2.1. The Effects of Nutritional Sources on Bioflocculant Production
The different carbon, nitrogen and cation sources evaluated for optimal utilization for bioflocculant production showed
Streptomyces sp. and
Brachybacterium sp. consortium to optimally utilize glucose, (NH
4)
2NO
3 and CaSO
4·H
2O respectively (
Table 1). The flocculation activities (in percentages) and bioflocculant yields (g/L) achieved by these carbon sources were: 56% and 2.78 ± 0.15 g/L (glucose), 51% and 2.52 ± 0.44 g/L (sucrose), 48% and 2.27 ± 0.18 g/L (fructose). Similarly, the nitrogen sources showed flocculation activities and bioflocculant yields of: 53% and 2.81 ± 0.37 g/L [(NH
4)
2NO
3], 49% and 1.96 ± 0.21 g/L (urea), 38% and 1.99 ± 0.56 g/L respectively (
Table 1). The cation sources with flocculation activities above 40% were CaSO
4·H
2O and MgCl
2 (
Table 1). Although glucose, [(NH
4)
2NO
3] and CaSO
4·H
2O were the preferred nutritional sources as they respectively yielded the optimal flocculation activity, the difference in flocculation activities achieved with other nutritional components, were not statistically significant (
p ≤ 0.05).
Table 1.
Nutritional sources optimally utilized by mixed culture of Brachybacterium sp. and Streptomyces sp. for the production of bioflocculant.
Table 1.
Nutritional sources optimally utilized by mixed culture of Brachybacterium sp. and Streptomyces sp. for the production of bioflocculant.
| Carbon Source | Glucose | Lactose | Fructose | Sucrose | Maltose | Starch | |
|---|
| MFA (%) | 56 | 42 | 48 | 51 | 33 | 46 | |
| BY (g/L) | 2.78 ± 0.15 | 2.34 ± 0.66 | 2.27 ± 0.18 | 2.52 ± 0.44 | 2.09 ± 0.61 | 1.99 ± 0.41 | |
| Nitrogen source | Urea | (NH4)2SO4 | (NH4)2NO3 | (NH4)2Cl4 | Peptone | | |
| MFA (%) | 49 | 36 | 53 | 38 | 42 | | |
| BY (g/L) | 1.96 ± 0.21 | 2.03 ± 0.26 | 2.81 ± 0.37 | 1.99 ± 0.56 | 2.31 ± 0.22 | | |
| Cation source | KCl | NaCl | MgCl2 | CaSO4·H2O | MnCl·4H2O | FeSO4 | FeCl3 |
| MFA (%) | 31 | 29 | 41 | 47 | 32 | 29 | 37 |
| BY (g/L) | 1.58 ± 0.11 | 1.26 ± 0.18 | 1.89 ± 0.21 | 2.19 ± 0.13 | 1.74 ± 0.17 | 1.55 ± 0.29 | 1.82 ± 0.41 |
Nonetheless, the consortium produced bioflocculant in an amount lower than the respective axenic cultures as noted by the yields and flocculation activity. A similar trend was observed with the assessed nitrogen and cation sources. Hence, yield optimization through mixed culture fermentation is achieved only when the respective culture acts in synergy [
24]. However,
Brachybacterium sp. UFH and
Streptomyces sp. Gansen seems to have acted in an antagonistic manner thus leading to the decline of bioflocculant yield and flocculation activity, respectively. The actinobacterial species responsible for the antagonistic effect is not known however, the decreased bioflocculant yield was taken as an indication of antagonism.
The utilization of various nutrient sources for the production of microbial secondary metabolites have been reported for axenic cultures [
3,
15,
25,
26], including the production of bioflocculants [
16,
17,
19]. However a dearth of information exists with respect to the use of mixed cultures in bioflocculant production, although mixed cultures effective in the degradation of environmental pollutants have been reported [
27], among other applications.
2.3. RSD Optimization of Critical Media Components for the Production of Bioflocculant
Glucose, (NH
4)
2NO
3 and CaSO
4·H
2O were next optimized in a 3-factor-5-level central composite design (
Table 4) following their emergence as critical media components in the PB design experimentation.
Table 4.
Central composite design matrix for critical media components showing the observed and predicted values for flocculation activity and bioflocculant yield.
Table 4.
Central composite design matrix for critical media components showing the observed and predicted values for flocculation activity and bioflocculant yield.
| Runs | Glucose | (NH4)2NO3 | CaSO4·H2O | Flocculation Activity (%) | Bioflocculant Yield (g/L) |
|---|
| Observed | Predicted | Observed | Predicted |
|---|
| 1 | 12.0(−1) | 0.5(−1) | 1.2(−1) | 52.5 | 53.68 | 2.53 | 2.48 |
| 2 | 12.0(−1) | 0.5(−1) | 1.6(+1) | 49.8 | 53.68 | 2.31 | 2.39 |
| 3 | 12.0(−1) | 1.5(+1) | 1.2(−1) | 58.1 | 56.12 | 2.92 | 2.74 |
| 4 | 12.0(−1) | 1.5(+1) | 1.6(+1) | 60.3 | 56.12 | 2.88 | 2.87 |
| 5 | 16.0(+1) | 0.5(−1) | 1.2(−1) | 63.7 | 55.53 | 3.02 | 2.93 |
| 6 | 16.0(+1) | 0.5(−1) | 1.6(+1) | 49.2 | 55.53 | 2.17 | 2.25 |
| 7 | 16.0(+1) | 1.5(+1) | 1.2(−1) | 61.0 | 57.97 | 2.92 | 2.74 |
| 8 | 16.0(+1) | 1.5(+1) | 1.6(+1) | 53.4 | 57.97 | 2.33 | 2.28 |
| 9 | 10.64(−1.73) | 1.0(0) | 1.4(0) | 51.2 | 54.27 | 2.46 | 2.51 |
| 10 | 17.36(+1.73) | 1.0(0) | 1.4(0) | 54.8 | 57.39 | 2.29 | 2.39 |
| 11 | 14.0(0) | 0.36(−1.73) | 1.4(0) | 58.6 | 54.26 | 2.61 | 2.54 |
| 12 | 14.0(0) | 1.74(+1.73) | 1.4(0) | 56.1 | 57.64 | 2.57 | 2.81 |
| 13 | 14.0(0) | 1.0(0) | 1.06(−1.73) | 53.3 | 55.83 | 2.48 | 2.74 |
| 14 | 14.0(0) | 1.0(0) | 1.84(+1.73) | 57.2 | 55.83 | 2.33 | 2.26 |
| 15 | 14.0(0) | 1.0(0) | 1.4(0) | 56.4 | 55.83 | 2.31 | 2.44 |
| 16 | 14.0(0) | 1.0(0) | 1.4(0) | 55.9 | 55.83 | 2.47 | 2.44 |
| 17 | 14.0(0) | 1.0(0) | 1.4(0) | 56.2 | 55.83 | 2.56 | 2.44 |
| 18 | 14.0(0) | 1.0(0) | 1.4(0) | 56.9 | 55.83 | 2.34 | 2.44 |
| 19 | 14.0(0) | 1.0(0) | 1.4(0) | 56.0 | 55.83 | 2.71 | 2.44 |
| 20 | 14.0(0) | 1.0(0) | 1.4(0) | 56.2 | 55.83 | 2.36 | 2.44 |
The respective proportion of critical media with the highest flocculation activities were 16.0 g/L, 0.5 g/L and 1.6 g/L of glucose, (NH4)2NO3 and CaSO4·H2O, respectively, following the twenty experimental trials shown in the 3-factor-5-level CCD matrix. The flocculation activity and bioflocculant yield achieved at this media components optimum were 63.7% and 3.02 g/L, respectively.
Following analysis of variance, the response surface was fitted to a second order model (
Table 5). The relatively high regression coefficient value obtained (R
2 = 0.6569), implied a 65.69% variability with respect to enhancing bioflocculant production as earmarked by the flocculation activity shown by
Brachybacterium sp. and
Streptomyces sp. consortium.
Table 5.
Analysis of variance showing fitted quadratic polynomial model for optimization of flocculation activity by Brachybacterium sp. and Streptomyces sp. consortium fermentation.
Table 5.
Analysis of variance showing fitted quadratic polynomial model for optimization of flocculation activity by Brachybacterium sp. and Streptomyces sp. consortium fermentation.
| Source | Flocculation Activity |
|---|
| DF | SS | MS | F-ratio | p-value | R2 |
|---|
| Regression model | 9 | 165.1615 | 18.3513 | 2.13 | 0.127576 | 0.656946 |
| Linear | 3 | 44.2462 | 14.7487 | 1.71 | 0.227695 | 0.175994 |
| Quadratic | 3 | 18.5453 | 6.1818 | 0.72 | 0.564227 | 0.073766 |
| Lin x Lin | 3 | 102.37 | 34.1233 | 3.96 | 0.042518 | 0.407187 |
| Total Error | 10 | 86.2466 | 8.6247 | | | 0.343054 |
| Lack of Fit | 5 | 85.6132 | 17.1226 | 135.18 | 0.000025 | 0.340535 |
| Pure Error | 5 | 0.63333 | 0.12667 | | | 0.002519 |
| Source | Bioflocculant Yield |
| DF | SS | MS | F-ratio | p-value | R2 |
| Regression model | 9 | 0.7881873 | 875.7637 | 2.43 | 0.091091 | 0.686541 |
| Linear | 3 | 0.3663863 | 0.1221288 | 3.39 | 0.061856 | 0.319137 |
| Quadratic | 3 | 0.122301 | 0.040767 | 1.13 | 0.382087 | 0.106529 |
| Lin x Lin | 3 | 0.2995 | 998.3333 | 2.77 | 0.096567 | 0.260876 |
| Total Error | 10 | 0.3598677 | 359.8677 | | | 0.313459 |
| Lack of Fit | 5 | 0.2403843 | 480.7687 | 2.01 | 0.230646 | 0.209384 |
| Pure Error | 5 | 0.1194833 | 238.9667 | | | 0.104075 |
On the same note, the F-test obtained from the regression analysis validates the result with probability value of 0.1276 and the coefficient for the lack-of-fit value (R
2 = 0.341) which was not statistically significant (
p ≤ 0.000025). Hence, there is an indication of the model adequacy for the prediction of enhanced flocculation activity following the assay conditions (
Table 5).
Similarly, the analysis of variance for the bioflocculant yield (
Table 5) shows a regression coefficient of R
2 = 0.6865 thus, an indication of 68.65% adequacy. The adequacy of this model has been shown in the harvesting of high-density cultures of
Scenedesmus sp. through flocculation [
28], activity optimization for composite bioflocculant and polyaluminum chloride [
29] and in bioflocculant production optimization by the axenic culture of
Halomonas sp.V3a’ [
15].
The levels of significance of the main effects of glucose, (NH
4)
2NO
3 and CaSO
4·H
2O to the production of bioflocculant were 1.8%, 91.3% and 28.9% respectively, as indicated by the linear model (
Table 6). However, following the quadratic model (NH
4)
2NO
3 and CaSO
4·H
2O showed positive contribution to bioflocculant yield while glucose did not. The negative regression coefficients shown by glucose following the quadratic polynomial model is an indication of the low impact shown by this carbon source towards enhancing bioflocculant yield during fermentation by the consortium. The interaction between glucose, (NH
4)
2NO
3 and CaSO
4·H
2O showed that (NH
4)
2NO
3 and CaSO
4·H
2O was significant while the rest was not, as their coefficients of estimate were negative (
Table 6).
Table 6.
Second order polynomial model following regression analysis of flocculation activity optimization for Brachybacterium sp. and Streptomyces sp. consortium.
Table 6.
Second order polynomial model following regression analysis of flocculation activity optimization for Brachybacterium sp. and Streptomyces sp. consortium.
| Parameter | Estimate | Standard Error | t-Value | p-Value |
|---|
| Intercept | −121.1504 | | | |
| Glucose | 18.66811 | 6.626398 | 2.82 | 0.018247 |
| (NH4)2NO3 | 2.500087 | 22.24044 | 0.11 | 0.912721 |
| CaSO4·H2O | 63.57429 | 56.72536 | 1.12 | 0.288592 |
| Glucose × Glucose | −0.2474871 | 0.1938682 | −1.28 | 0.230601 |
| (NH4)2NO3 × (NH4)2NO3 | 2.360935 | 4.069187 | 0.58 | 0.574621 |
| CaSO4·H2O × CaSO4·H2O | 3.926695 | 14.69922 | 0.27 | 0.794792 |
| Glucose × (NH4)2NO3 | −1.825 | 1.038307 | −1.76 | 0.109316 |
| Glucose × CaSO4·H2O | −6.75 | 2.595768 | −2.60 | 0.026474 |
| (NH4)2NO3 × CaSO4·H2O | 14.75 | 10.38307 | 1.42 | 0.185859 |
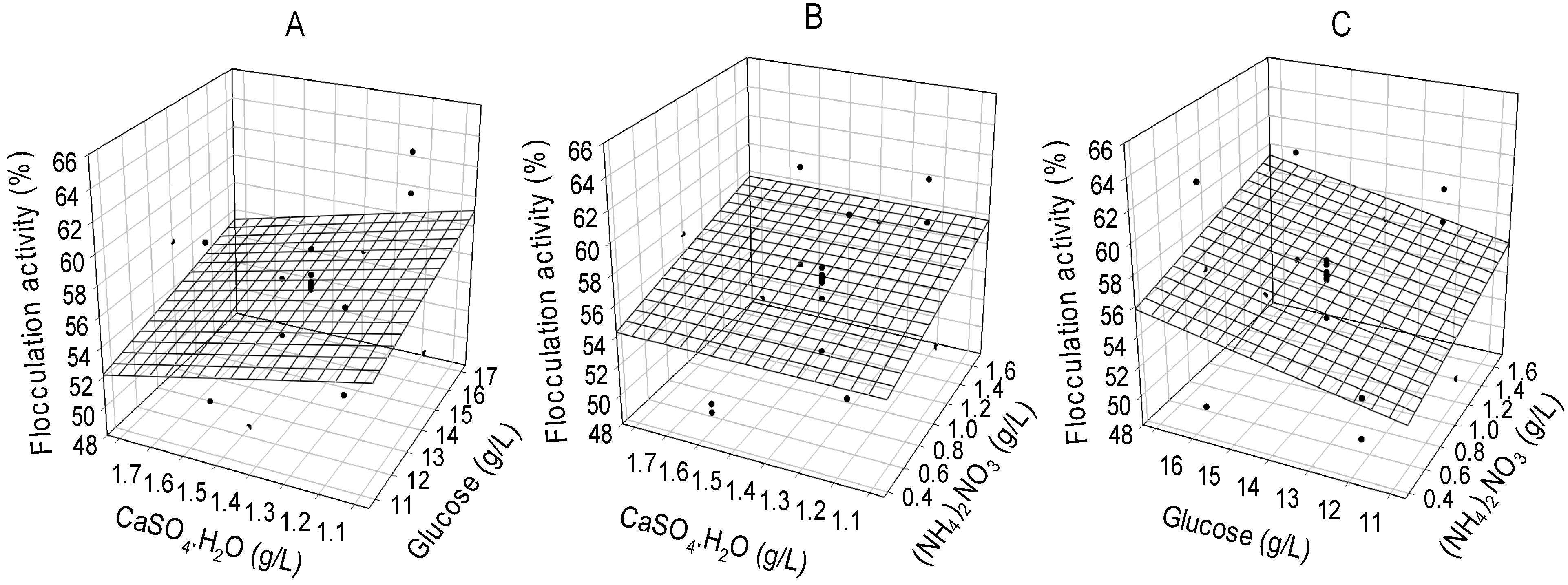
The three dimension surface response plot (
Figure 1) showing the concentrations of critical media components with response (flocculation activity) revealed that at a higher concentration of glucose and lower concentration of CaSO
4·H
2O, flocculation activity increased (
Figure 1A). Likewise, at higher concentrations of glucose and (NH
4)
2NO
3 flocculation activities increased (
Figure 1C) while the interaction between glucose and (NH
4)
2NO
3 apparently showed no increase in flocculation activity at any level (
Figure 1B). Consequently, the optimum ratio of the critical media components for the production of bioflocculant by the consortium of
Brachybacterium sp. and
Streptomyces sp. were: 16.0 g/L (glucose), 0.5 g/L [(NH
4)
2NO
3] and 1.2 g/L (CaSO
4·H
2O), respectively. The maximum flocculation activity and bioflocculant yield achieved were 63.7% and 3.02 g/L, respectively.
Figure 1.
Three dimensional representations of interactions of critical media components after flocculation activity optimization following application of surface response design.
Figure 1.
Three dimensional representations of interactions of critical media components after flocculation activity optimization following application of surface response design.
The feasibility of optimizing critical media components is grim if cultures are unable to grow effectively. However, since we did not ascertain the survival of the respective axenic culture in the consortium then, it will be prudent not state that growth was poor particularly as it has been shown that bioflocculant production occurs at the exponential phase of bacterial growth [
5,
30]. The central composite design revealed the optimum ratio of respective critical media components for bioflocculant production while the PB design showed the utmost contribution towards bioflocculant production to have come from (NH
4)
2NO
3 and no clear reason can be adduced to this observation as carbon sources are known to be the most relevant factors for microbial growth. On the other hand, it may be that nitrogen sources were more important in bioflocculant production. Microbial secondary metabolites including poly-γ-glutamic acid from
Bacillus subtilis RKY3 [
31], bioflocculants from axenic culture of
Halomonas sp. V3a’ [
15], biosurfactants production by probiotic bacteria [
32] and in the production of cold active protease by a psychrophilic bacteria belonging to the genus of Colwellia [
33] have been optimized through response models. Despite the fact that surface response methodology is known for adequacy in yield optimization (secondary metabolites) and enhancing the output of desired effects, it did not show adequacy in the mixed culture fermentation as a negative regression coefficient was achieved. This may still be attributed to the antagonistic effects of the biomolecules produced in the fermentation process.
2.4. Micrographic Imaging and Compositional Characteristics of the Purified Bioflocculant
Electron micrographic imaging of the purified bioflocculant showed loosely packed fluffy materials with irregular arrangement patterns (
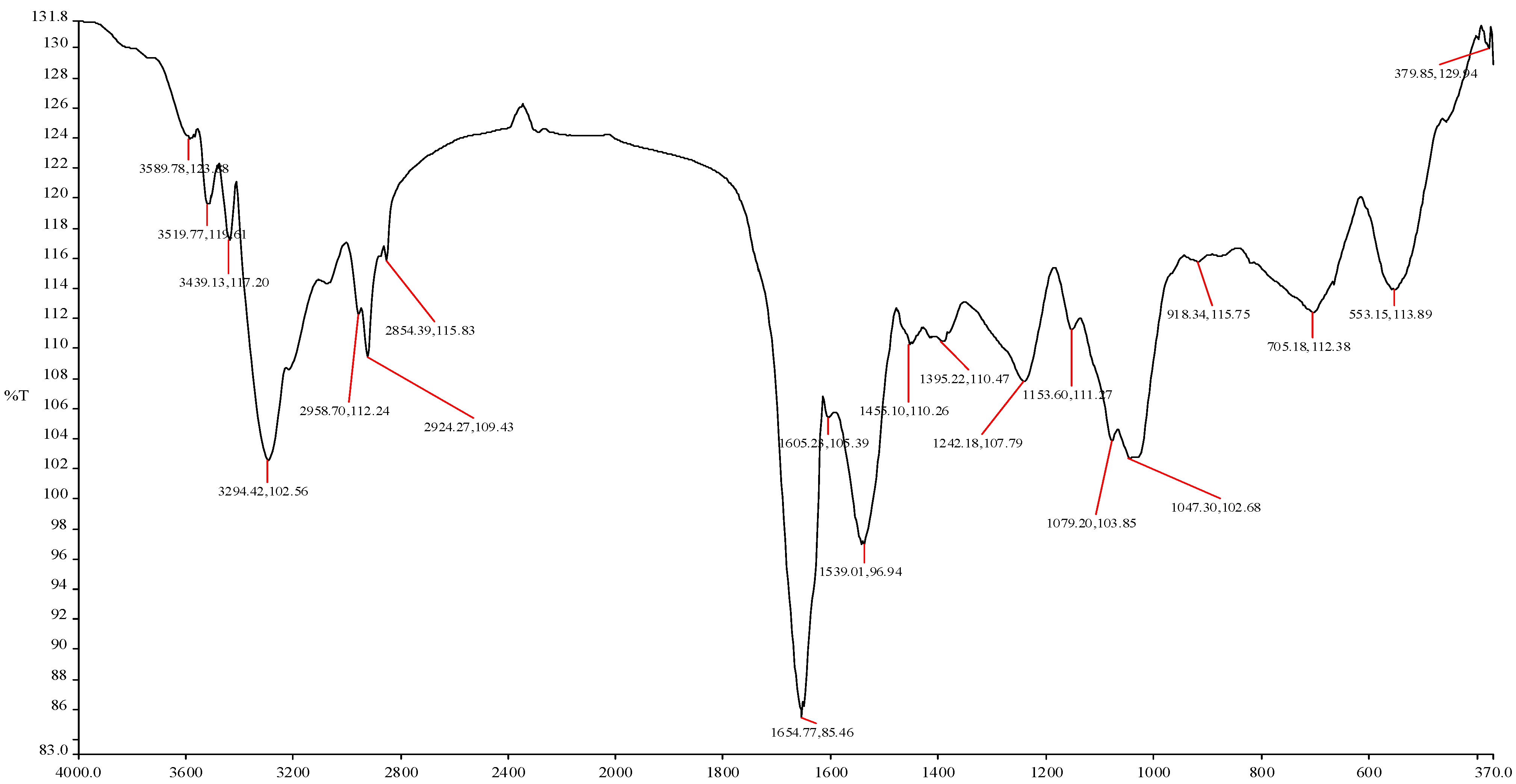
Figure 2). The interstices between the crispy flakes were less than 1 µm in size. The Fourier transform infrared spectrum (
Figure 3) of the purified bioflocculant showed broad stretching peaks at 3589.78 to 3294.42 (cm
−1), characteristic of hydroxyl groups from polymeric, dimeric and monomeric OH groups. Similarly, peaks from 2958.70 to 2854.39 cm
−1 correspond to weak C–H stretching bands from methylene groups, and those from 1654.77 to 1539.01 cm
−1 are indicative of the presence of aromatic rings [
5,
34,
35]. Furthermore, wave numbers 1455.10 to 1395.22 cm
−1 and 1242.18 to 1047.30 cm
−1 shown were typical of phenol and tertiary alcohol OH bend, indicative of the presence of carboxylic groups, carboxylate ions, aromatic ring stretch and C–O and C–O–C from polysaccharides [
34].
Figure 2.
Scan electron micrographic image of the purified bioflocculant produced by the consortium of Brachybacterium sp. and Streptomyces sp.
Figure 2.
Scan electron micrographic image of the purified bioflocculant produced by the consortium of Brachybacterium sp. and Streptomyces sp.
Figure 3.
FTIR spectrum of purified bioflocculant from mixed culture fermentation of Brachybacterium sp. UFH and Streptomyces sp. Gansen.
Figure 3.
FTIR spectrum of purified bioflocculant from mixed culture fermentation of Brachybacterium sp. UFH and Streptomyces sp. Gansen.
The loose amorphous fluffy nature of the bioflocculant is a marked variation from the clump-like nature shown by bioflocculants with high flocculation activity [
5]. Similarly, the micrographic image of the respective axenic cultures was more compact, hence it may be suggested that the formation of bioflocculant was adversely affected in the consortium. The loose nature may also be understood as weak bonding between the molecules constituting the bioflocculant which leads to the low flocculation activity observed. The various functional groups such as hydroxyl, benzylic, allylic, carboxyl, esters and amino groups, amongst others, shown by FTIR spectroscopy are suggestive of an amalgam of polymers containing uronic acids, carbohydrates, glycoproteins and proteins.



{kind=link}
{kind=link}
{kind=link}




