Shikonin Inhibits the Proliferation of Human Lens Epithelial Cells by Inducing Apoptosis through ROS and Caspase-Dependent Pathway

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
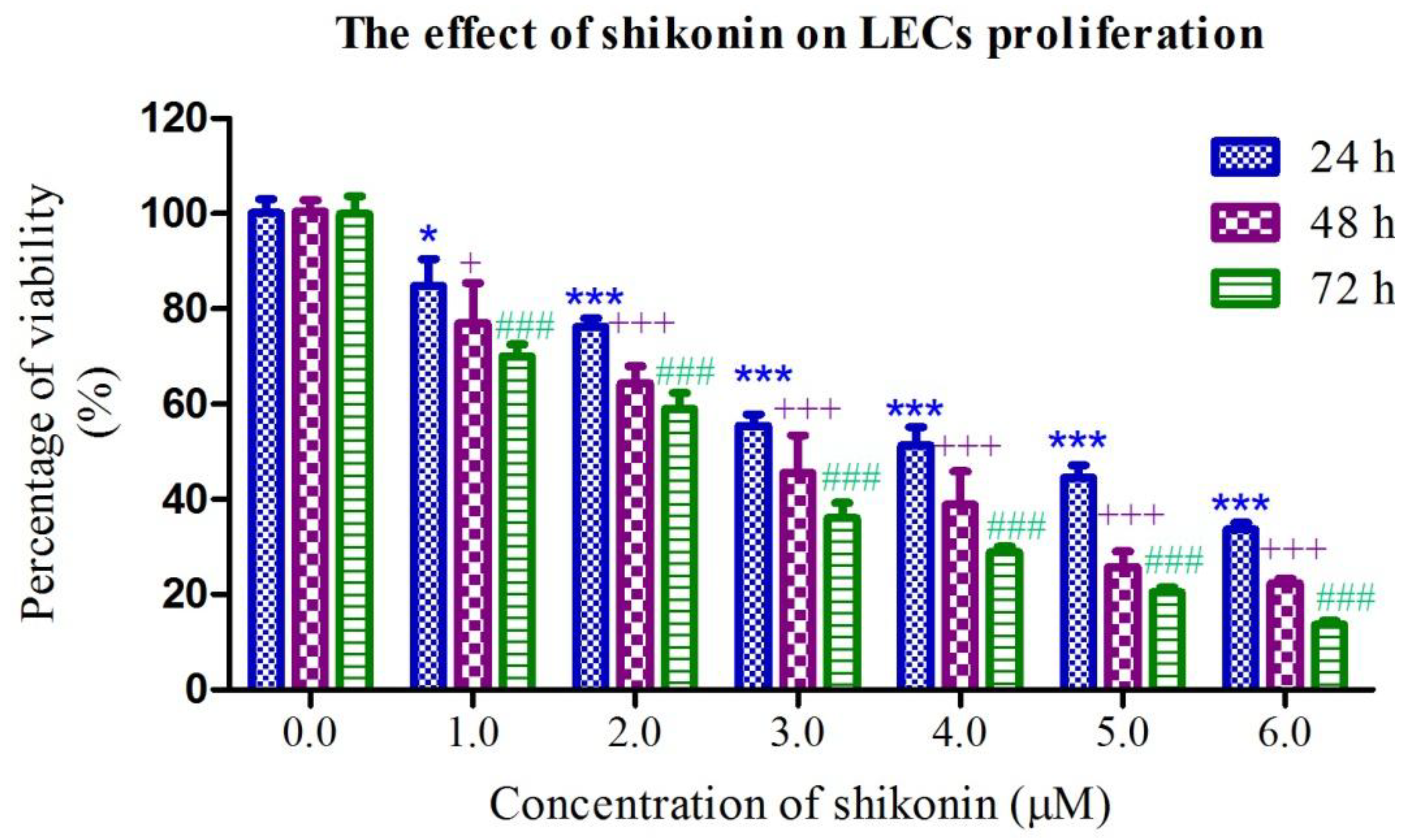
2.1. Shikonin Inhibited the Proliferation of HLEs

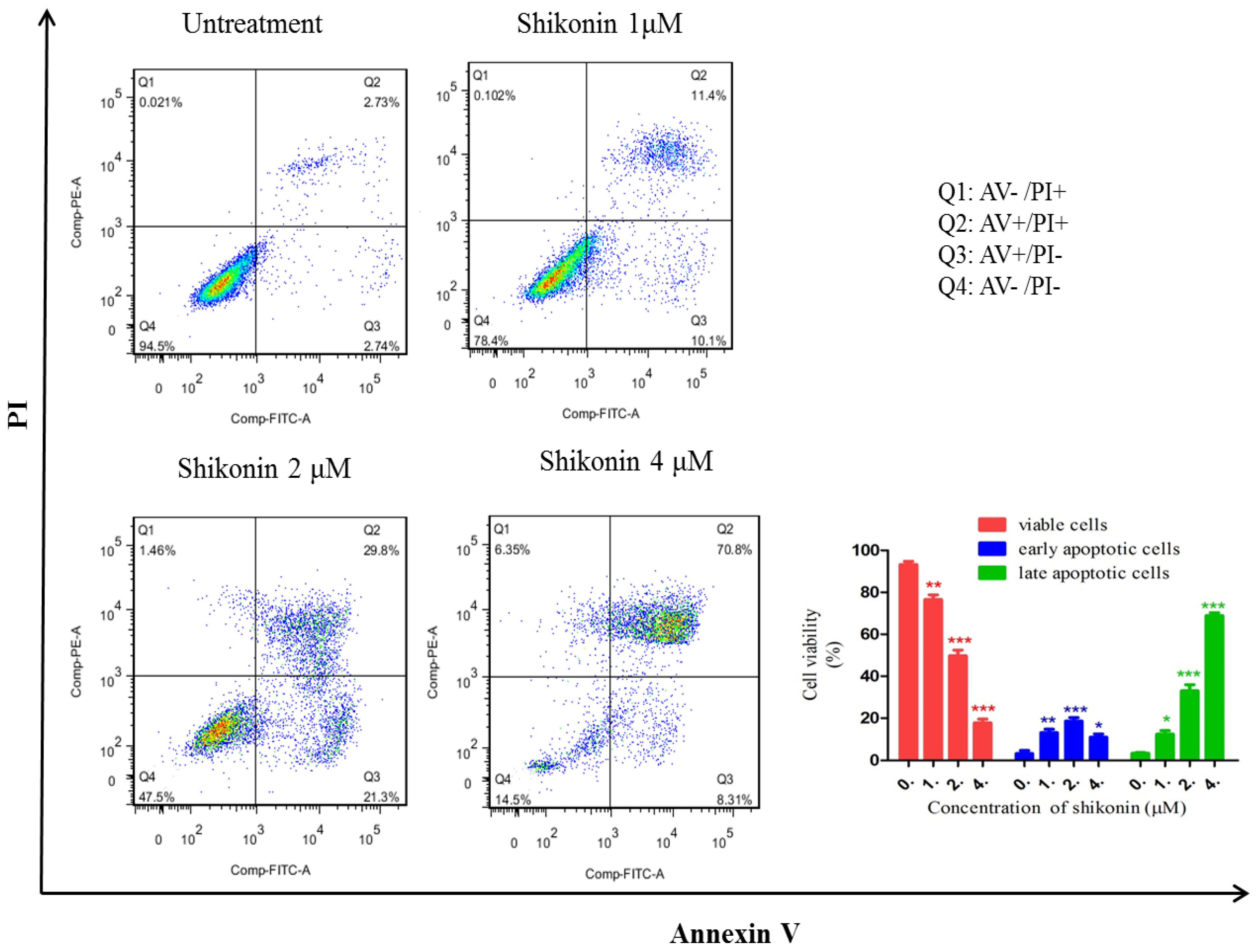
2.2. Shikonin Induced Apoptosis in HLEs
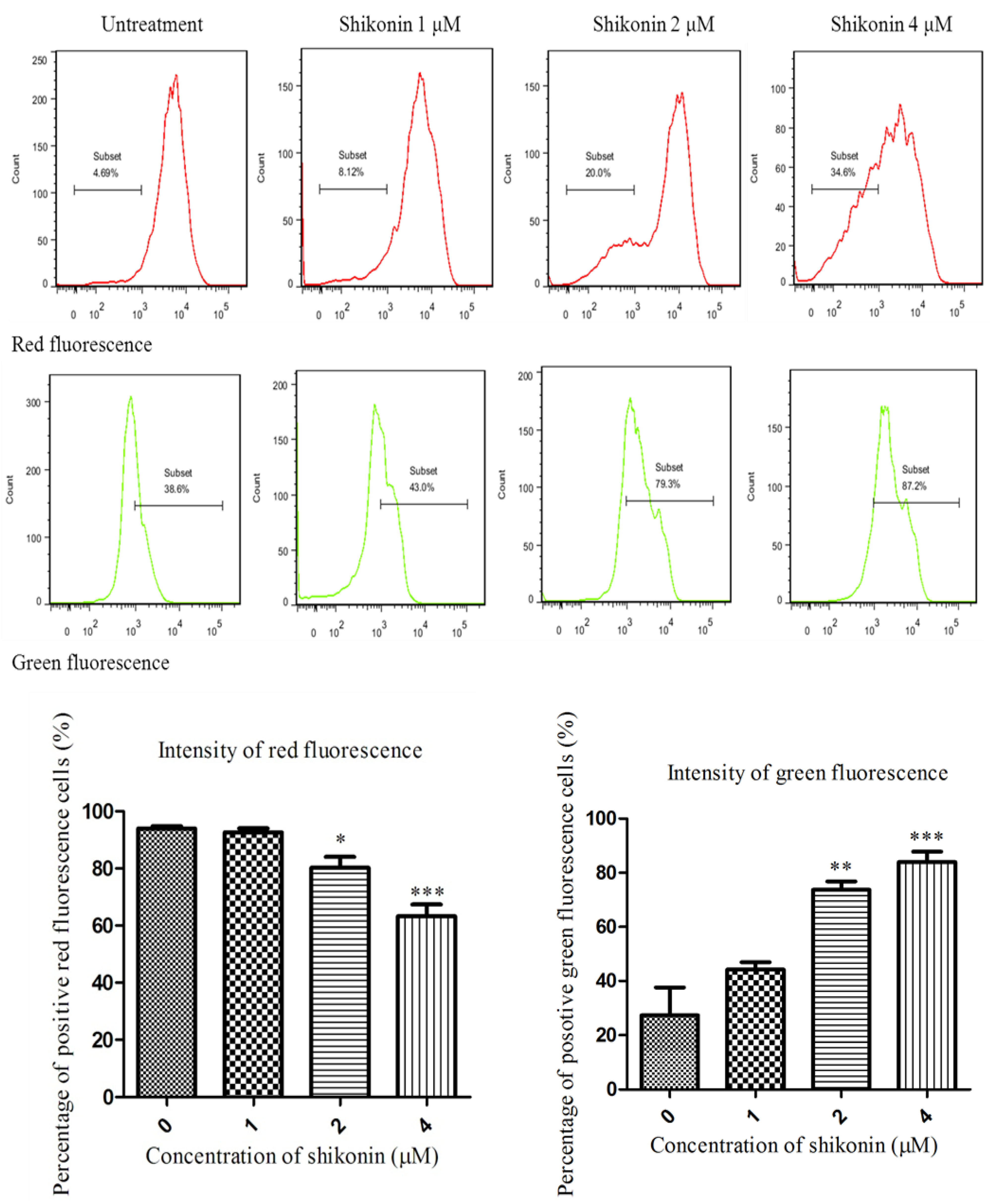
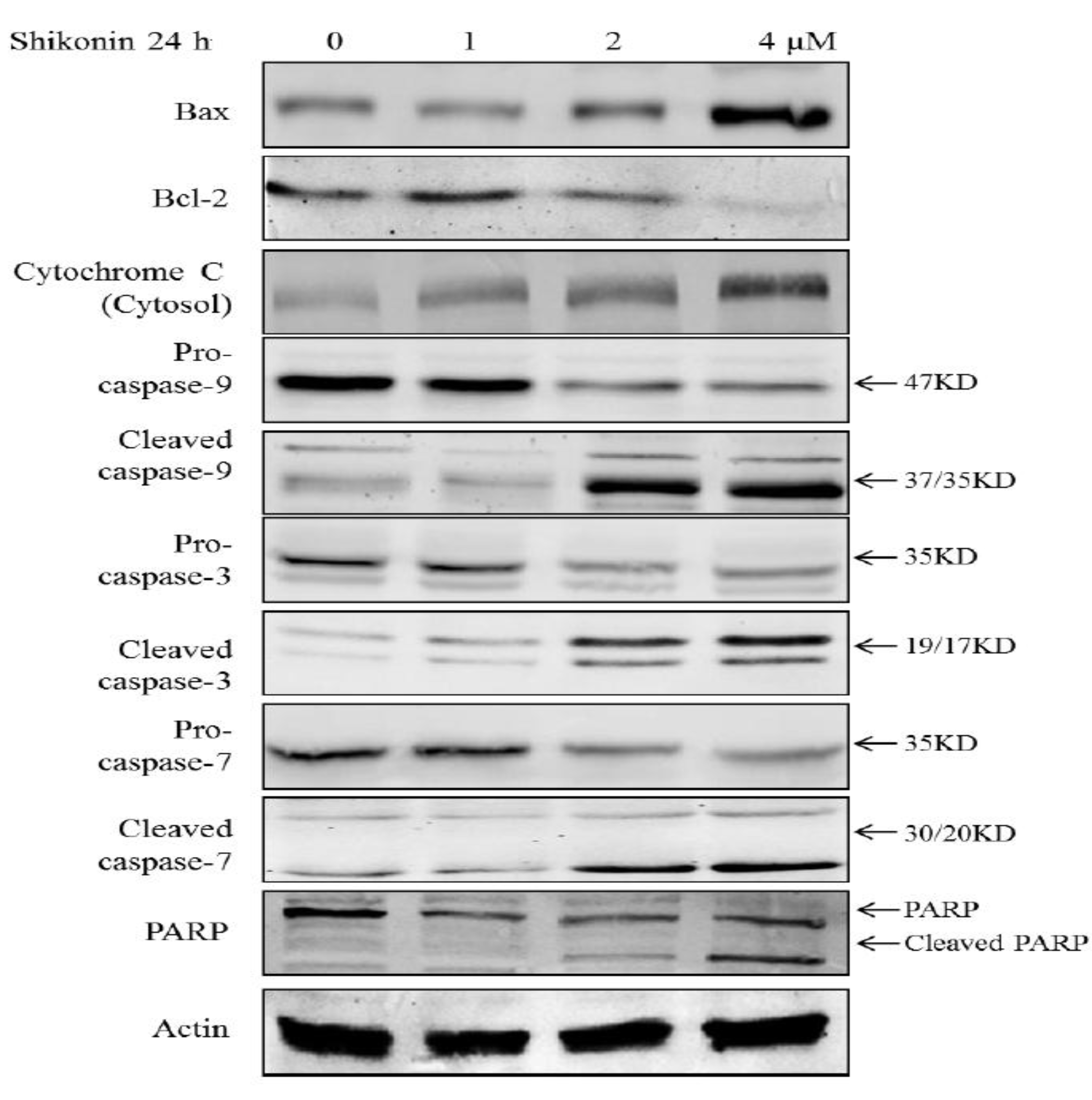
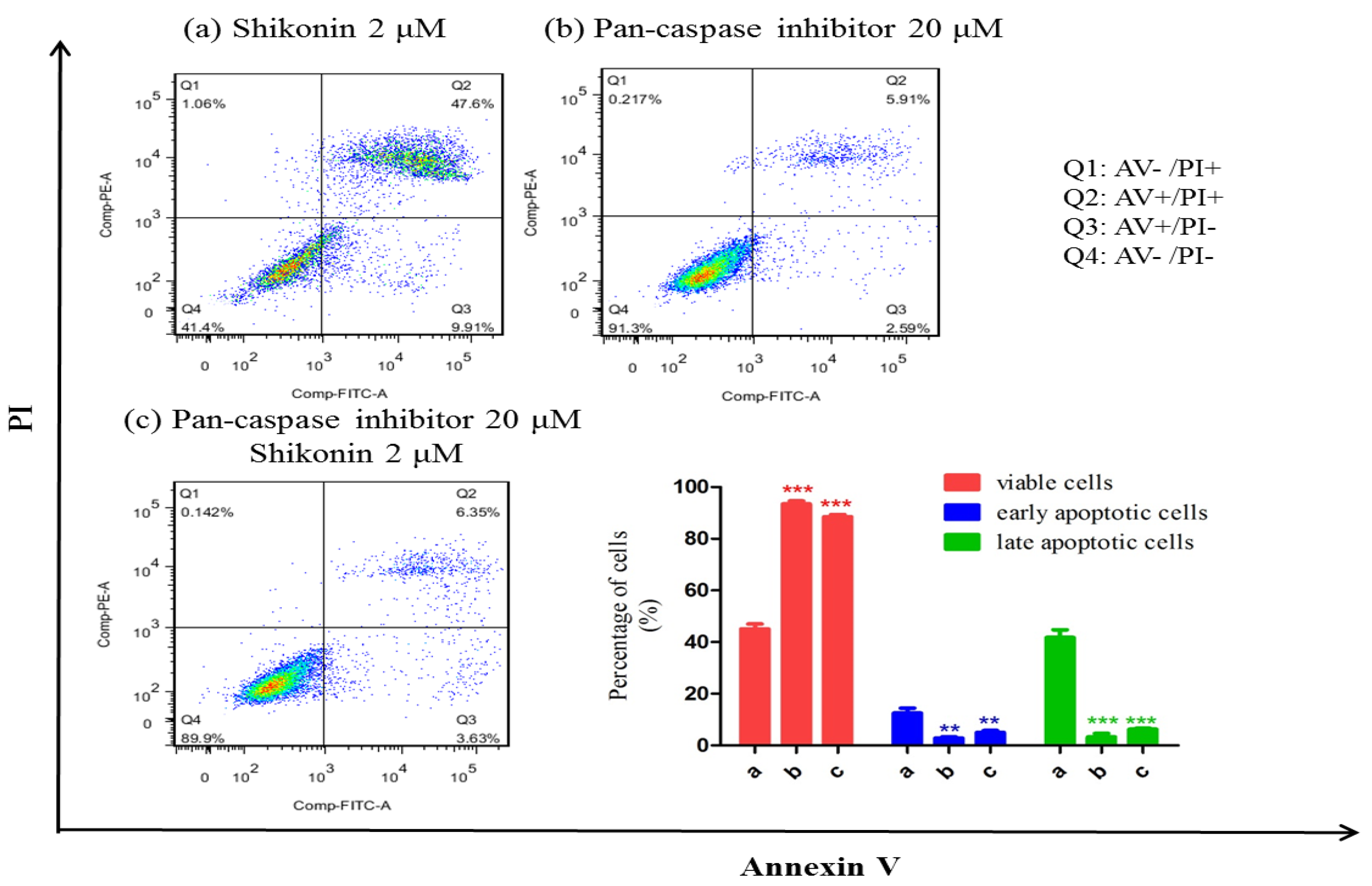
2.3. Shikonin Induced Apoptosis Through Mitochondrial and Caspases Dependent Pathway




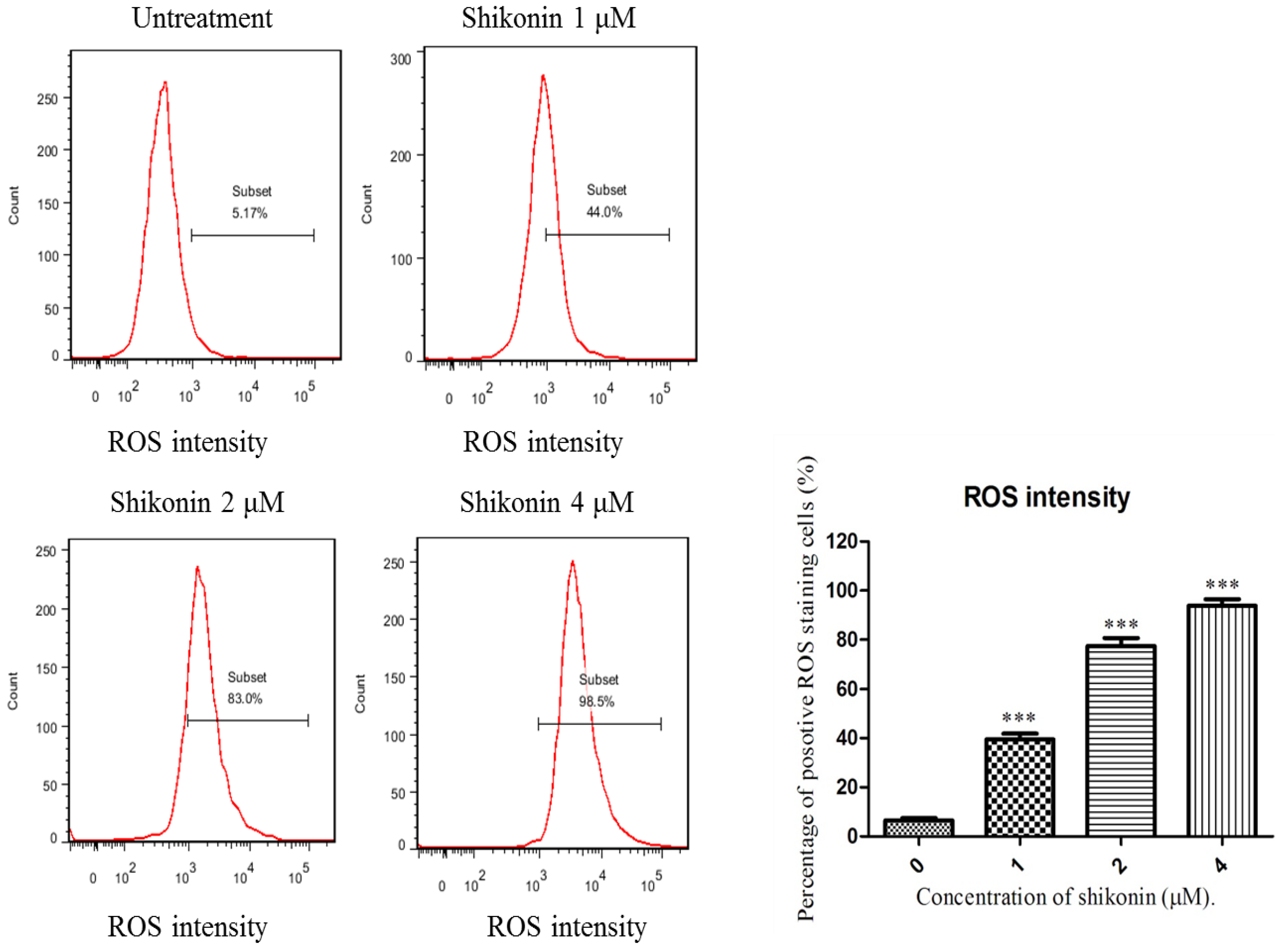
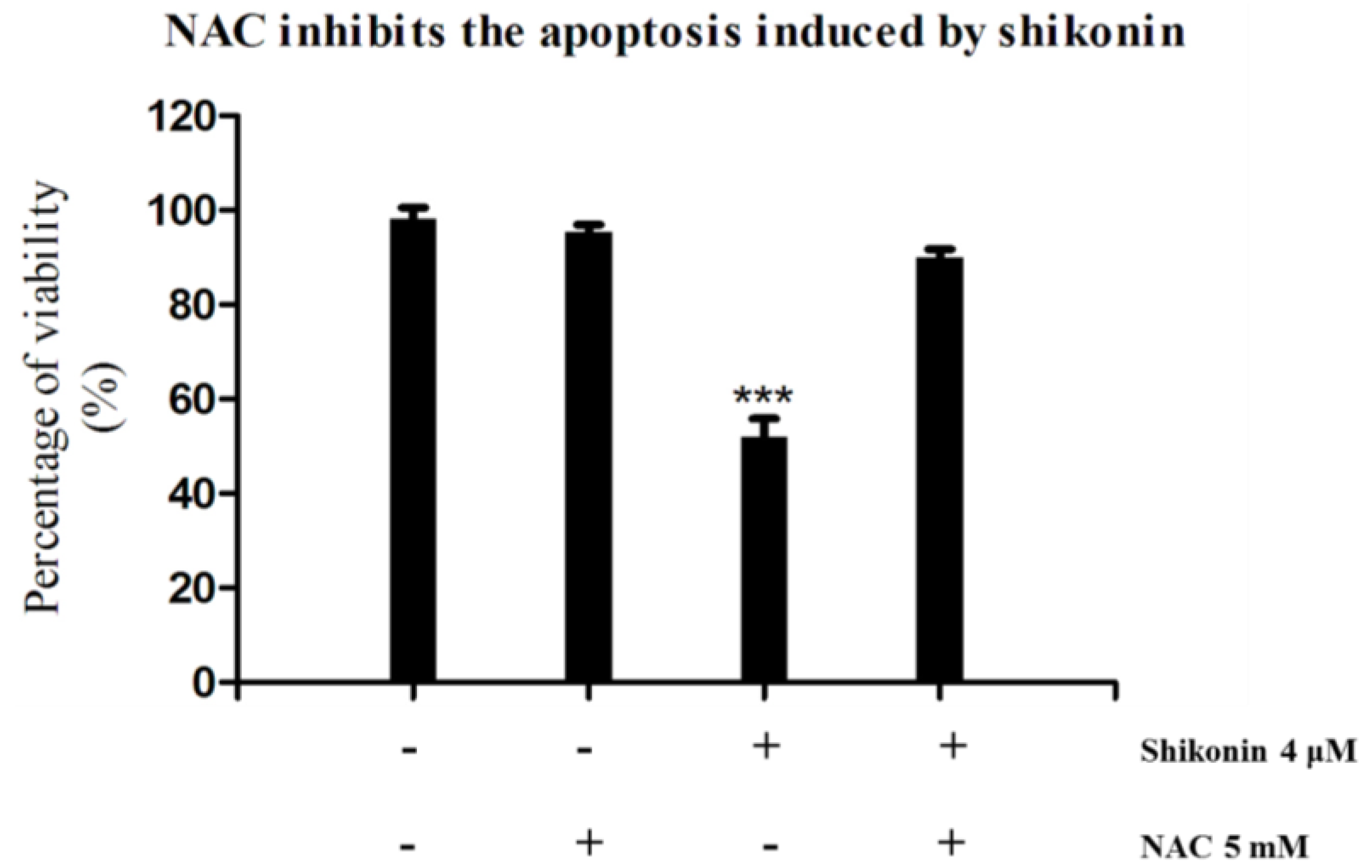
2.4. ROS Generation was Required for Shikonin-Induced Apoptosis in HLEs


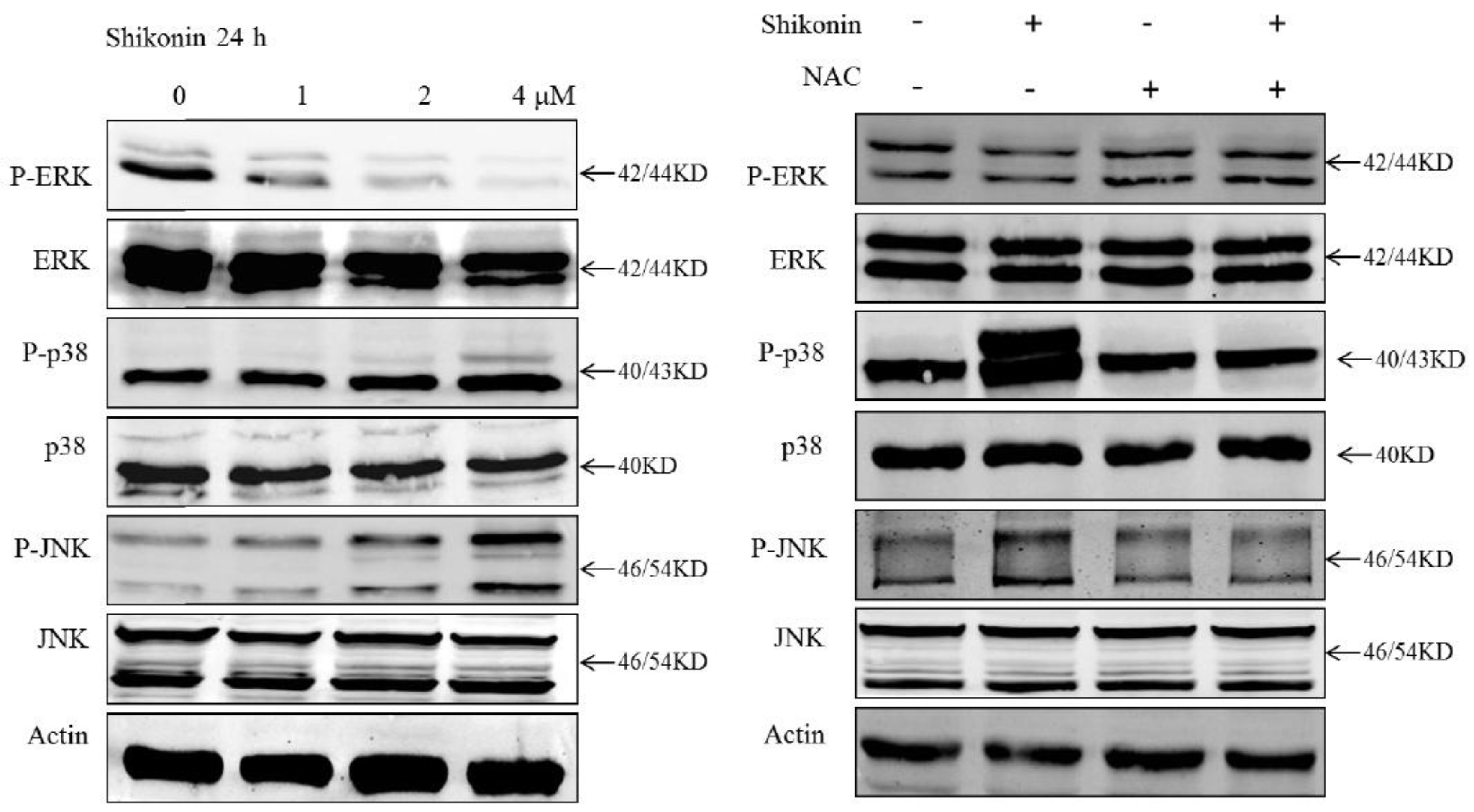
2.5. MAPK Pathway was Involved in Shikonin-Induced Apoptosis

3. Experimental Section
3.1. Chemicals
3.2. Cell Line and Cell Culture
3.3. 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium Bromide (MTT) Assay
3.4. Assessment of Apoptosis Levels by Annexin V/PI Staining
3.5. Analysis of Mitochondrial Membrane Potential (MMP)
3.6. Western Blot Analysis
3.7. Detection of ROS Generation
3.8. Statistical Analysis
4. Discussion and Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Meacock, W.R.; Spalton, D.J.; Boyce, J.; Marshall, J. The effect of posterior capsule opacification on visual function. Invest. Ophthalmol. Vis. Sci. 2003, 44, 4665–4669. [Google Scholar] [CrossRef]
- Fernandez, V.; Fragoso, M.A.; Billotte, C.; Lamar, P.; Orozco, M.A.; Dubovy, S.; Willcox, M.; Parel, J.M. Efficacy of various drugs in the prevention of posterior capsule opacification: Experimental study of rabbit eyes. J. Cataract Refract. Surg. 2004, 30, 2598–2605. [Google Scholar] [CrossRef]
- Spalton, D.J. Posterior capsular opacification after cataract surgery. Eye (Lond) 1999, 13, 489–492. [Google Scholar] [CrossRef]
- Billotte, C.; Berdeaux, G. Adverse clinical consequences of neodymium:YAG laser treatment of posterior capsule opacification. J. Cataract Refract. Surg. 2004, 30, 2064–2071. [Google Scholar] [CrossRef]
- Totan, Y.; Yagci, R.; Erdurmus, M.; Bayrak, R.; Hepsen, I.F. Cyclosporin effectively inhibits posterior capsule opacification after phacoemulsification in rabbits: A preliminary study. Clin. Exp. Ophthalmol. 2008, 36, 62–66. [Google Scholar] [CrossRef]
- Liu, H.; Wu, L.; Fu, S.; Hou, Y.; Liu, P.; Cui, H.; Liu, J.; Xing, L.; Zhang, X. Polylactide-glycoli acid and rapamycin coating intraocular lens prevent posterior capsular opacification in rabbit eyes. Graefes. Arch. Clin. Exp. Ophthalmol. 2009, 247, 801–807. [Google Scholar] [CrossRef]
- Legler, U.F.; Apple, D.J.; Assia, E.I.; Bluestein, E.C.; Castaneda, V.E.; Mowbray, S.L. Inhibition of posterior capsule opacification: The effect of colchicine in a sustained drug delivery system. J. Cataract Refract. Surg. 1993, 19, 462–470. [Google Scholar] [CrossRef]
- Malecaze, F.; Decha, A.; Serre, B.; Penary, M.; Duboue, M.; Berg, D.; Levade, T.; Lubsen, N.H.; Kremer, E.J.; Couderc, B. Prevention of posterior capsule opacification by the induction of therapeutic apoptosis of residual lens cells. Gene Ther. 2006, 13, 440–448. [Google Scholar] [CrossRef]
- Xu, Q.; Jia, R.B. bFGF siRNA expression plasmid inhibits growth of human LEC-B3 cells. Zhonghua Yan Ke Za Zhi 2008, 44, 1078–1082. [Google Scholar]
- Park, H.Y.; Kim, I.T.; Lee, K.M.; Choi, J.S.; Park, M.O.; Joo, C.K. Effects of nuclear factor-kappaB small interfering RNA on posterior capsule opacification. Invest. Ophthalmol. Vis. Sci. 2010, 51, 4707–4715. [Google Scholar] [CrossRef]
- Li, J.; Tang, X.; Chen, X. Comparative effects of TGF-beta2/Smad2 and TGF-beta2/Smad3 signaling pathways on proliferation, migration, and extracellular matrix production in a human lens cell line. Exp. Eye Res. 2011, 92, 173–179. [Google Scholar] [CrossRef]
- Awasthi, N.; Guo, S.; Wagner, B.J. Posterior capsular opacification: A problem reduced but not yet eradicated. Arch. Ophthalmol. 2009, 127, 555–562. [Google Scholar]
- Andujar, I.; Recio, M.C.; Giner, R.M.; Rios, J.L. Traditional chinese medicine remedy to jury: The pharmacological basis for the use of shikonin as an anticancer therapy. Curr. Med. Chem. 2013, 20, 2892–2898. [Google Scholar] [CrossRef]
- Yang, H.; Zhou, P.; Huang, H.; Chen, D.; Ma, N.; Cui, Q.C.; Shen, S.; Dong, W.; Zhang, X.; Lian, W.; et al. Shikonin exerts antitumor activity via proteasome inhibition and cell death induction in vitro and in vivo. Int. J. Cancer 2009, 124, 2450–2459. [Google Scholar] [CrossRef]
- Gong, K.; Li, W. Shikonin, a Chinese plant-derived naphthoquinone, induces apoptosis in hepatocellular carcinoma cells through reactive oxygen species: A potential new treatment for hepatocellular carcinoma. Free Radic. Biol. Med. 2011, 51, 2259–2271. [Google Scholar] [CrossRef]
- Mao, X.; Yu, C.R.; Li, W.H.; Li, W.X. Induction of apoptosis by shikonin through a ROS/JNK-mediated process in Bcr/Abl-positive chronic myelogenous leukemia (CML) cells. Cell Res. 2008, 18, 879–888. [Google Scholar] [CrossRef]
- Yeh, C.C.; Kuo, H.M.; Li, T.M.; Lin, J.P.; Yu, F.S.; Lu, H.F.; Chung, J.G.; Yang, J.S. Shikonin-induced apoptosis involves caspase-3 activity in a human bladder cancer cell line (T24). In Vivo 2007, 21, 1011–1019. [Google Scholar]
- Perelman, A.; Wachtel, C.; Cohen, M.; Haupt, S.; Shapiro, H.; Tzur, A. JC-1: Alternative excitation wavelengths facilitate mitochondrial membrane potential cytometry. Cell Death Disease 2012, 3, e430. [Google Scholar] [CrossRef]
- Huang, W.R.; Fan, X.X.; Tang, X. SiRNA targeting EGFR effectively prevents posterior capsular opacification after cataract surgery. Mol. Vis. 2011, 17, 2349–2355. [Google Scholar]
- Su, C.M.; Wang, S.W.; Lee, T.H.; Tzeng, W.P.; Hsiao, C.J.; Liu, S.C.; Tang, C.H. Trichodermin induces cell apoptosis through mitochondrial dysfunction and endoplasmic reticulum stress in human chondrosarcoma cells. Toxicol. Appl. Pharmacol. 2013, 272, 335–344. [Google Scholar] [CrossRef]
- Marcantonio, J.M.; Vrensen, G.F. Cell biology of posterior capsular opacification. Eye (Lond) 1999, 13, 484–488. [Google Scholar] [CrossRef]
- Inan, U.U.; Ozturk, F.; Kaynak, S.; Kurt, E.; Emiroglu, L.; Ozer, E.; Ilker, S.S.; Guler, C. Prevention of posterior capsule opacification by intraoperative single-dose pharmacologic agents. J. Cataract Refract. Surg. 2001, 27, 1079–1087. [Google Scholar]
- Xu, G.X.; Wang, T.T. Apoptosis of lens epithelial cells induced by cinobufagin in vitro. Int. J. Ophthalmol. 2010, 3, 128–131. [Google Scholar]
- Chen, X.; Yang, L.; Zhang, N.; Turpin, J.A.; Buckheit, R.W.; Osterling, C.; Oppenheim, J.J.; Howard, O.M. Shikonin, a component of chinese herbal medicine, inhibits chemokine receptor function and suppresses human immunodeficiency virus type 1. Antimicro. Agents Chemother. 2003, 47, 2810–2816. [Google Scholar] [CrossRef]
- Chan, D.C. Mitochondria: Dynamic organelles in disease, aging, and development. Cell 2006, 125, 1241–1252. [Google Scholar] [CrossRef]
- Cowan, K.J.; Storey, K.B. Mitogen-activated protein kinases: New signaling pathways functioning in cellular responses to environmental stress. J. Exp. Biol. 2003, 206, 1107–1115. [Google Scholar] [CrossRef]
- Andujar, I.; Recio, M.C.; Bacelli, T.; Giner, R.M.; Rios, J.L. Shikonin reduces oedema induced by phorbol ester by interfering with IkappaBalpha degradation thus inhibiting translocation of NF-kappaB to the nucleus. Br. J. Pharmacol. 2010, 160, 376–388. [Google Scholar] [CrossRef]
- Wada, T.; Penninger, J.M. Mitogen-activated protein kinases in apoptosis regulation. Oncogene 2004, 23, 2838–2849. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.
© 2014 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Huang, W.-R.; Zhang, Y.; Tang, X. Shikonin Inhibits the Proliferation of Human Lens Epithelial Cells by Inducing Apoptosis through ROS and Caspase-Dependent Pathway. Molecules 2014, 19, 7785-7797. https://doi.org/10.3390/molecules19067785
Huang W-R, Zhang Y, Tang X. Shikonin Inhibits the Proliferation of Human Lens Epithelial Cells by Inducing Apoptosis through ROS and Caspase-Dependent Pathway. Molecules. 2014; 19(6):7785-7797. https://doi.org/10.3390/molecules19067785
Chicago/Turabian StyleHuang, Wan-Rong, Yue Zhang, and Xin Tang. 2014. "Shikonin Inhibits the Proliferation of Human Lens Epithelial Cells by Inducing Apoptosis through ROS and Caspase-Dependent Pathway" Molecules 19, no. 6: 7785-7797. https://doi.org/10.3390/molecules19067785