Antiadhesive Properties of Arabinogalactan Protein from Ribes nigrum Seeds against Bacterial Adhesion of Helicobacter pylori

Abstract
:1. Introduction
2. Results and Discussion
2.1. Isolation and Characterization of Arabinogalactan Protein F2 from R. nigrum Seeds



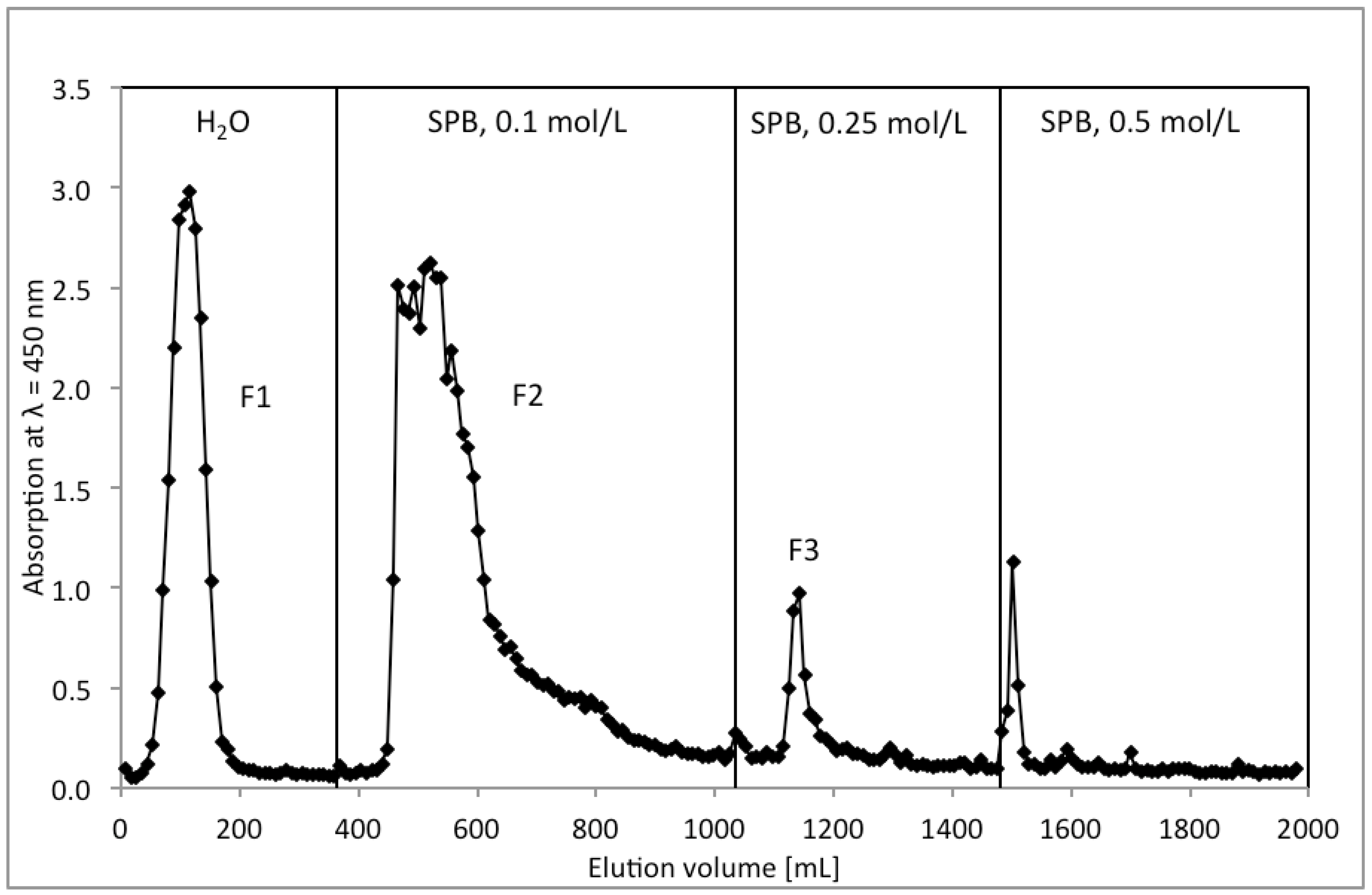{kind=link}
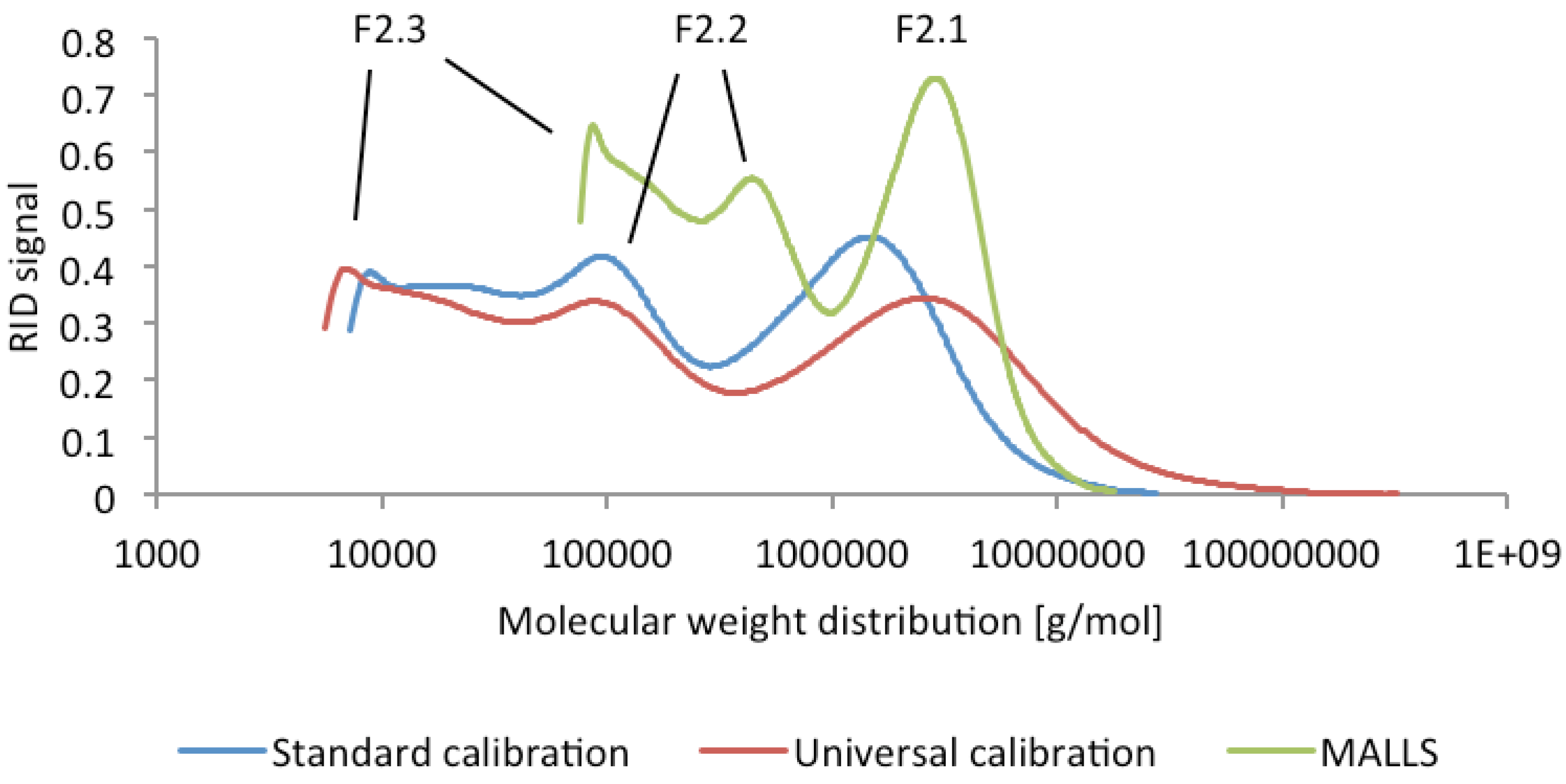{kind=link}
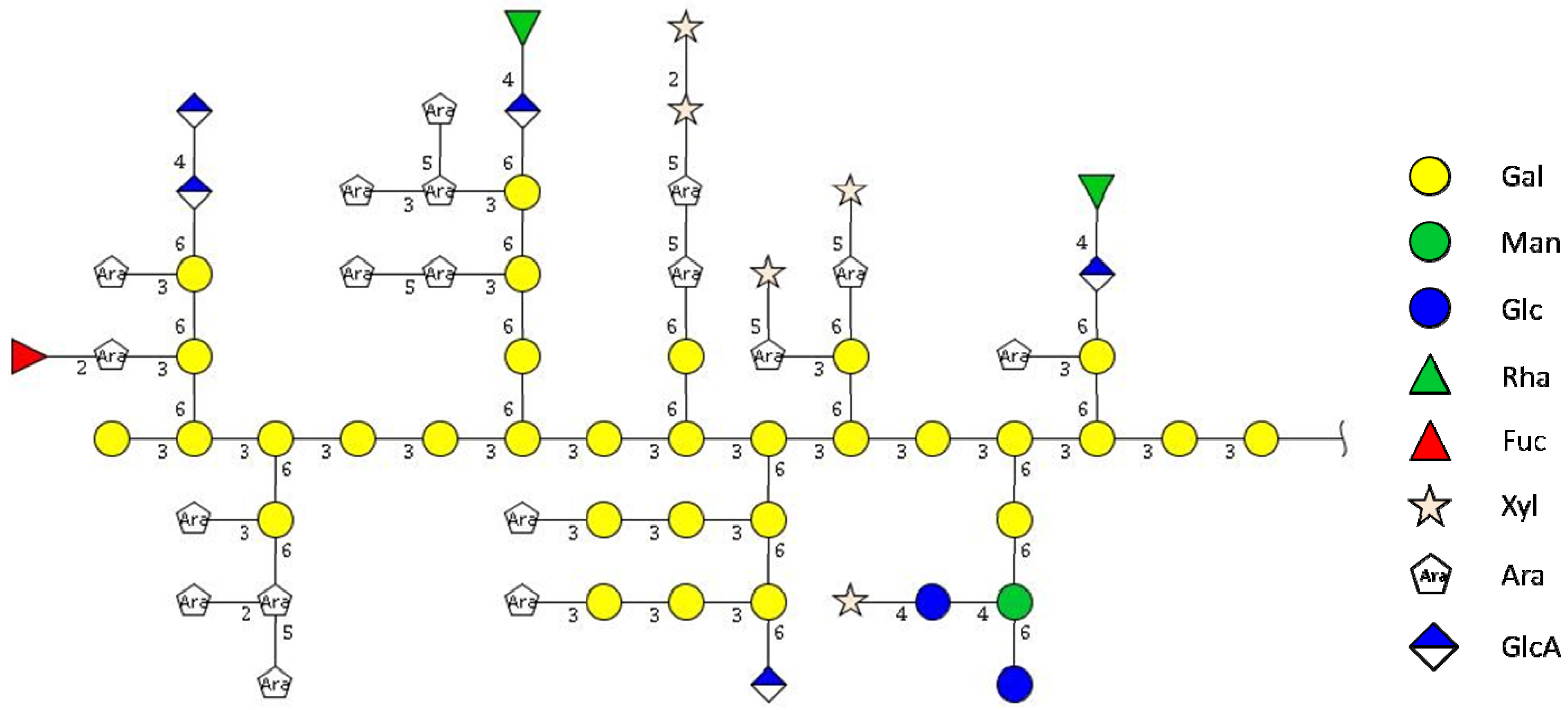{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amount [%] | Carbohydrate | HPAEC- PAD | Linkage type | Amount GC-MS | |
|---|---|---|---|---|---|
| (mol %) | (mol %) * | ||||
| Neutral carbohydrates | 76 | d-Gal | 28.3 | 1-p | 0.3 |
| 1,3-p | 13.5 | ||||
| 1,4-p | 0.1 | ||||
| 1,6-p | 0.3 | ||||
| 1,3,6-p | 14.1 | ||||
| l-Ara | 24.5 | 1-f | 4.2 | ||
| 1,2-f | 0.3 | ||||
| 1,5-f | 10.9 | ||||
| 1,2,5-f | 8.6 | ||||
| 1,3,5-f | 0.5 | ||||
| d-Xyl | 15.7 | 1-p | 9.2 | ||
| 1,2-p | 6.5 | ||||
| d-Man | 3.5 | 1,2-p | 0.1 | ||
| 1,4-p | 1.0 | ||||
| 1,4,6-p | 2.4 | ||||
| l-Rha | 2.4 | 1-p | tr. | ||
| 1,2,4-p | 2.4 | ||||
| d-Glc | 1.0 | 1-p | 0.4 | ||
| 1,4-p | 0.6 | ||||
| l-Fuc | 0.6 | 1-p | 0.6 | ||
| Uronic acids | 23 **** | d-GalA | 16.0 | 1,4-p | 16.0 |
| d-GlcA | 7.0 | 1-p | 1.4 | ||
| 1,4-p | 5.6 | ||||
| Protein | 1.0 ** 2.1*** | Amino acid | [mol %] | ||
| Histidine | 12.1 | ||||
| Serine | 12.1 | ||||
| Hydroxyproline | 10.9 | ||||
| Glycine | 9.8 | ||||
| Alanine | 8.7 | ||||
| Valine | 7.9 | ||||
| Lysine | 7.3 | ||||
| Threonine | 6.2 | ||||
| Glutamic acid/glutamine | 5.7 | ||||
| Cysteine | 5.4 | ||||
| Phenylalanine | 3.8 | ||||
| Isoleucine | 3.7 | ||||
| Leucine | 2.3 | ||||
| Proline | 1.8 | ||||
| Tyrosine | 1.0 | ||||
| Aspartic acid/asparagine | 0.6 | ||||
| Methionine | 0.3 | ||||
| Tryptophan | n.d. | ||||
| Arginine | n.d. | ||||
| Norvaline | n.d. | ||||

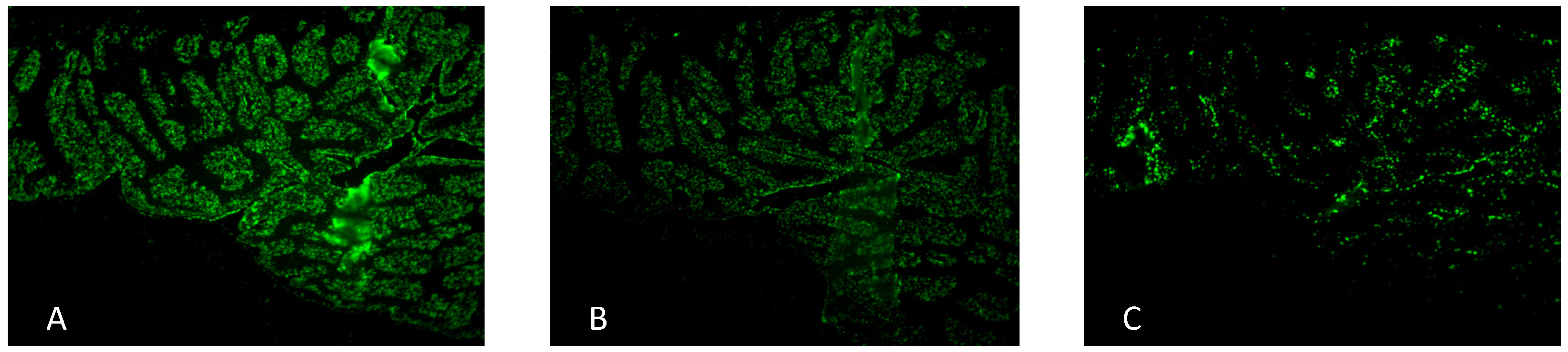
2.2. Antiadhesive Activity of F2 against H. pylori

| H. pylori pre-treated with | concentration | Blinded fluorescence microscopy evaluation * | rel. fluorescence area (%) |
|---|---|---|---|
| A untreated negative control | -- | + + + + + | 100 |
| B Arabinogalactan protein F2 | 1.0 mg/mL | + + + | 75 |
| C Acidic human milk oligosaccharides (positive control) | 1.0 mg/mL | + + | 13 |


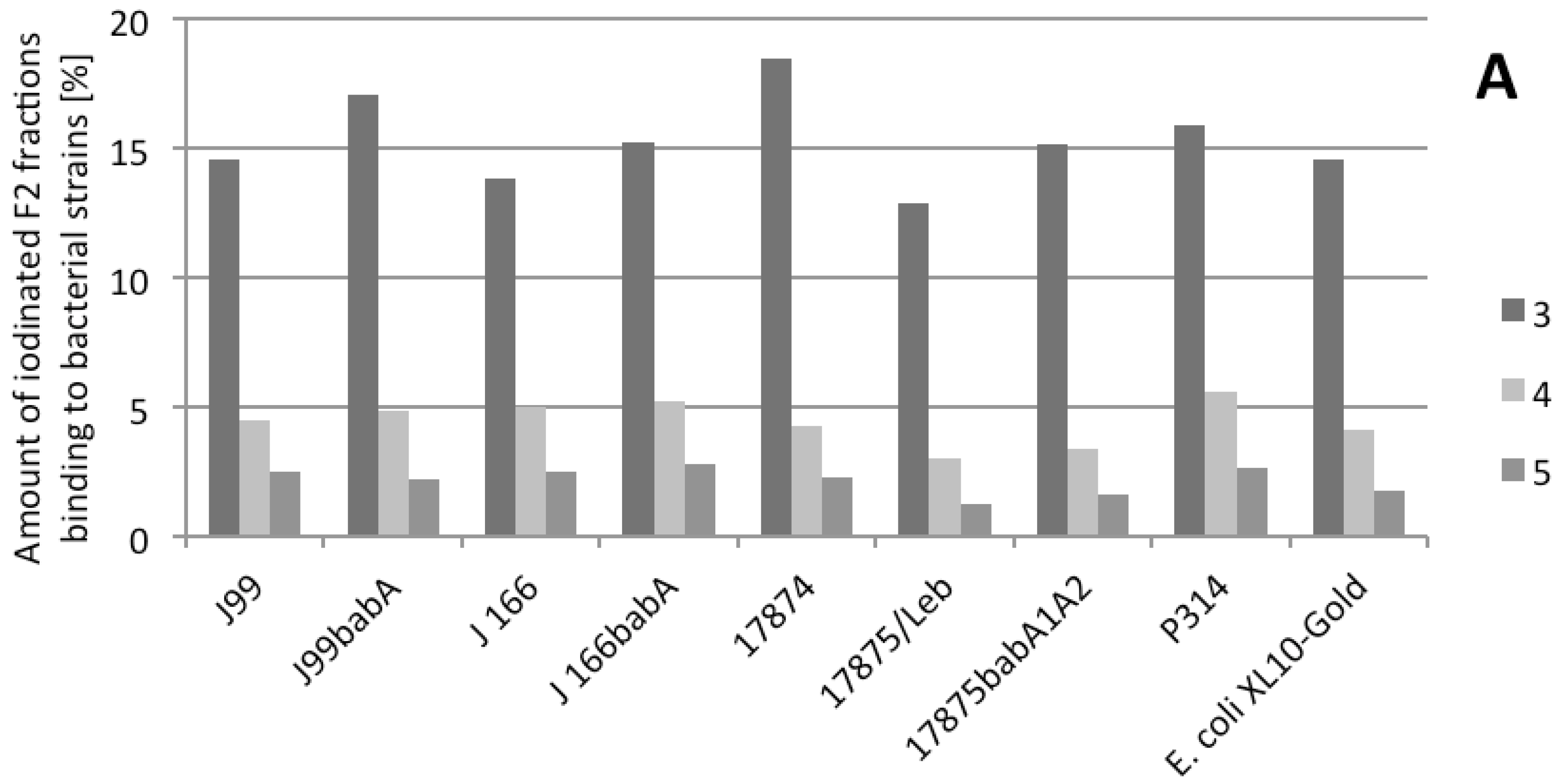
2.3. Binding of Radiolabeled F2 to Different Strains

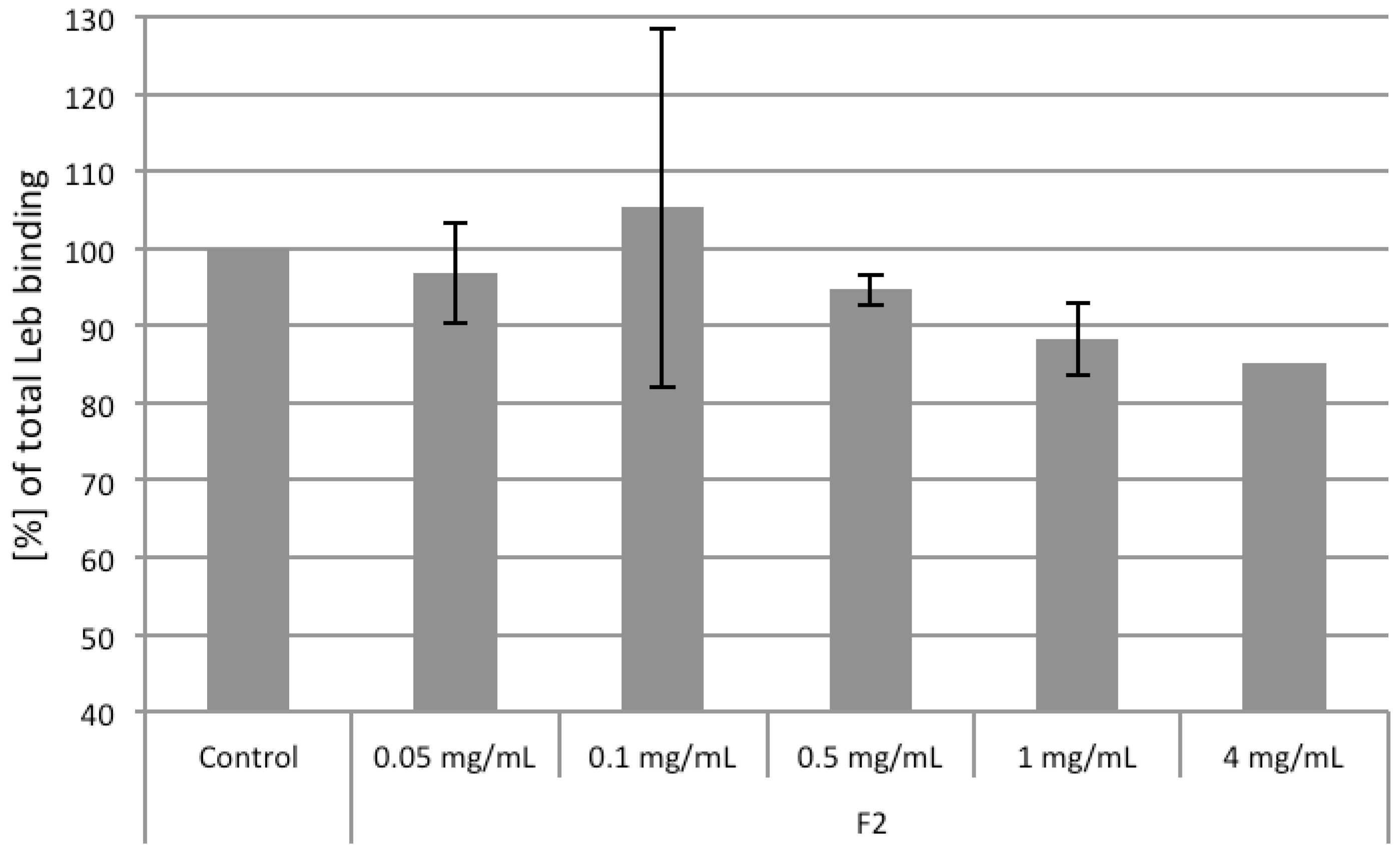
2.4. Competiton of Arabinogalactan Protein F2 with Leb and sLex Ligands

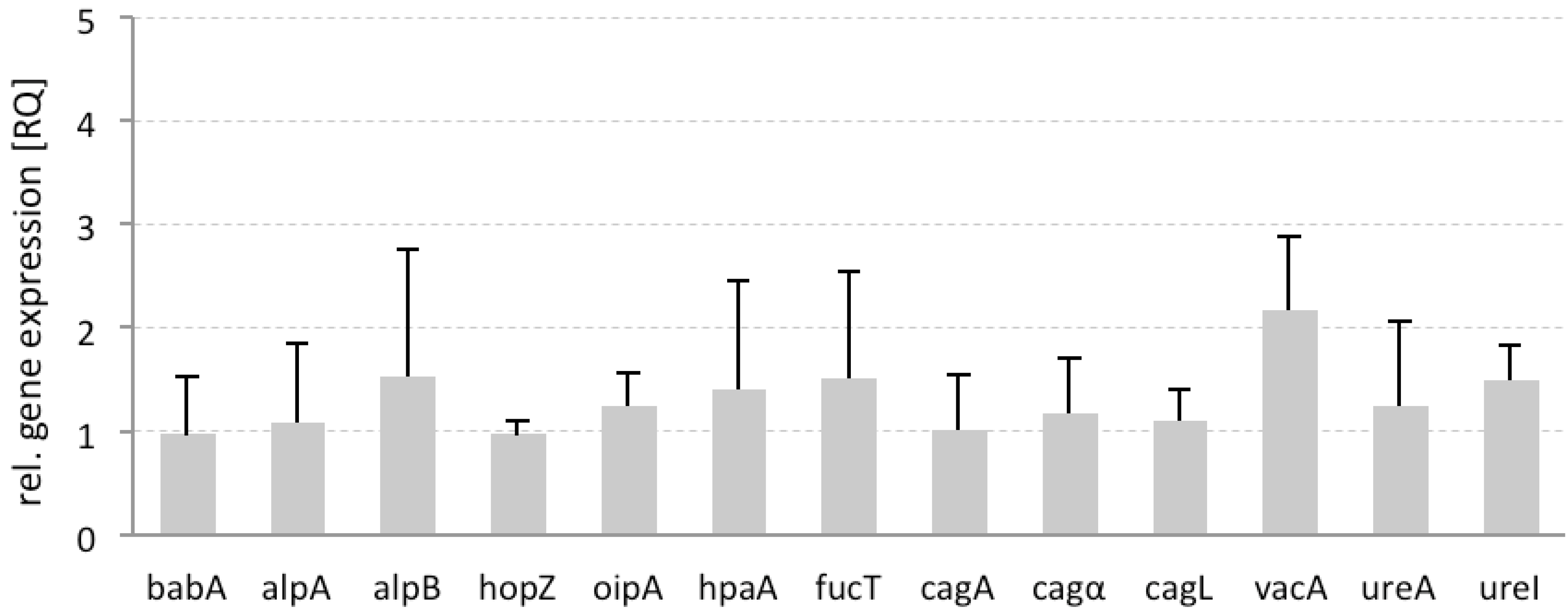
2.5. Differential Gene Expression

2.6. Discussion
| Primer | Gene/locus tag (Ref.) | Sequence, 5' → 3' orientation | |
|---|---|---|---|
| 23S rRNA | jhpr2 [44] | F | CCTTAGATTTACGGCGGATACA |
| R | CAGTGGTAGAAGAGCGTTCATA | ||
| P | TCACTTCATACCGCTCC | ||
| AlpA | jhp0848 [44] | F | GCTCACTAAAAACACCATT |
| R | CATGCTTCTCACCTGATTGTTG | ||
| P | TCCAACCAAGCTAACGC | ||
| AlpB | jhp0849 [44] | F | GAGTCAAAACATCAGCAAGA |
| R | ACTGAGCTGGTTGGAGAGATT | ||
| P | GACAACAACACCACGA | ||
| BabA | jhp0833 [44] | F | GATTTGTTATCGTTTGTCCTAC |
| R | AGGCTTAGCGGGACTTTT | ||
| P | GTTGATGG*GTTGTTGC | ||
| HopZ | jhp0007 [44] | F | CCAAGAAATCGTAACGCAAG |
| R | TGTTTTGAGCGAAAGCCTATC | ||
| P | TCCTTACACCTCTGCT | ||
| OipA | jhp0581 [44] | F | ATAAGCGAGCGTGTCAAGAA |
| R | ATGCCAATCACAAGCCCTGAA | ||
| P | GAAAGAAGG*GTAAAAGG | ||
| HpaA | jhp0733 [44] | F | CAAACCAGTGGAGAATAATAAC |
| R | GGATAGCAGCGATAAAGACGAT | ||
| P | CCATTCATAGCGACAG | ||
| FucT | jhp0596 [44] | F | TGCAAGTATCTCACGTAATCAA |
| R | CTCAAGGCTATGGCTATGTAAC | ||
| P | TGG*GAATGGTGTGGCT | ||
| CagA | jhp0495 [44] | F | TGGCAGTGGGTTAGTCATA |
| R | CCTGTGAGTTGGTCTTCTTTGT | ||
| P | AGGTGGTGAGAAAGG*GA | ||
| CagL | jhp0487 [44] | F | CTCGATTTTCAGCTTCCC |
| R | TCAATCCCTTAGACCAAAAGACT | ||
| P | ATTCCGCATTGTTGCT | ||
| Cagα | jhp1344 [44] | F | GAGACAAGCTCCATGAGA |
| R | ACCCCCGGTTCATAAAGACT | ||
| P | ACTTATTCTCCCACTTGC | ||
| VacA | jhp0819 [44] | F | AAACGACAAGAAAGAGATCAGT |
| R | CCAGCAAAAGGCCCATCAA | ||
| P | CAATAGCAACACAGAGG | ||
| Ure I | jhp0066 [44] | F | AGTGTTGATCGCTACGAATAAG |
| R | AGCGACTGGGTTATTGTTTGG | ||
| P | AGTGTGGTTGATAGCGG | ||
| UreA | jhp0068 [44] | F | TTGCCTTCGTTGATAGTGATG |
| R | CTGATGGGACCAAACTCGTAA | ||
| P | AACAACTCACCAGGAA |
3. Experimental
3.1. Isolation of R. nigrum Arabinogalactan Protein (AGP)
3.2. Glycoprotein Analysis
3.3. Bacteria and Growth Conditions
3.4. Cell Culture
3.5. Radiolabeling of Arabinogalactan F2 and Glycoconjugates
3.6. Analysis of Binding Properties of F2 towards Different H. pylori Strains by Radioimmunoassay
3.7. Inhibitory Effects of F2 on Lewisb Binding of H. pylori
3.8. Inhibitory Effects of F2 on Sialyl-Lewisx Binding of H. pylori
3.9. Statistics
4. Conclusions
Abbreviations
| AEC | anion exchange chromatography |
| AGP | arabinogalactan protein |
| BabA | blood group binding adhesion |
| F2 | arabinogalactan protein fraction from R. nigrum after AEC |
| FITC | fluorescein isothiocyanate |
| MALLS | multi angle laser light scattering |
| OMP | outer membrane protein |
| RT-PCR | real-time polymerase chain reaction |
| SabA | sialic acid binding adhesion |
Acknowledgments and Disclosures
Authors Contribution
Conflictts of Interest
References
- List, P.H.; Hörhammer, L. Ribes. Hagers Handbuch der Pharmazeutischen Praxis, 4th ed.; Hänsel, R., Keller, K., Rimpler, H., Schneider, G., Eds.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1979; Volume 6, pp. 132–135. [Google Scholar]
- Traitler, H.; Winter, H.; Richli, U.; Ingenbleek, Y. Characterization of γ-linolenic acid in Ribes seeds. Lipids 1984, 12, 923–928. [Google Scholar]
- Bakowska-Barczak, A.M.; Schieber, A.; Kolodziejczyk, P. Characterization of Canadian black currant (Ribes nigrum L.) seed oils and residues. J. Agric. Food Chem. 2009, 24, 11528–11536. [Google Scholar] [CrossRef]
- Zippel, J.; Deters, A.; Pappai, D.; Hensel, A. A high molecular arabinogalactan from Ribes nigrum L.: Influence on cell physiology of human skin fibroblasts and keratinocytes and internalization into cells via endosomal transport. Carbohydr. Res. 2009, 344, 1001–1008. [Google Scholar] [CrossRef]
- Lengsfeld, C.; Deters, A.; Faller, G.; Hensel, A. High molecular weight polysaccharides from black currant seed inhibit adhesion of Helicobacter pylori to human gastric mucosa. Planta Med. 2004, 70, 620–626. [Google Scholar] [CrossRef]
- Bendaoud, M.; Vinogradov, E.; Balashova, N.V.; Kadouri, D.E.; Kachlany, S.C.; Kapplan, J.B. Broad-Spectrum Biofilm Inhibition by Kingella kingae exopolysaccharide. J. Bacteriol. 2011, 193, 3879–3886. [Google Scholar] [CrossRef]
- Mysore, J.V.; Wigginton, T.; Simon, P.M.; Zopf, D.; Heman-Ackah, L.M.; Dubois, A. Treatment of Helicobacter pylori infection in rhesus monkeys using a novel antiadhesion compound. Gastroenterology 1999, 117, 1316–1325. [Google Scholar] [CrossRef]
- Parente, F.; Cucino, C.; Anderloni, A.; Grandinetti, G.; Bianchi Porro, G. Treatment of Helicobacter pylori infection using a novel antiadhesion compound (3'sialyllactose sodium salt). A double blind, placebo-controlled clinical study. Helicobacter 2003, 8, 252–156. [Google Scholar] [CrossRef]
- Niehues, A.; Euler , M.; Georgi, G.; Mank, M.; Stahl, B.; Hensel, A. Peptides from Pisum sativum L. enzymatic protein digest with anti-adhesive activity against Helicobacter pylori: Structure-activity and inhibitory activity against BabA, SabA, HpaA and a fibronectin-binding adhesion. Mol. Nutr. Food Res. 2010, 54, 1–11. [Google Scholar]
- Wittschier, N.; Faller, G.; Hensel, A. An extract of Pelargonium sidoides (EPs 7630) inhibits in situ adhesion of Helicobacter pylori to human stomach. Phytomedicine 2007, 14, 285–288. [Google Scholar] [CrossRef]
- Shmuely, H.; Yahav, J.; Samra, Z.; Niv, Y.; Sharon, N.; Weiss, E.; Athamna, A.; Tabak, M.; Ofek, I. Susceptibility of Helicobacter pylori isolates to the antiadhesion activity of a high-molecular-weight constituent of cranberry. Diagn. Microbiol. Infect. Dis. 2004, 50, 231–235. [Google Scholar] [CrossRef]
- Hofmann, T.; Deters, A.; Müller, G.; Stark, T.; Wittschier, N.; Hensel, A. Occurence of N-Phenylpropenoyl-L-amino acids in different herbal drugs and influence on human keratinocytes, human liver cells and against adhesion of H. pylori to human stomach. Planta Med. 2007, 73, 142–150. [Google Scholar] [CrossRef]
- Wittschier, N.; Faller, G.; Hensel, A. Aqueous extracts and polysaccharides from Liquorice roots (Glycyrrhiza glabra L.) inhibit adhesion of Helicobacter pylori to human gastric mucosa. J. Ethnopharmacol. 2009, 125, 218–223. [Google Scholar] [CrossRef]
- Lengsfeld, C.; Titgemeyer, F.; Faller, G.; Hensel, A. Glycosylated compounds from Okra inhibit adhesion of Helicobacter pylori to human gastric mucosa. J. Agric. Food Chem. 2004, 52, 1495–1503. [Google Scholar] [CrossRef]
- Messing, J.; Thöle, C.; Niehues, M.; Shevtsova, A.; Glocker, E.; Borén, T.; Hensel, A. Antiadhesive properties of Abelmoschus esculentus (Okra) immature fruit extract against Helicobacter pylori ddhesion. PLoS One 2014, 9, e84836. [Google Scholar]
- Ilver, D.; Arnqvist, A.; Ögren, J.; Frick, I.M.; Kersulyte, D.; Incecik, E.T.; Berg, D.E.; Covacci, A.; Engstrand, L.; Borén, T. Helicobacter pylori adhesin binding fucosylated histo-blood group antigens revealed by retagging. Science 1998, 279, 373–377. [Google Scholar] [CrossRef]
- Khamri, W.; Moran, A.P.; Worku, M.L.; Karimm, Q.N.; Walker, M.M.; Annuk, H.; Ferris, J.A.; Appelmelk, B.J.; Eggleton, P.; Reid, K.B.; et al. Variations in Helicobacter pylori lipopolysaccharide to evade the innate immune component surfactant protein D. Infect. Immun. 2005, 73, 7677–7686. [Google Scholar] [CrossRef]
- Van De Bovenkamp, J.H.; Mahdavi, J.; Korteland-Van Male, A.M.; Büller, H.A.; Einerhand, A.W. The MUC5AC glycoprotein is the primary receptor for Helicobacter pylori in the human stomach. Helicobacter 2003, 8, 521–532. [Google Scholar] [CrossRef]
- Mahdavi, J.; Hurtig, M.; Olfat, F.O.; Forsberg, L.; Roche, N.; Angstrom, J.; Larsson, T.; Teneberg, S.; Karlsson, K.A.; Altraja, S.; et al. Helicobacter pylori SabA adhesin in persistent infection and chronic inflammation. Science 2002, 297, 573–578. [Google Scholar] [CrossRef]
- Aspholm, M.; Olfat, F.O.; Nordén, J.; Sondén, B.; Lundberg, C.; Sjöström, R.; Altraja, S.; Odenbreit, S.; Haas, R.; Wadström, T.; et al. SabA is the Helicobacter pylori hemagglutinin and is polymorphic in binding to sialylated glycans. PLoS Pathog. 2006, 2, e110. [Google Scholar] [CrossRef]
- Simon, P.M.; Goode, P.L.; Mobasseri, A.; Zopf, D. Inhibition of Helicobacter pylori binding to gastrointestinal epithelial cells by sialic acid-containing oligosaccharides. Infect. Immun. 1997, 65, 750–757. [Google Scholar]
- Odenbreit, S.; Faller, G.; Haas, R. Role of the alpAB proteins and lipopolysaccharide in adheion of Helicobacter pylori to human gastric tissue. Int. J. Med. Microbiol. 2002, 292, 247–256. [Google Scholar] [CrossRef]
- Odenbreit, S.; Till, M.; Hofreuter, M.D.; HFaller, G.; Haas, R. Genetic and functional characterisation of the alpAB gene locus essential for bacterial adherence of Helicobacter pylori to human gastric tissue. Mol. Microbiol. 1999, 31, 1537–1548. [Google Scholar] [CrossRef]
- Yamaoka, Y.; Alm, R.A. Helicobacter outer membrane proteins. In Helicobacter pylori: Molecular Genetics and Cellular Biology Helicobacter pylori Outer Membrane Proteins, 1st ed.; Yamaoka, Y., Ed.; Caister Academic Press: Norfolk, UK, 2008; pp. 37–60. [Google Scholar]
- Lengsfeld, C.; Faller, G.; Hensel, A. Okra polysaccharides inhibit adhesion of Campylobacter jejuni to mucosa isolated from poultry in vitro but not in vivo. Anim. Feed Sci. Technol. 2007, 135, 113–125. [Google Scholar] [CrossRef]
- Valkonen, K.H.; Ringner, M.; Ljungh, Å.; Wadström, T. High-affinity binding of laminin by Helicobacter pylori: Evidence for a lectin-like interaction. FEMS 1993, 7, 29–38. [Google Scholar]
- Valkonen, K.H.; Wadström, T.; Moran, A.P. Identification of the N-acetylneuraminyllactose-specific laminin-binding protein of Helicobacter pylori. Infect. Immun. 1997, 65, 916–923. [Google Scholar]
- Selgrad, M.; Malfertheiner, P. Treatment of Helicobacter pylori. Cur. Opin. Gastroenterol. 2011, 27, 565–570. [Google Scholar] [CrossRef]
- Van Holst, G.; Clarke, A.E. Quantification of arabinogalactan-protein in plant extracts by single radial gel diffusion. Anal. Biochem. 1995, 148, 446–450. [Google Scholar] [CrossRef]
- Gilbert, R.G.; Hess, M.; Jenkins, A.D.; Jones, R.G. Dispersity in polymer science (IUPAC recommendations 2009). Pure Appl. Chem. 2009, 81, 351–353. [Google Scholar]
- Blumenkranz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Sommer-Knudsen, J.; Clarke, A.E; Bacic, A. A galactose-rich, cell-wall glycoprotein from styles of Nicotiana alata. Plant J. 1996, 9, 71–83. [Google Scholar]
- Sehlbach, M.; König, S.; Mormann, M.; Sendker, J.D.; Hensel, A. The arabinogalactan protein cluster isolated from Jatropha curcas L seed embryo contains fasciclin, xylogen and LysM proteins. Carbohydr. Polym. 2013, 98, 522–531. [Google Scholar] [CrossRef]
- Hermann, A.; König, S.; Lechtenberg, M.; Sehlbach, M.; Vakrushev, S.; Peter-Katalinic, J.; Hensel, A. Proteoglycans from Boswellia serrata Roxb. And B. carteri Birdw. and identification of a proteolytic plant basic secretory protein. Glycobiology 2012, 22, 1424–1439. [Google Scholar] [CrossRef]
- Serpe, M.D.; Nothnagel, E.A. Fractionation and structural characterization of arabinogalactan-proteins from the cell wall of rose cells. Plant Physiol. 1995, 109, 1007–1016. [Google Scholar]
- Gleeson, P.A.; McNamara, M.; Wettenhall, R.E.H.; Stone, B.A.; Fincher, G.B. Characterization of the hydroxyproline-rich protein core of an arabinogalactan-protein secreted from suspension-cultured Lolium multiflorum (Italian ryegrass) endosperm cells. Biochem. J. 1989, 264, 857–862. [Google Scholar]
- Hunter, W.M.; Greenwood, F.C. Preparation of iodine-131 labelled human growth hormone of high specific activity. Nature 1962, 194, 495–496. [Google Scholar] [CrossRef]
- Aspholm-Hurtig, M.; Dailide, G.; Lahmann, M.; Kalia, A; Ilver, D.; Roche, N.; Vikström, S.; Sjöström, R.; Lindén, S.; Bäckström, A.; et al. Functional adaptation of BabA, the H. pylori ABO blood group antigen binding adhesin. Science 2004, 305, 519–522. [Google Scholar] [CrossRef]
- Sun, H.Y.; Lin, S.W.; Ko, T.P.; Pan, J.F.; Liu, C.L.; Lin, C.L.; Wang, A.H.; Lin, C.H. Structure and mechanism of Helicobacter pylori fucosyltransferase. A basis for lipopolysaccharide variation and inhibitor design. J. Biol. Chem. 2007, 282, 9973–9982. [Google Scholar]
- Stingl, K.; de Reuse, H. Staying alive overdosed: How does Helicobacter pylori control urease activity? Int. J. Med. Microbiol. 2005, 295, 305–315. [Google Scholar]
- Kieliszewski, M.J.; Lamport, D.T.A.; Tan, L.; Cannon, M.C. Hydroxyproline-rich glycoproteins: Form and Function. Annu. Plant Rev.: Plant Polysacch. 2011, 41, 321–342. [Google Scholar]
- Gisbert, J.P. The recurrence of Helicobacter pylori infection: Incidence and variables influencing it. A critical review. Am. J. Gastrenterol. 2005, 100, 2083–2099. [Google Scholar] [CrossRef]
- Niv, Y. H. pylori recurrence after successful eradication. World J. Gastroenterol. 2008, 14, 1477–1478. [Google Scholar] [CrossRef]
- Niehues, M.; Georgi, G.; Euler, M.; Mank, M.; Klapperich, M.; Hensel, A. Pea protein peptides with anti Helicobacter pylori activity. WO2011122937 A1, 6 October 2011. [Google Scholar]
- Rasko, D.A.; Sperandion, V. Anti-virulence strategies to combat bacteria-mediated disease. Nat. Rev. 2010, 4, 117–127. [Google Scholar]
- Hensel, A. Antiadhesive polysaccharides. University of Münster: Münster, Germany, 2012; unpublished results. [Google Scholar]
- Niehues, M.; Stark, T.; Keller, D.; Hofmann, T.; Hensel, A. Antiadhesion as a functional concept: Antiadhesive N-phenylpropenoyl-L-amino acid amides as specific inhibitors of the Helicobacter pylori BabA outer membrane protein: Structure-activity relationship and functional aspects. Mol. Nutr. Food Res. 2011, 55, 1104–1117. [Google Scholar] [CrossRef]
- Skoog, E.M.; Sjöling, A.; Navabi, N.; Holgersson, J.; Lundin, S.B.; Lindén, S.K. Human gastric mucins differently regulate Helicobacter pylori proliferation, gene expression and interactions with host cells. PLoS One 2012, 7, e36378. [Google Scholar]
- Radziejewska, I.; Leszczyńska, K.; Borzym-Kluczyk, M. Influence of monoclonal anti-Lewis b, anti-H type 1, and anti-sialyl Lewis x antibodies on binding of Helicobacter pylori to MUC1 mucin. Mol. Cell Biochem. 2014, 385, 249–255. [Google Scholar] [CrossRef]
- Kalia, V.C.; Wood, T.K.; Kumar, P. Evolution of Resistance to Quorum-Sensing Inhibitors. Microb. Ecol. 2014. In press. [Google Scholar]
- Mellbye, B.; Schuster, M. The sociomicrobiology of antivirulence drug resistance: A proof of concept. mBio 2011, 2, e00131-11. [Google Scholar]
- Maeda, T.; García-Contreras, R.; Pu, M.; Sheng, L.; Garcia, L.R.; Tomás, M.; Wood, T.K. Quorum quenching quandary: Resistance to antivirulence compounds. ISME J. 2012, 6, 493–501. [Google Scholar] [CrossRef]
- Sharon, N. Safe as mother’s milk: Carbohydrates as future anti-adhesion drugs for bacterial diseases. Glycoconjugate J. 2000, 17, 659–664. [Google Scholar] [CrossRef]
- Ilver, D.; Arnqvist, A.; Ögren, J.; Frick, I.; Kersulyte, D. Helicobacter pylori adhesin binding fucosylated histo-blood group antigens revealed by retagging. Science 1989, 279, 5349–5373. [Google Scholar]
- Dubois, A.; Berg, D.E.; Incecik, E.T.; Fiala, N.; Heman-Ackah, L.M.; Del Valle, J.; Yang, M.; Wirth, H.P.; Perez-Perez, G.I.; Blaser, M.J. Host specificity of Helicobacter pylori strains and host responses in experimentally challenged nonhuman primates. Gastroenterology 1999, 116, 90–96. [Google Scholar] [CrossRef]
- Linden, S.; Nordman, H.; Hedenbro, J.; Hurtig, M.; Boren, T.; Carlstedt, I. Strain- and blood group-dependent binding of Helicobacter pylori to human gastric MUC5AC glycoforms. Gastroenterology 2002, 6, 1923–1930. [Google Scholar]
- Motulsky, H.; Christopoulos, A. Fitting Models TO Biological Data Using Linear and Nonlinear Regression: A Practical Guide To Curve Fitting; Oxford: New York, NY, USA, 2004; pp. 1–10. [Google Scholar]
- Sample Availability: Samples of F2 are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Messing, J.; Niehues, M.; Shevtsova, A.; Borén, T.; Hensel, A. Antiadhesive Properties of Arabinogalactan Protein from Ribes nigrum Seeds against Bacterial Adhesion of Helicobacter pylori. Molecules 2014, 19, 3696-3717. https://doi.org/10.3390/molecules19033696
Messing J, Niehues M, Shevtsova A, Borén T, Hensel A. Antiadhesive Properties of Arabinogalactan Protein from Ribes nigrum Seeds against Bacterial Adhesion of Helicobacter pylori. Molecules. 2014; 19(3):3696-3717. https://doi.org/10.3390/molecules19033696
Chicago/Turabian StyleMessing, Jutta, Michael Niehues, Anna Shevtsova, Thomas Borén, and Andreas Hensel. 2014. "Antiadhesive Properties of Arabinogalactan Protein from Ribes nigrum Seeds against Bacterial Adhesion of Helicobacter pylori" Molecules 19, no. 3: 3696-3717. https://doi.org/10.3390/molecules19033696