Eurycomanone and Eurycomanol from Eurycoma longifolia Jack as Regulators of Signaling Pathways Involved in Proliferation, Cell Death and Inflammation



Abstract
:1. Introduction

2. Results and Discussion
2.1. Results
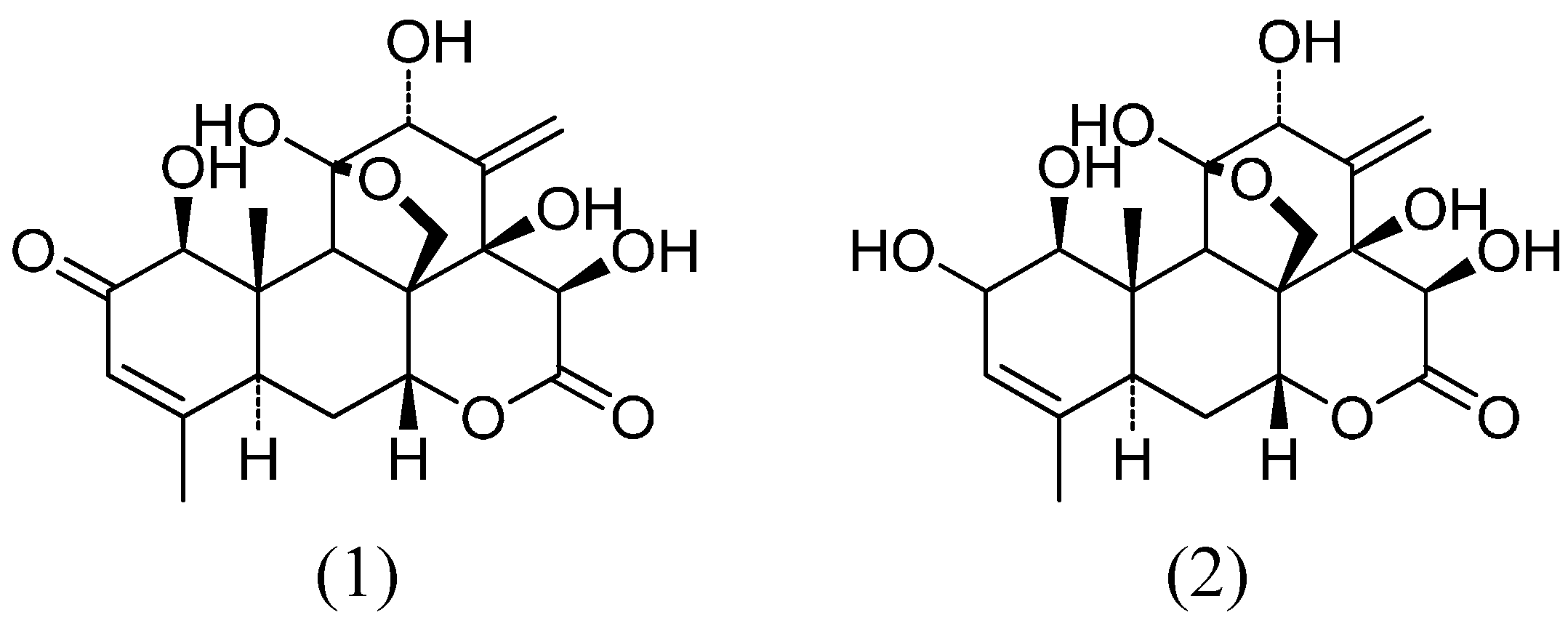
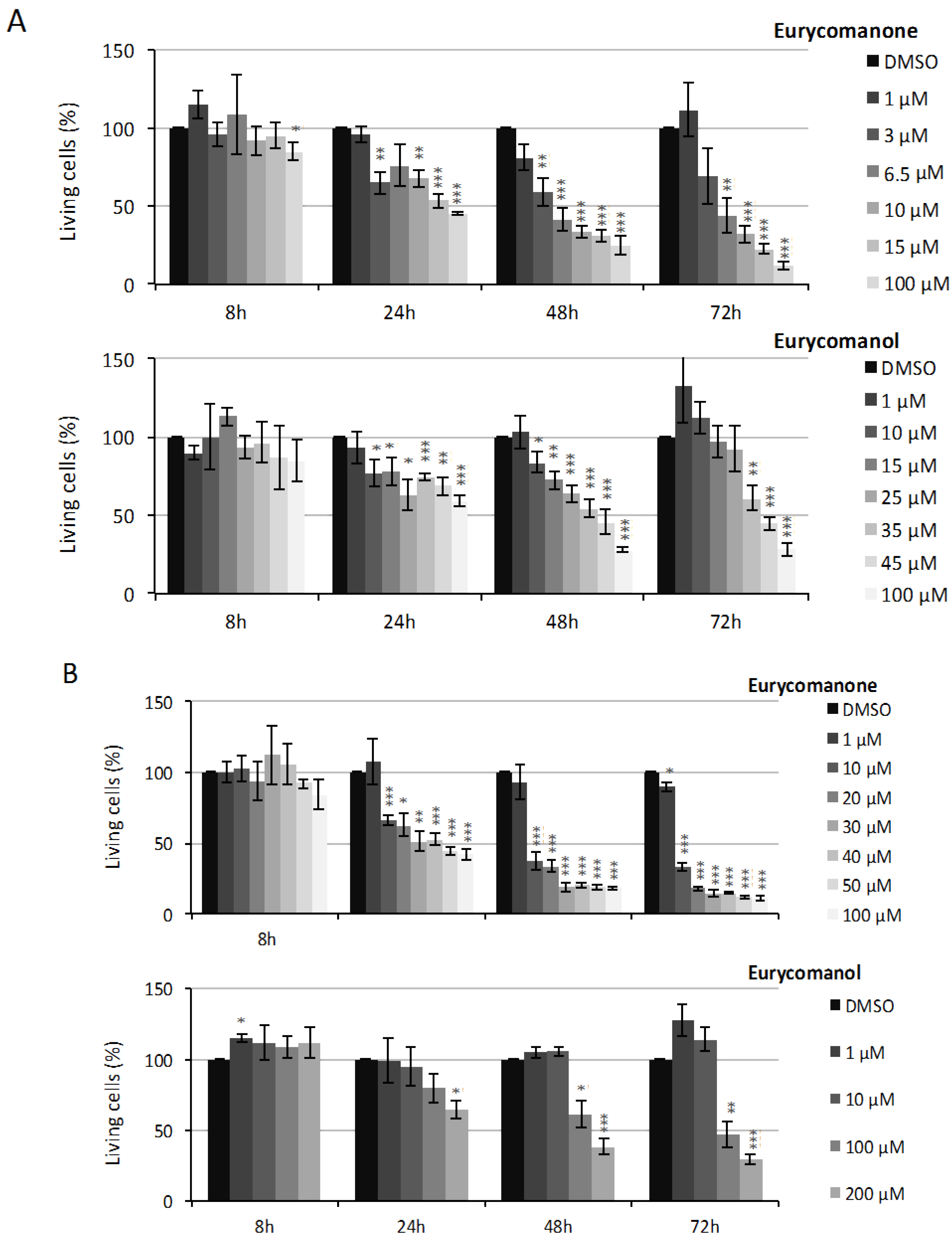
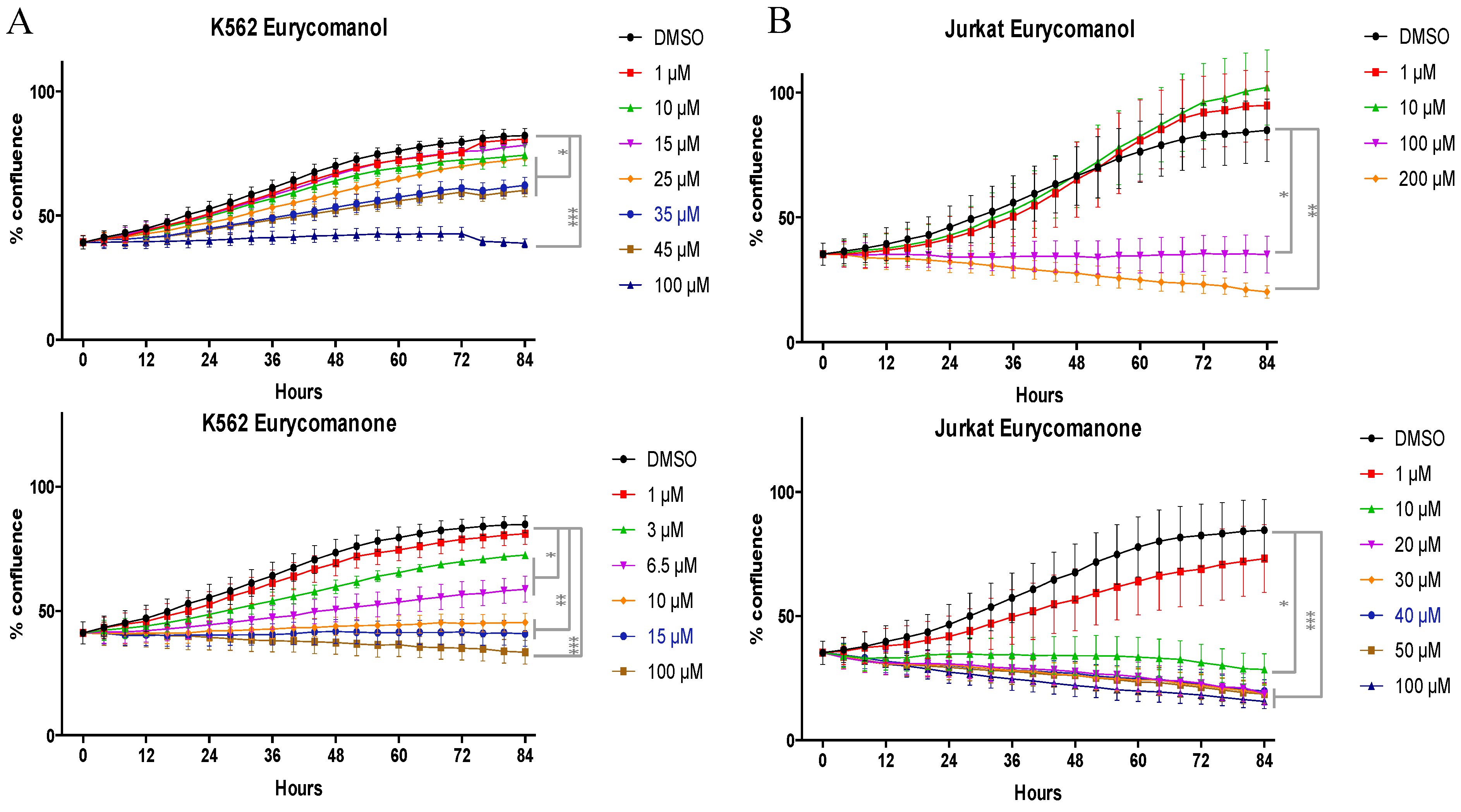
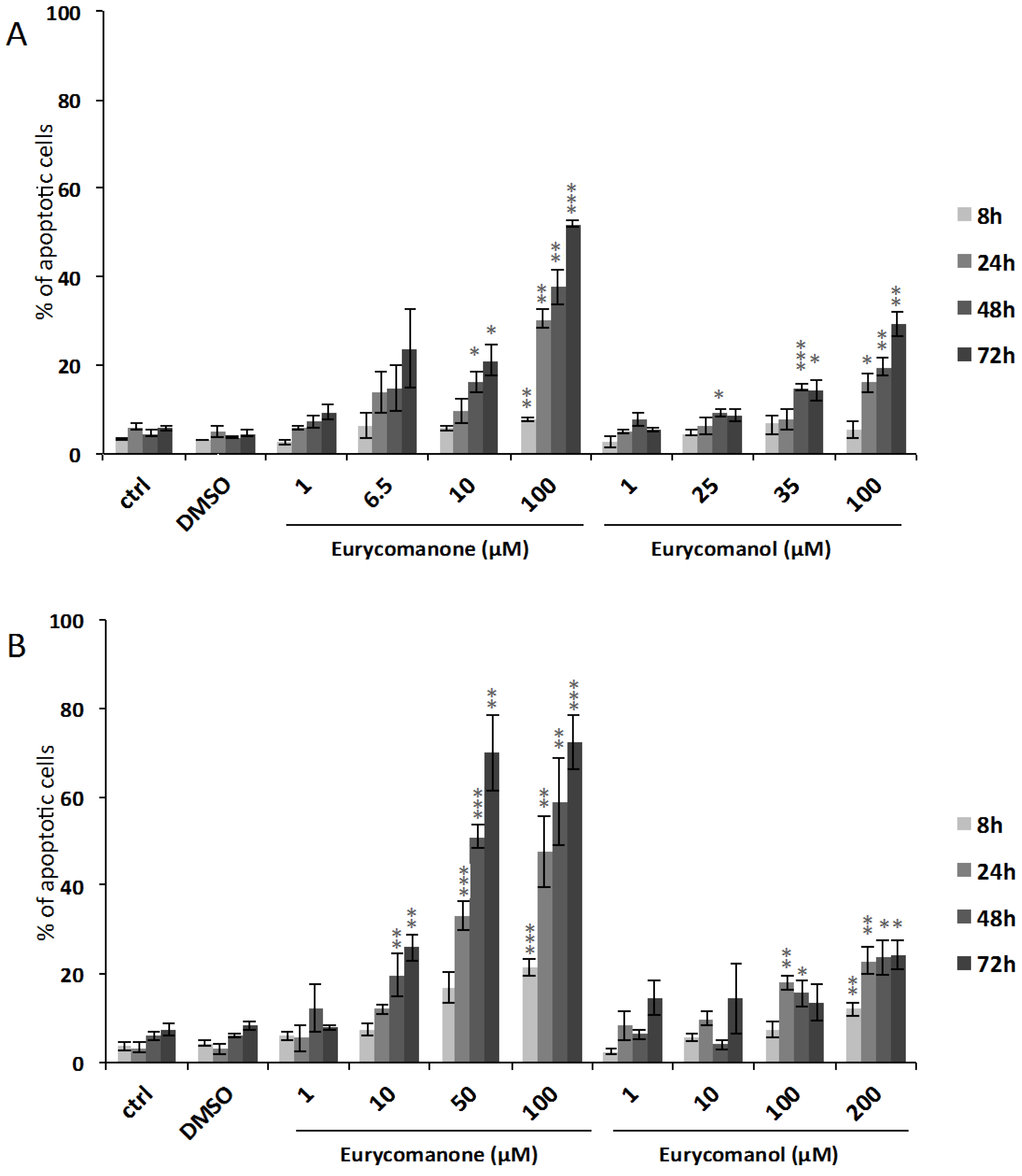
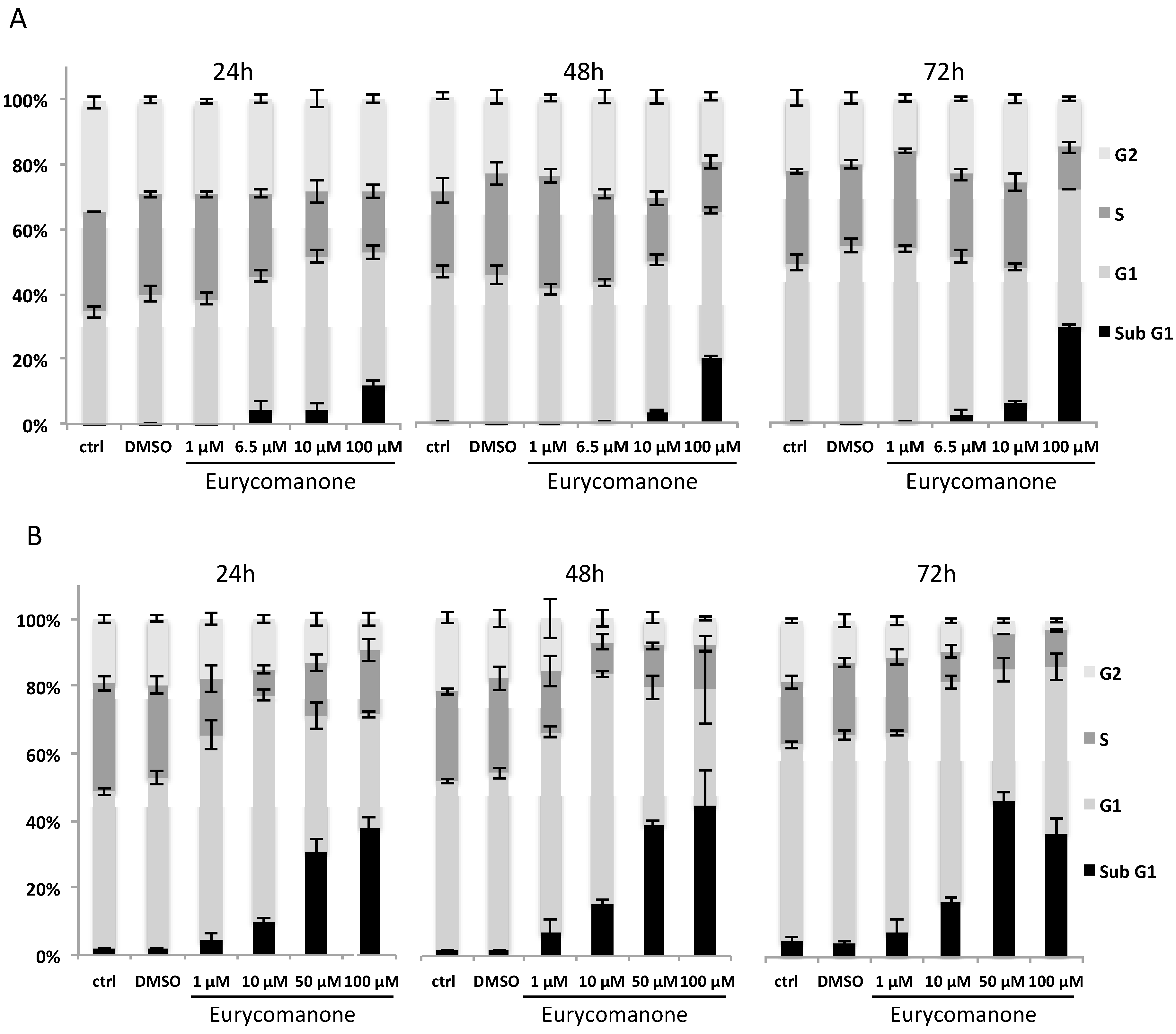
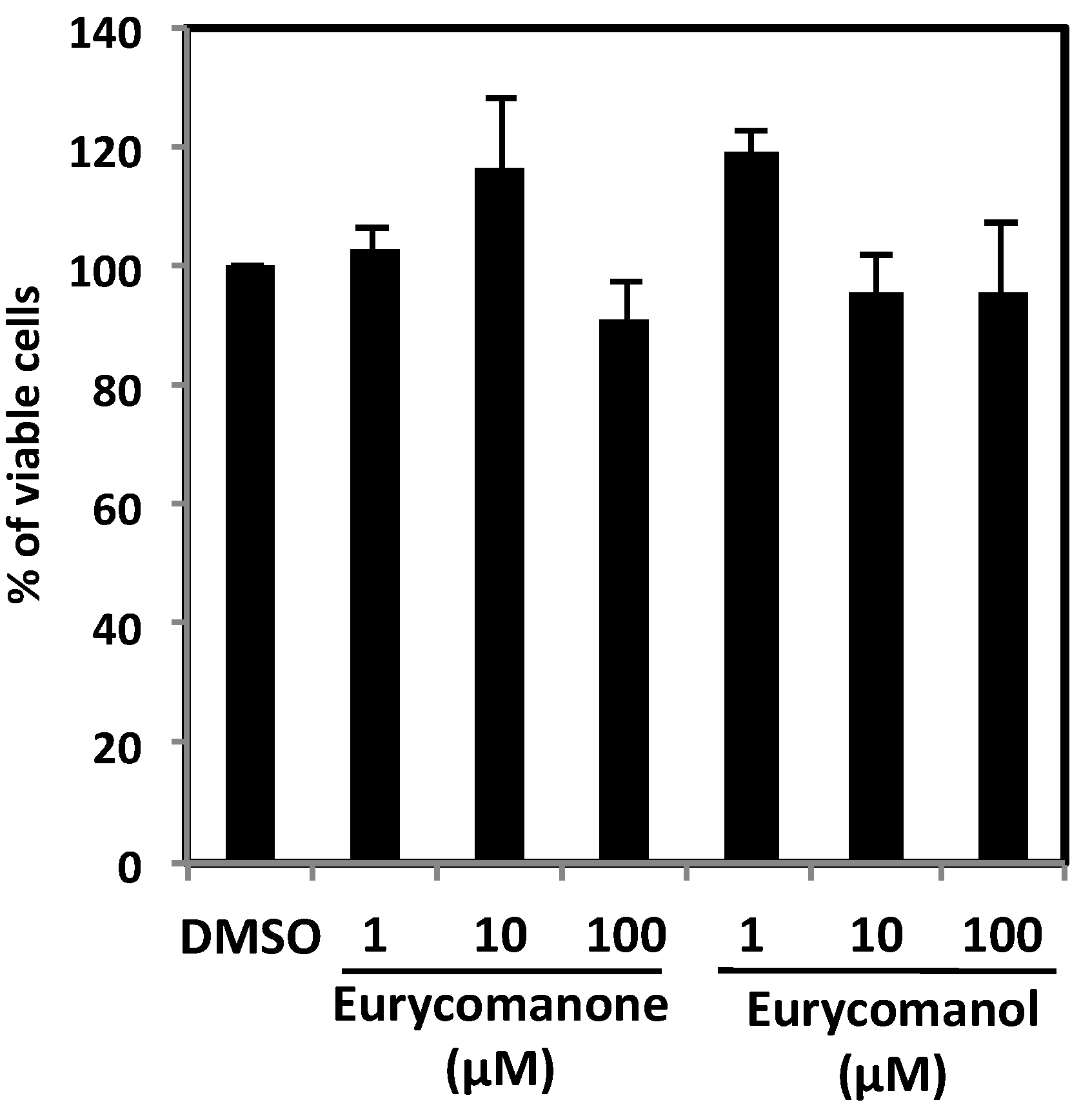
2.1.1. Eurycomanone and Eurycomanol Specifically Affect Cancer Cell Viability and Proliferation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cells | Compound | 8 h | 24 h | 48 h | 72 h |
|---|---|---|---|---|---|
| K562 | Eurycomanone | >150 µM | 48.92 µM | 4.25 µM | 5.7 µM |
| Eurycomanol | >150 µM | >150 µM | 39.78 µM | 46.4 µM | |
| Jurkat | Eurycomanone | >150 µM | 40.2 µM | 5.7 µM | 6.2 µM |
| Eurycomanol | >150 µM | >150 µM | 118.6 µM | 90.7 µM |


| Cells | Compound | 8 h | 24 h | 48 h | 72 h |
|---|---|---|---|---|---|
| K562 | Eurycomanone | >150 µM | >150 µM | >150 µM | 14.2 µM |
| Eurycomanol | >200 µM | >200 µM | >200 µM | 143.8 µM | |
| Jurkat | Eurycomanone | >150 µM | >150 µM | 9.58 µM | 5.89 µM |
| Eurycomanol | >150 µM | >150 µM | 102.1 µM | 93.9 µM |

| Compound | Inhibition of Cell Viability at 24 h (µM) | Inhibition of Cell Proliferation at 72 h (µM) | Inhibition of NF-κB Activity at 8 h (µM) | Reference |
|---|---|---|---|---|
| Eurycomanol | >150 | 143.8 | 35.6 | present study |
| Eurycomanone | 48.92 | 14.2 | 6.6 | present study |
| 4-Hydroxychalcone | 31 | nd | 24 | [9] |
| 4-Methoxychalcone | 35 | nd | 29 | [9] |
| Altersolanol A | 4 | 1 | 0.8 | [20] |
| Butein | 13 | nd | 38 | [9] |
| Embellicine A | 2.9 | nd | 3 | [19] |
| Embellicine B | 0.3 | 0.5 | 0.4 | [19] |
| Flavokawain C | 13 | nd | 8 | [9] |
| Goniothalamin | >20 | nd | 7 | [12] |
| Homobutein | 29 | nd | 38 | [9] |
| Isoliquiritigenin | 44 | nd | 32 | [9] |
| Phloretin | 59 | nd | 41 | [9] |
| Tetraacetylaltersolanol | 2.5 | nd | 4.6 | [20] |


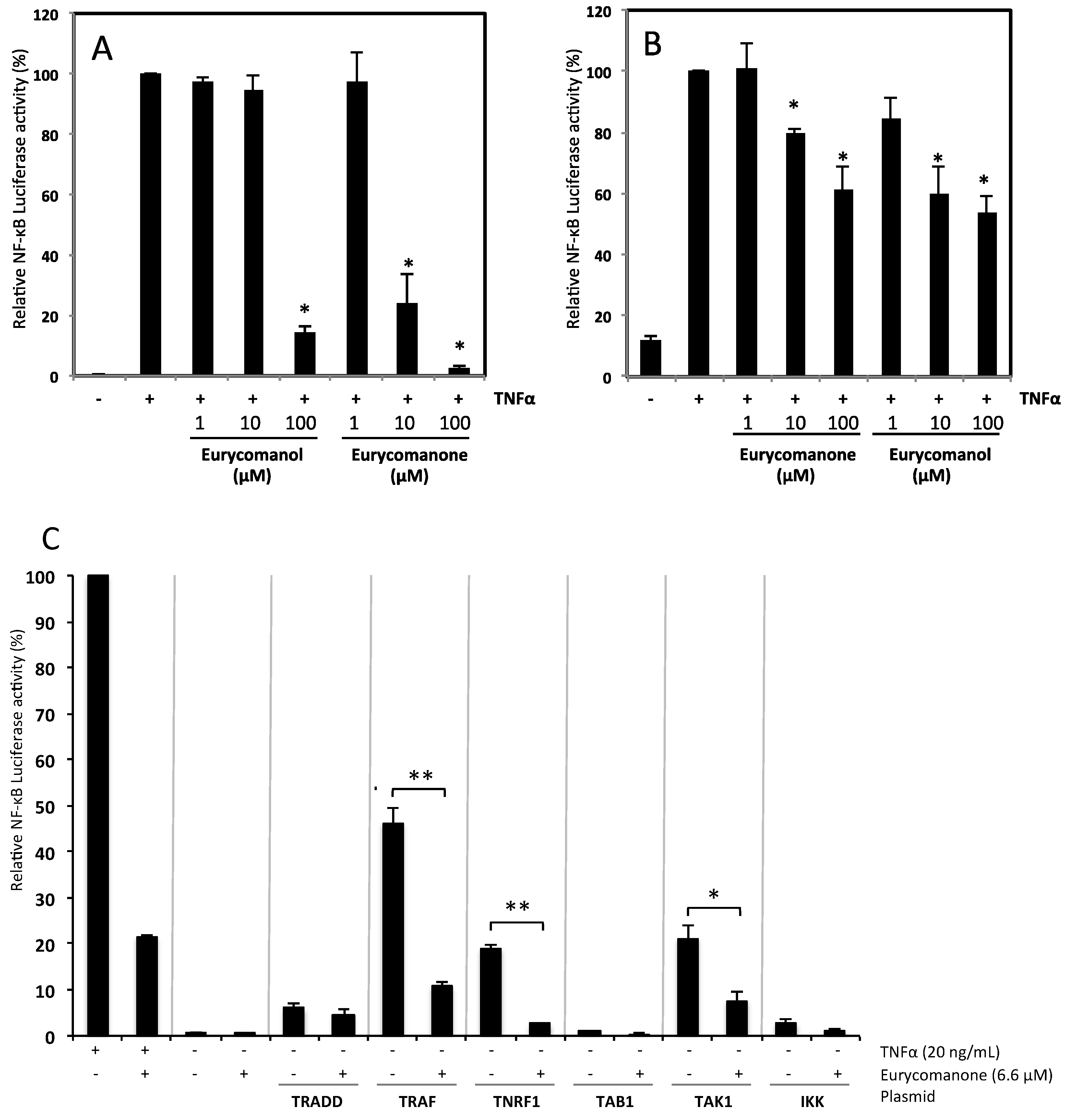
2.1.2. Eurycomanone and Eurycomanol Inhibit TNFα-Induced NF-κB Pathway Activation

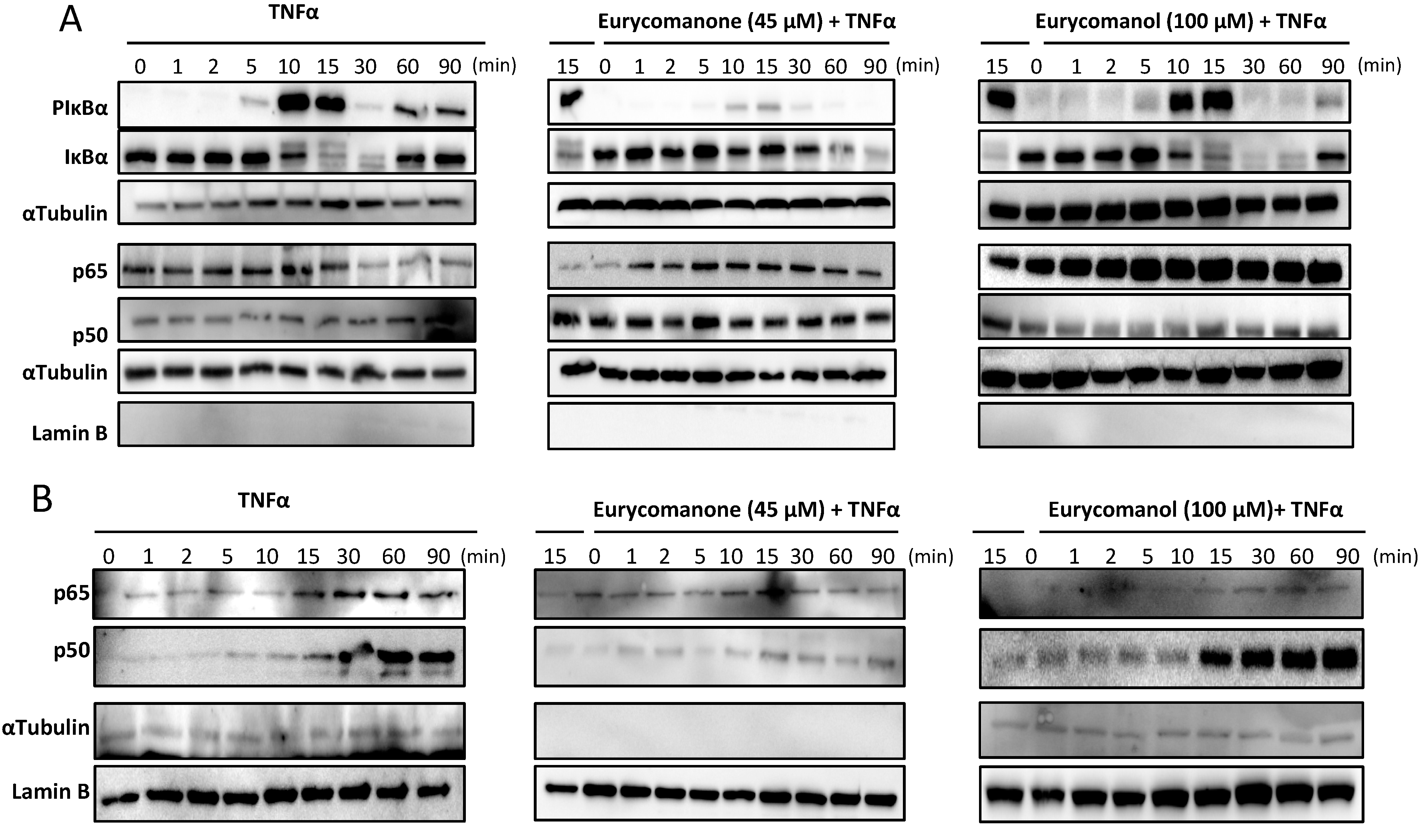
2.1.3. Eurycomanone Inhibits TNFα-Dependent Degradation of IκBα and Prevents p50/p65 Nuclear Translocation

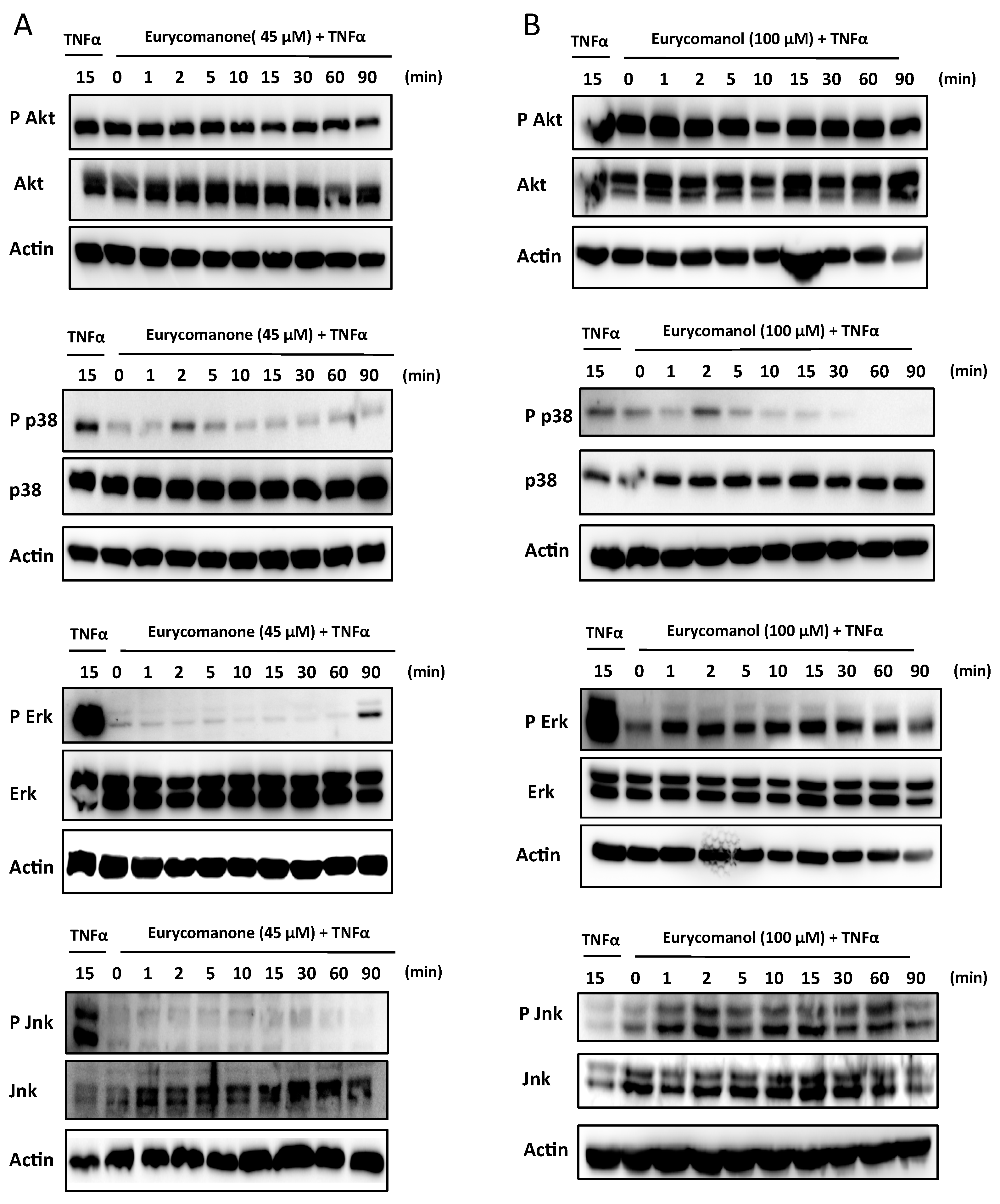
2.1.4. Eurycomanone Inhibits MAPK Activation

2.2. Discussion
3. Experimental Section
3.1. Plant Material
3.2. Isolation of Pure Compounds
3.3. Cell Culture
3.4. Cell Viability Assessment
3.5. Proliferation Assay
3.6. Analysis of Nuclear Fragmentation
3.7. Cell Cycle Analysis
3.8. Transient Transfection and Luciferase Reporter Gene Assay
3.9. Extraction of Cellular Proteins
3.10. Western Blot Analysis
3.11. Statistical Analysis
4. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Newman, D.; Cragg, G. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef]
- Bhat, R.; Karim, A.A. Tongkat ali (Eurycoma longifolia jack): A review on its ethnobotany and pharmacological importance. Fitoterapia 2010, 81, 669–679. [Google Scholar] [CrossRef]
- Delhalle, S.; Blasius, R.; Dicato, M.; Diederich, M. A beginner’s guide to NF-kappab signaling pathways. Ann. N. Y. Acad. Sci. 2004, 1030, 1–13. [Google Scholar]
- Folmer, F.; Blasius, R.; Morceau, F.; Tabudravu, J.; Dicato, M.; Jaspars, M.; Diederich, M. Inhibition of TNFalpha-induced activation of nuclear factor kappab by kava (piper methysticum) derivatives. Biochem. Pharmacol. 2006, 71, 1206–1218. [Google Scholar] [CrossRef]
- Folmer, F.; Dicato, M.; Diederich, M. From the deepest sea shelf to the uppermost kitchen cabinet shelf: The quest for novel TNF-alpha inhibitors. Curr. Top. Med. Chem. 2012, 12, 1392–1407. [Google Scholar] [CrossRef]
- Folmer, F.; Jaspars, M.; Solano, G.; Cristofanon, S.; Henry, E.; Tabudravu, J.; Black, K.; Green, D.H.; Kupper, F.C.; Aalbersberg, W.; et al. The inhibition of TNF-alpha-induced NF-kappab activation by marine natural products. Biochem. Pharmacol. 2009, 78, 592–606. [Google Scholar] [CrossRef]
- Orlikova, B.; Legrand, N.; Panning, J.; Dicato, M.; Diederich, M. Anti-inflammatory and anticancer drugs from nature. Cancer Treat. Res. 2014, 159, 123–143. [Google Scholar] [CrossRef]
- Orlikova, B.; Tasdemir, D.; Golais, F.; Dicato, M.; Diederich, M. The aromatic ketone 4'-hydroxychalcone inhibits TNFalpha-induced NF-kappab activation via proteasome inhibition. Biochem. Pharmacol. 2011, 82, 620–631. [Google Scholar] [CrossRef]
- Orlikova, B.; Schnekenburger, M.; Zloh, M.; Golais, F.; Diederich, M.; Tasdemir, D. Natural chalcones as dual inhibitors of hdacs and NF-kappab. Oncol. Rep. 2012, 28, 797–805. [Google Scholar]
- Duvoix, A.; Blasius, R.; Delhalle, S.; Schnekenburger, M.; Morceau, F.; Henry, E.; Dicato, M.; Diederich, M. Chemopreventive and therapeutic effects of curcumin. Cancer Lett. 2005, 223, 181–190. [Google Scholar] [CrossRef]
- Chen, W.Y.; Wu, C.C.; Lan, Y.H.; Chang, F.R.; Teng, C.M.; Wu, Y.C. Goniothalamin induces cell cycle-specific apoptosis by modulating the redox status in mda-mb-231 cells. Eur. J. Pharmacol. 2005, 522, 20–29. [Google Scholar] [CrossRef]
- Orlikova, B.; Schumacher, M.; Juncker, T.; Yan, C.C.; Inayat-Hussain, S.H.; Hajjouli, S.; Cerella, C.; Dicato, M.; Diederich, M. Styryl-lactone goniothalamin inhibits TNF-alpha-induced NF-kappab activation. Food Chem. Toxicol. 2013, 59, 572–578. [Google Scholar] [CrossRef]
- Tran, T.V.; Malainer, C.; Schwaiger, S.; Atanasov, A.G.; Heiss, E.H.; Dirsch, V.M.; Stuppner, H. NF-kappaB inhibitors from eurycoma longifolia. J. Nat. Prod. 2014, 77, 483–488. [Google Scholar] [CrossRef]
- Juncker, T.; Cerella, C.; Teiten, M.H.; Morceau, F.; Schumacher, M.; Ghelfi, J.; Gaascht, F.; Schnekenburger, M.; Henry, E.; Dicato, M.; et al. Unbs1450, a steroid cardiac glycoside inducing apoptotic cell death in human leukemia cells. Biochem. Pharmacol. 2011, 81, 13–23. [Google Scholar] [CrossRef]
- Schumacher, M.; Cerella, C.; Eifes, S.; Chateauvieux, S.; Morceau, F.; Jaspars, M.; Dicato, M.; Diederich, M. Heteronemin, a spongean sesterterpene, inhibits TNF alpha-induced NF-kappa b activation through proteasome inhibition and induces apoptotic cell death. Biochem. Pharmacol. 2010, 79, 610–622. [Google Scholar] [CrossRef]
- Folmer, F.; Jaspars, M.; Dicato, M.; Diederich, M. Marine natural products as targeted modulators of the transcription factor nf-kappab. Biochem. Pharmacol. 2008, 75, 603–617. [Google Scholar] [CrossRef]
- Mayer, A.M.; Glaser, K.B.; Cuevas, C.; Jacobs, R.S.; Kem, W.; Little, R.D.; McIntosh, J.M.; Newman, D.J.; Potts, B.C.; Shuster, D.E. The odyssey of marine pharmaceuticals: A current pipeline perspective. Trends Pharmacol. Sci. 2010, 31, 255–265. [Google Scholar] [CrossRef]
- Schumacher, M.; Kelkel, M.; Dicato, M.; Diederich, M. Gold from the sea: Marine compounds as inhibitors of the hallmarks of cancer. Biotechnol. Adv. 2011, 29, 531–547. [Google Scholar] [CrossRef]
- Ebrahim, W.; Aly, A.H.; Wray, V.; Mandi, A.; Teiten, M.H.; Gaascht, F.; Orlikova, B.; Kassack, M.U.; Lin, W.; Diederich, M.; et al. Embellicines a and b: Absolute configuration and NF-kappab transcriptional inhibitory activity. J. Med. Chem. 2013, 56, 2991–2999. [Google Scholar] [CrossRef]
- Teiten, M.H.; Mack, F.; Debbab, A.; Aly, A.H.; Dicato, M.; Proksch, P.; Diederich, M. Anticancer effect of altersolanol a, a metabolite produced by the endophytic fungus stemphylium globuliferum, mediated by its pro-apoptotic and anti-invasive potential via the inhibition of NF-kappab activity. Bioorg. Med. Chem. 2013, 21, 3850–3858. [Google Scholar] [CrossRef]
- Reuther, J.Y.; Reuther, G.W.; Cortez, D.; Pendergast, A.M.; Baldwin, A.S., Jr. A requirement for NF-kappab activation in BCR-ABL-mediated transformation. Genes Dev. 1998, 12, 968–981. [Google Scholar] [CrossRef]
- Kardono, L.B.; Angerhofer, C.K.; Tsauri, S.; Padmawinata, K.; Pezzuto, J.M.; Kinghorn, A.D. Cytotoxic and antimalarial constituents of the roots of eurycoma longifolia. J. Nat. Prod. 1991, 54, 1360–1367. [Google Scholar] [CrossRef]
- Wong, P.F.; Cheong, W.F.; Shu, M.H.; Teh, C.H.; Chan, K.L.; AbuBakar, S. Eurycomanone suppresses expression of lung cancer cell tumor markers, prohibitin, annexin 1 and endoplasmic reticulum protein 28. Phytomedicine 2012, 19, 138–144. [Google Scholar] [CrossRef]
- Zakaria, Y.; Rahmat, A.; Pihie, A.H.; Abdullah, N.R.; Houghton, P.J. Eurycomanone induce apoptosis in HepG2 cells via up-regulation of p53. Cancer Cell Int. 2009, 9, 16. [Google Scholar] [CrossRef]
- Miyake, K.; Tezuka, Y.; Awale, S.; Li, F.; Kadota, S. Canthin-6-one alkaloids and a tirucallanoid from eurycoma longifolia and their cytotoxic activity against a human ht-1080 fibrosarcoma cell line. Nat. Prod. Commun. 2010, 5, 17–22. [Google Scholar]
- Beutler, J.A.; Kang, M.I.; Robert, F.; Clement, J.A.; Pelletier, J.; Colburn, N.H.; McKee, T.C.; Goncharova, E.; McMahon, J.B.; Henrich, C.J. Quassinoid inhibition of ap-1 function does not correlate with cytotoxicity or protein synthesis inhibition. J. Nat. Prod. 2009, 72, 503–506. [Google Scholar] [CrossRef]
- Chan, K.L.; Choo, C.Y.; Morita, H.; Itokawa, H. High performance liquid chromatography in phytochemical analysis of eurycoma longifolia. Planta Med. 1998, 64, 741–745. [Google Scholar] [CrossRef]
- Choo, C.Y.; Chan, K.L. High performance liquid chromatography analysis of canthinone alkaloids from eurycoma longifolia. Planta Med. 2002, 68, 382–384. [Google Scholar] [CrossRef]
- Cerella, C.; Scherer, C.; Cristofanon, S.; Henry, E.; Anwar, A.; Busch, C.; Montenarh, M.; Dicato, M.; Jacob, C.; Diederich, M. Cell cycle arrest in early mitosis and induction of caspase-dependent apoptosis in u937 cells by diallyltetrasulfide (Al2S4). Apoptosis 2009, 14, 641–654. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hajjouli, S.; Chateauvieux, S.; Teiten, M.-H.; Orlikova, B.; Schumacher, M.; Dicato, M.; Choo, C.-Y.; Diederich, M. Eurycomanone and Eurycomanol from Eurycoma longifolia Jack as Regulators of Signaling Pathways Involved in Proliferation, Cell Death and Inflammation. Molecules 2014, 19, 14649-14666. https://doi.org/10.3390/molecules190914649
Hajjouli S, Chateauvieux S, Teiten M-H, Orlikova B, Schumacher M, Dicato M, Choo C-Y, Diederich M. Eurycomanone and Eurycomanol from Eurycoma longifolia Jack as Regulators of Signaling Pathways Involved in Proliferation, Cell Death and Inflammation. Molecules. 2014; 19(9):14649-14666. https://doi.org/10.3390/molecules190914649
Chicago/Turabian StyleHajjouli, Shéhérazade, Sébastien Chateauvieux, Marie-Hélène Teiten, Barbora Orlikova, Marc Schumacher, Mario Dicato, Chee-Yan Choo, and Marc Diederich. 2014. "Eurycomanone and Eurycomanol from Eurycoma longifolia Jack as Regulators of Signaling Pathways Involved in Proliferation, Cell Death and Inflammation" Molecules 19, no. 9: 14649-14666. https://doi.org/10.3390/molecules190914649