zhu, and Ye Yang. 2014. "Telmisartan Exerts Anti-Tumor Effects by Activating Peroxisome Proliferator-Activated Receptor-γ in Human Lung Adenocarcinoma A549 Cells" Molecules 19, no. 3: 2862-2876. https://doi.org/10.3390/molecules19032862