Effects of Tatariside G Isolated from Fagopyrum tataricum Roots on Apoptosis in Human Cervical Cancer HeLa Cells

Abstract
:1. Introduction

2. Results and Discussion
2.1. Effects of TG on HeLa Cells Viability

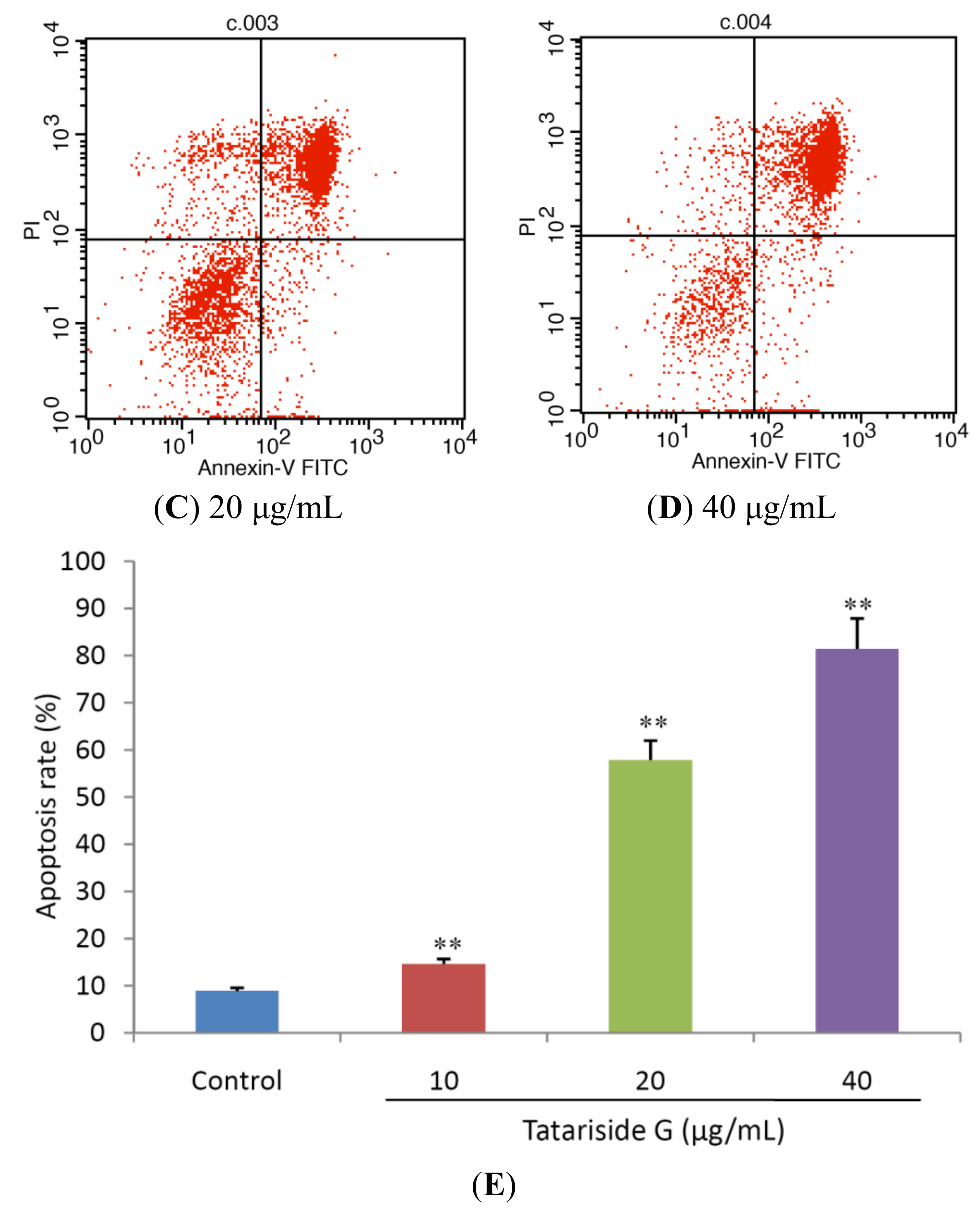
2.2. TG Induces Apoptosis of HeLa Cells



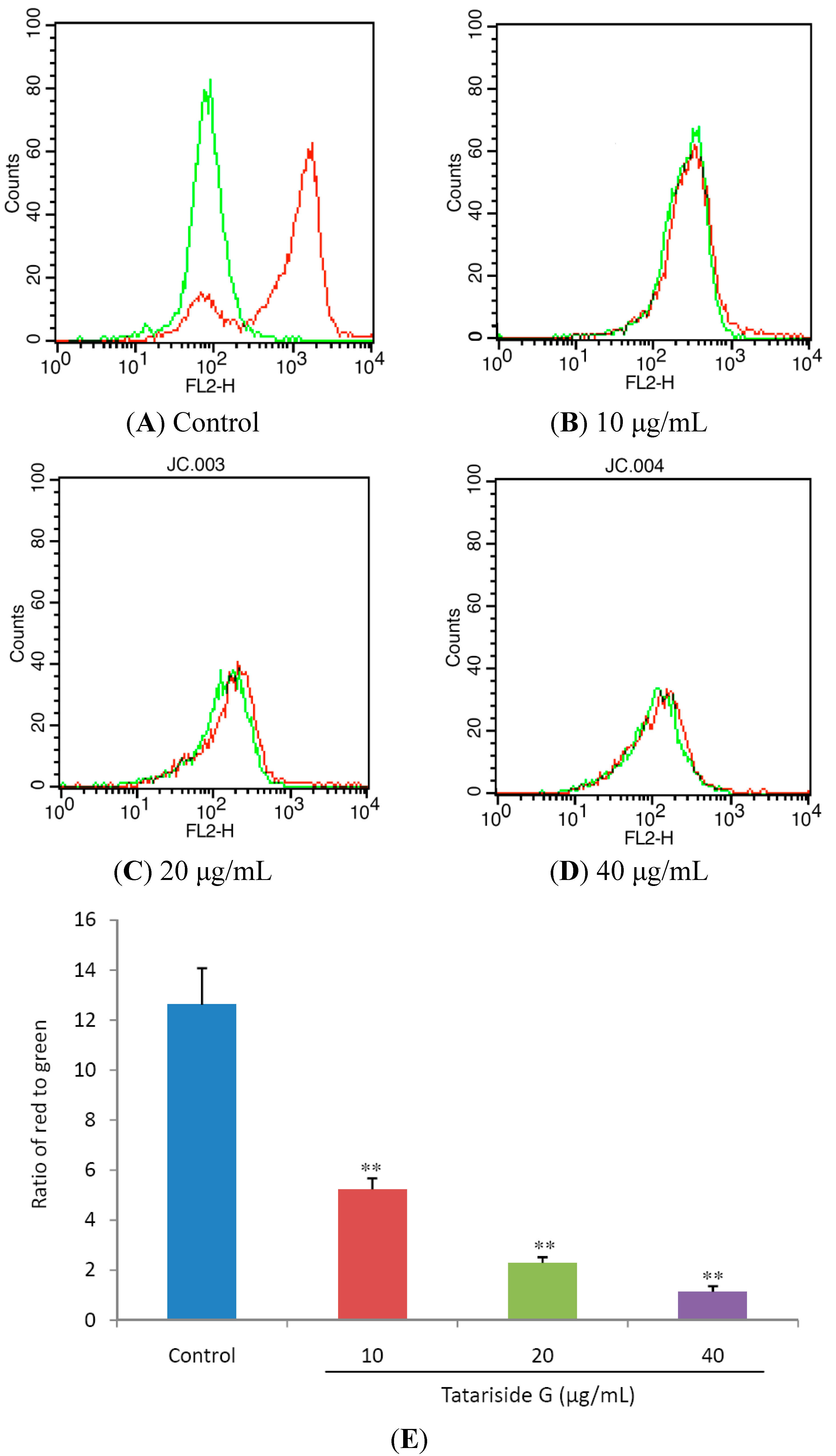
2.3. TG Decreases Mitochondrial Membrane Potential (MMP)

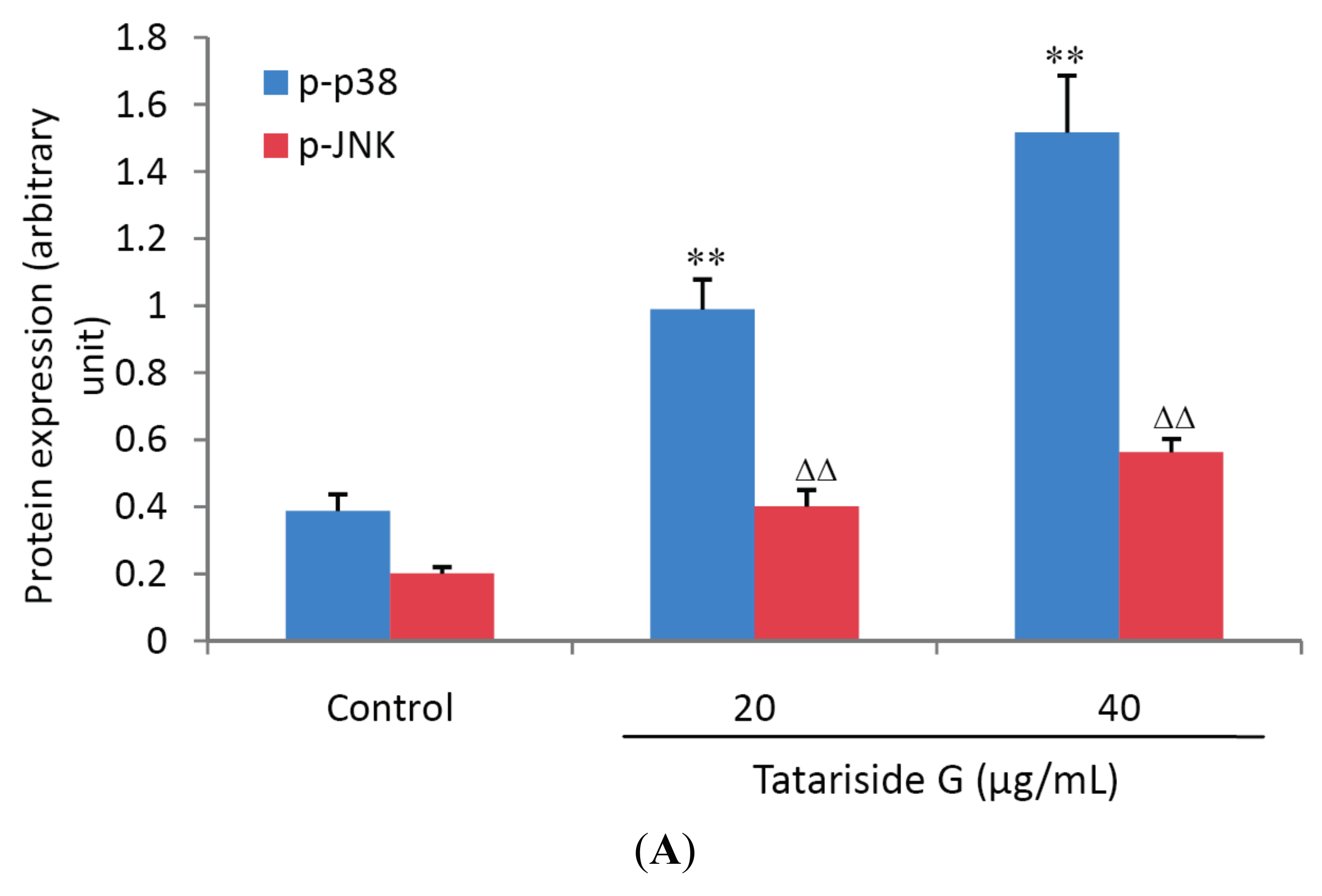
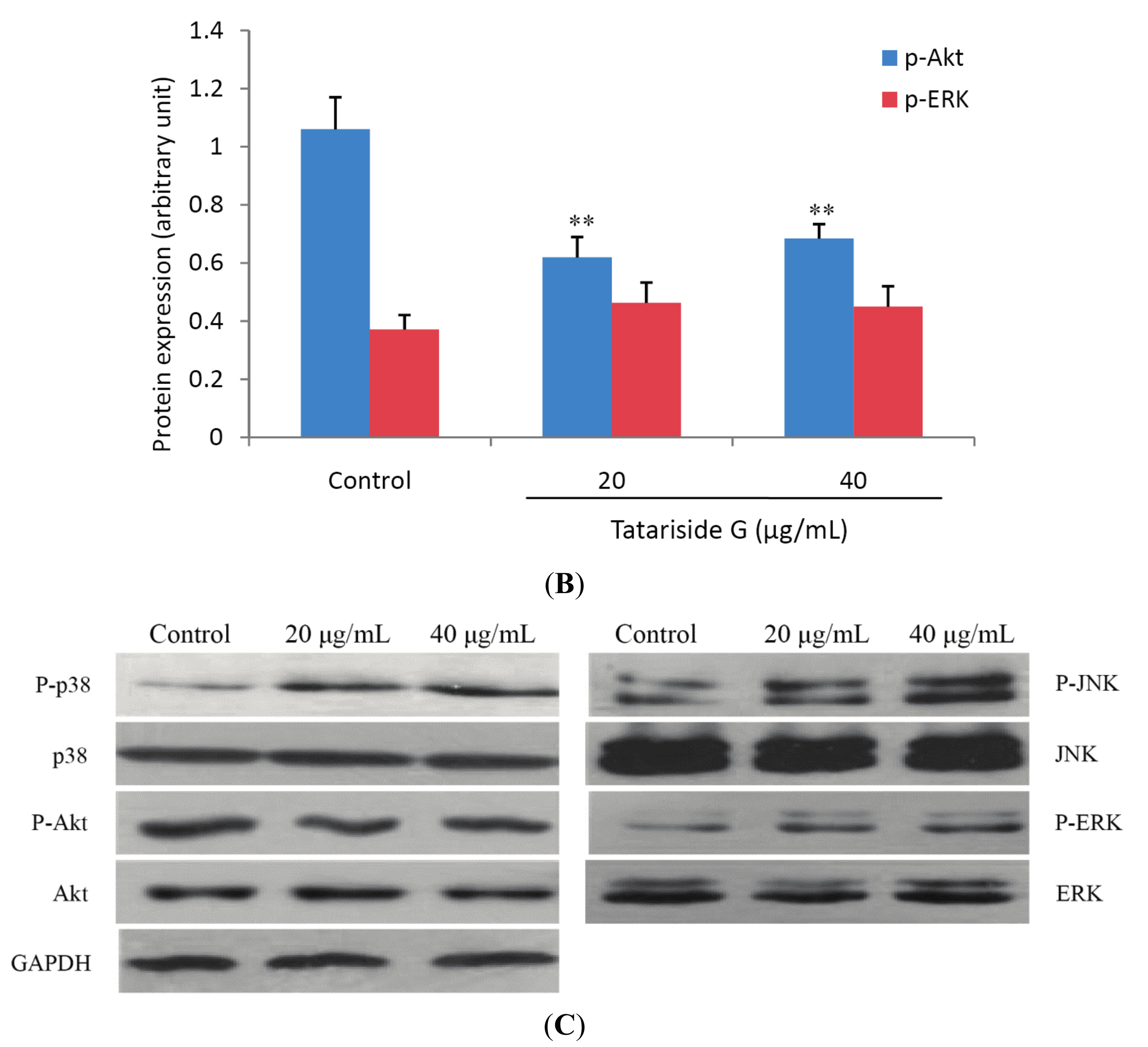
2.4. TG Regulates Protein Phosphorylation of p38, JNK, and Akt


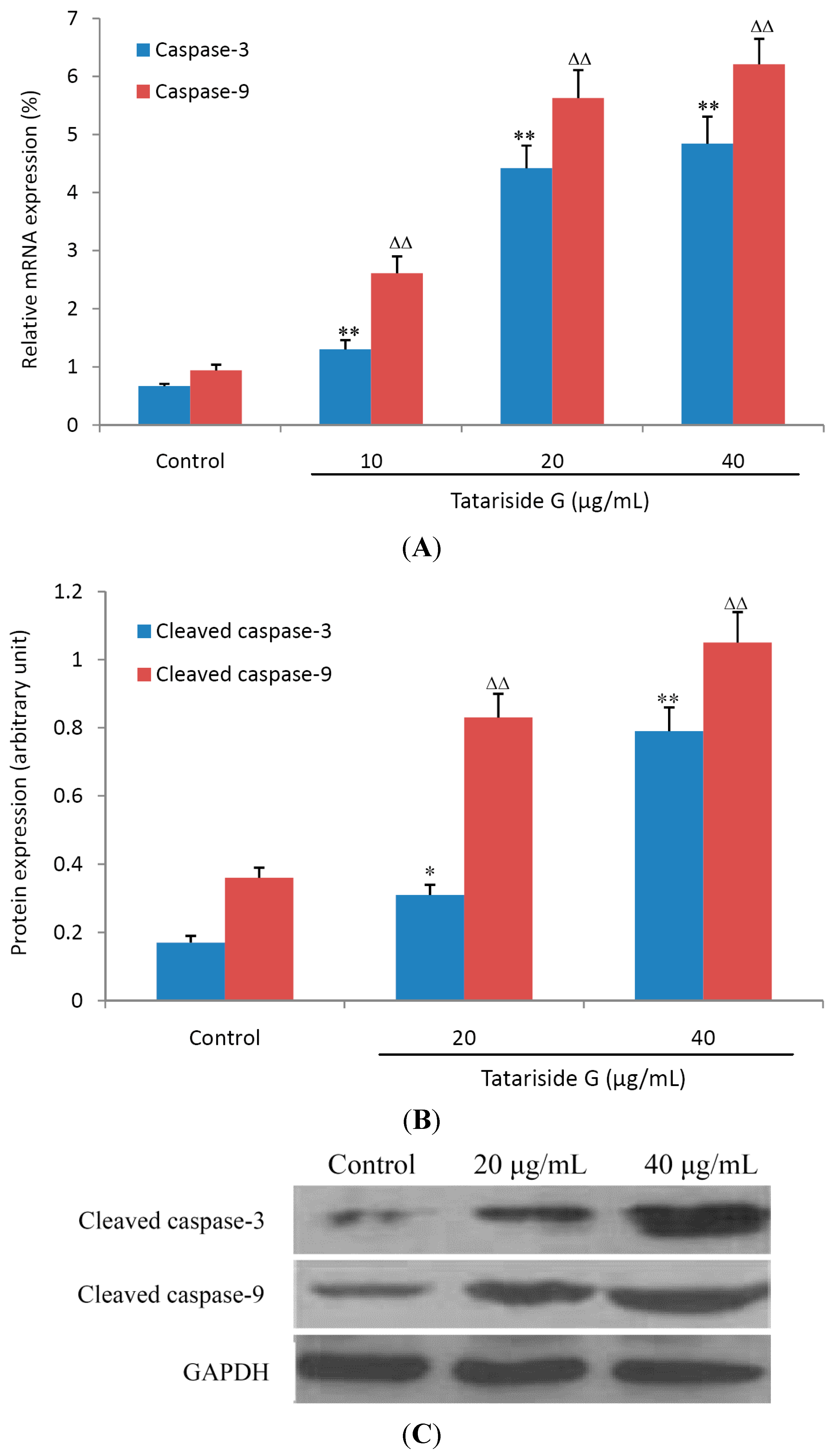
2.5. TG Elevates the mRNA and Cleaved Protein Expression of Caspase-3 and -9
2.6. Discussion

3. Experimental Section
3.1. Chemical and Reagents
3.2. Cell Culture
3.3. Determination of Cell Viability
3.4. Nuclear Morphology Assay
3.5. Annexin V-Propidium Iodide Apoptosis Detection
3.6. MMP Measurement
3.7. Western Blot Analysis
3.8. FQ-RT-PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer sequence | Species | Amplicon size (bp) |
|---|---|---|---|
| Caspase-3 | Forward: 5' AACTGGACTGTGGCATTGAG 3' | Human | 161 |
| Reverse: 5' ACAAAGCGACTGGATGAACC 3' | |||
| Caspase-9 | Forward: 5' CCTCACCCTGCCTTATCTTG 3' | Human | 189 |
| Reverse: 5' TCCCTCTTCCTCCACTGTTC 3' | |||
| GAPDH | Forward: 5' CACCCACTCCTCCACCTTTG 3' | Human | 110 |
| Reverse: 5' CCACCACCCTGTTGCTGTAG 3' |
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Su, J.H.; Wu, A.; Scotney, E.; Ma, B.; Monie, A.; Hung, C.F.; Wu, T.C. Immunotherapy for cervical cancer: Research status and clinical potential. Biodrugs 2010, 24, 109–129. [Google Scholar] [CrossRef]
- Ferguson, P.J.; Brisson, A.R.; Koropatnick, J.; Vincent, M.D. Enhancement of cytotoxicity of natural product drugs against multidrug resistant variant cell lines of human head and neck squamous cell carcinoma and breast carcinoma by tesmilifene. Cancer Lett. 2009, 274, 279–289. [Google Scholar] [CrossRef]
- Kim, H.G.; Song, H.; Yoon, D.H.; Song, B.W.; Park, S.M.; Sung, G.H.; Cho, J.Y.; Park, H.I.; Choi, S.; Song, W.O.; Hwang, K.C.; Kim, T.W. Cordyceps pruinosa extracts induce apoptosis of HeLa cells by a caspase dependent pathway. J. Ethnopharmacol. 2010, 128, 342–351. [Google Scholar] [CrossRef]
- Peng, B.; Hu, Q.; Liu, X.; Wang, L.; Chang, Q.; Li, J.; Tang, J.; Wang, N.; Wang, Y. Duchesnea phenolic fraction inhibits in vitro and in vivo growth of cervical cancer through induction of apoptosis and cell cycle arrest. Exp. Biol. Med. (Maywood) 2009, 234, 74–83. [Google Scholar] [CrossRef]
- Lin, B.; Hu, C.L.; Huang, F.; Han, T. Research progress on chemical constituents and pharmacological effect of Fagopyrum tataricum. Drugs Clinic 2011, 26, 29–32. [Google Scholar]
- Karki, R.; Park, C.H.; Kim, D.W. Extract of buckwheat sprouts scavenges oxidation and inhibits pro-inflammatory mediators in lipopolysaccharide-stimulated macrophages (RAW264.7). J. Integr. Med. 2013, 11, 246–252. [Google Scholar] [CrossRef]
- Guo, Z.J. Shanxi Qi Yao; Shanxi Science and Technology Press: Shanxi, China, 2003. [Google Scholar]
- Guo, Z.J.; Bu, X.X.; Wang, J.X.; Lv, J.X. Plant resources and research summary on “Shanxi Qi Yao”. Chin. J. Ethnomed. Ethnopharm. 2006, 7, 79–81. [Google Scholar]
- Brown, L.L.; Larson, S.R.; Sneden, A.T. Vanicosides C–F; new phenylpropanoid glycosides from Polygonum pensylvanicum. J. Nat. Prod. 1998, 61, 762–766. [Google Scholar] [CrossRef]
- Kuo, Y.H.; Hsu, Y.W.; Liaw, C.C.; Lee, J.K.; Huang, H.C.; Kuo, L.M. Cytotoxic phenylpropanoid glycosides from the stems of Smilax china. J. Nat. Prod. 2005, 68, 1475–1478. [Google Scholar] [CrossRef]
- Takasaki, M.; Konoshima, T.; Kuroki, S.; Tokuda, H.; Nishino, H. Cancer chemopreventive activity of phenylpropanoid esters of sucrose, vanicoside B and lapathoside A from Polygonum lapathifolium. Cancer Lett. 2001, 173, 133–138. [Google Scholar] [CrossRef]
- Saraste, A.; Pulkki, M.K. Morphologic and biochemical hallmarks of apoptosis. Cardiovasc. Res. 2000, 45, 528–537. [Google Scholar] [CrossRef]
- Wu, J.G.; Ma, L.; Zhang, S.Y.; Zhu., Z.Z.; Zhang, H.; Qin, L.P.; Wei, Y.J. Essential oil from rhizomes of Ligusticum chuanxiong induces apoptosis in hypertrophic scar fibroblasts. Pharm. Biol. 2011, 49, 86–93. [Google Scholar] [CrossRef]
- Thornberry, N.A. Caspases: Key mediators of apoptosis. Chem. Biol. 1998, 5, 97–103. [Google Scholar] [CrossRef]
- Mandal, S.K.; Biswas, R.; Bhattacharyya, S.S.; Paul, S.; Dutta, S.; Pathak, S.; Khuda-Bukhsh, A.R. Lycopodine from Lycopodium clavatum extract inhibits proliferation of HeLa cells through induction of apoptosis via caspase–3 activation. Eur. J. Pharmacol. 2010, 626, 115–122. [Google Scholar] [CrossRef]
- Wajant, H. The Fas signaling pathway: More than a paradigm. Science 2002, 296, 1635–1636. [Google Scholar] [CrossRef]
- Cargnello, M.; Roux, P.P. Activation and function of the MAPKs and their substrates; the MAPK–activated protein kinases. Microbiol. Mol. Biol. Rev. 2011, 75, 50–83. [Google Scholar] [CrossRef]
- Fresno, V.J.; Casado, E.; de Castro, J.; Cejas, P.; Belda-Iniesta, C.; Gonzalez-Baron, M. PI3K/Akt signalling pathway and cancer. Cancer Treat. Rev. 2004, 30, 193–204. [Google Scholar] [CrossRef]
- Sun, X.M.; MacFarlane, M.; Zhuang, J.; Wolf, B.B.; Green, D.R.; Cohen, G.M. Distinct caspase cascades are initiated in receptor-mediated and chemical-induced apoptosis. J. Biol. Chem. 1999, 274, 5053–5060. [Google Scholar] [CrossRef]
- Zheng, C.J.; Hu, C.L.; Ma, X.Q.; Peng, C.; Zhang, H.; Qin, L.P. Cytotoxic phenylpropanoid glycosides from Fagopyrum tataricum (L.) Gaertn. Food Chem. 2011, 132, 433–438. [Google Scholar]
- Zheng, G.Y.; Qin, L.P.; Yue, X.Q.; Gu, W.; Zhang, H.; Xing, H.L. Portulacerebroside A induces apoptosis via activation of the mitochondrial death pathway in human liver cancer HCCLM3 cells. Phytochem. Lett. 2014, 7, 77–84. [Google Scholar] [CrossRef]
- Ghosh, S.; Bishayee, K.; Paul, A.; Mukherjee, A.; Sikdar, S.; Chakraborty, D.; Boujedaini, N.; Khuda-Bukhsh, A.R. Homeopathic mother tincture of Phytolacca decandra induces apoptosis in skin melanoma cells by activating caspase-mediated signaling via reactive oxygen species elevation. J. Integr. Med. 2013, 11, 116–124. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, C.H.; Jiang, Y.P.; Peng, C.; He, K.; Tang, J.Y.; Xin, H.L. Protective effects of polydatin from Polygonum cuspidatum against carbon tetrachloride–induced liver injury in mice. PLoS One 2012, 7, e46574. [Google Scholar]
- Sample Availability: Please contact the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, Y.; Wang, S.-J.; Xia, W.; Rahman, K.; Zhang, Y.; Peng, H.; Zhang, H.; Qin, L.-P. Effects of Tatariside G Isolated from Fagopyrum tataricum Roots on Apoptosis in Human Cervical Cancer HeLa Cells. Molecules 2014, 19, 11145-11159. https://doi.org/10.3390/molecules190811145
Li Y, Wang S-J, Xia W, Rahman K, Zhang Y, Peng H, Zhang H, Qin L-P. Effects of Tatariside G Isolated from Fagopyrum tataricum Roots on Apoptosis in Human Cervical Cancer HeLa Cells. Molecules. 2014; 19(8):11145-11159. https://doi.org/10.3390/molecules190811145
Chicago/Turabian StyleLi, Yuan, Su-Juan Wang, Wei Xia, Khalid Rahman, Yan Zhang, Hao Peng, Hong Zhang, and Lu-Ping Qin. 2014. "Effects of Tatariside G Isolated from Fagopyrum tataricum Roots on Apoptosis in Human Cervical Cancer HeLa Cells" Molecules 19, no. 8: 11145-11159. https://doi.org/10.3390/molecules190811145