
Marine Sponge Lectins: Actual Status on Properties and Biological Activities

Abstract
:
1. Introduction
2. Characteristics of Lectins from Marine Sponges

{kind=link}
{kind=link}
{kind=link}
| Marine Sponges | Molecular Weight (kDa) | Conformation | Disulfide Bonds | Thermal Stability | Specificity | Carbohydrates (%) | Author |
|---|---|---|---|---|---|---|---|
| Aplysina archeri | 16 | Tetramer | − | N. D. | β-d-Gal-Oligossaccharides | 3.5 | [11] |
| Aplysina lawnosa | 16 | Tetramer | − | N.D. | β-d-Gal-Oligossaccharides | 5 | [11] |
| Axinella polypoides I | 21 | Dimer | − | d-Gal and d-Fuc | 0.5 | [17] | |
| Axinella polypoides II | 15 | Monomer | − | − | d-Gal and d-Fuc | 0.5 | [17] |
| Axinella corrugata I | 13.9 | Hexamer | + | + | N,N',N''-Triacetylchitotriose | N.D. | [18] |
| Axinella corrugata II | 13.9 | N.D | N.D. | + | N,N',N''-Triacetylchitotriose | N.D. | [18] |
| Cliona varians | 28.5 | Tetramer | + | + | Galactose | N.D. | [21] |
| Craniella australiensis | 17.8 | Trimer | + | + | Asialo-PSM | 27.6 | [22] |
| Cinachyrella alloclada | 17 | Monomer/Dimer | + | + | Lactose | + | [23] |
| Cinachyrella apion | 15.5 | Octamer | N.D. | + | Lactose | N.D. | [20] |
| Cinachyrella sp. | 16 | Tetramer | N.D. | + | Lactose | N.D. | [24] |
| Geodia cydonium | 13 | Trimer | + | N.D. | Lactose | 9.92 | [19] |
| Halichondria okadai | 30 | Dimer | − | − | Galactose | N.D. | [12] |
| Haliclona cratera | 29 | Monomer | − | Relatively | α-1 Glycoprotein and Mucin | 3.7 | [25] |
| Haliclona caerulea I | 14 | Monomer | N.D. | − | N.D. | N.D. | [26] |
| Haliclona caerulea II | 15 | Dimer | N.D. | + | N.D. | N.D. | [26] |
| Pellina semitubulosa | 34 | Hexamer | N.D. | Relatively | Lactose | 3.4 | [27] |

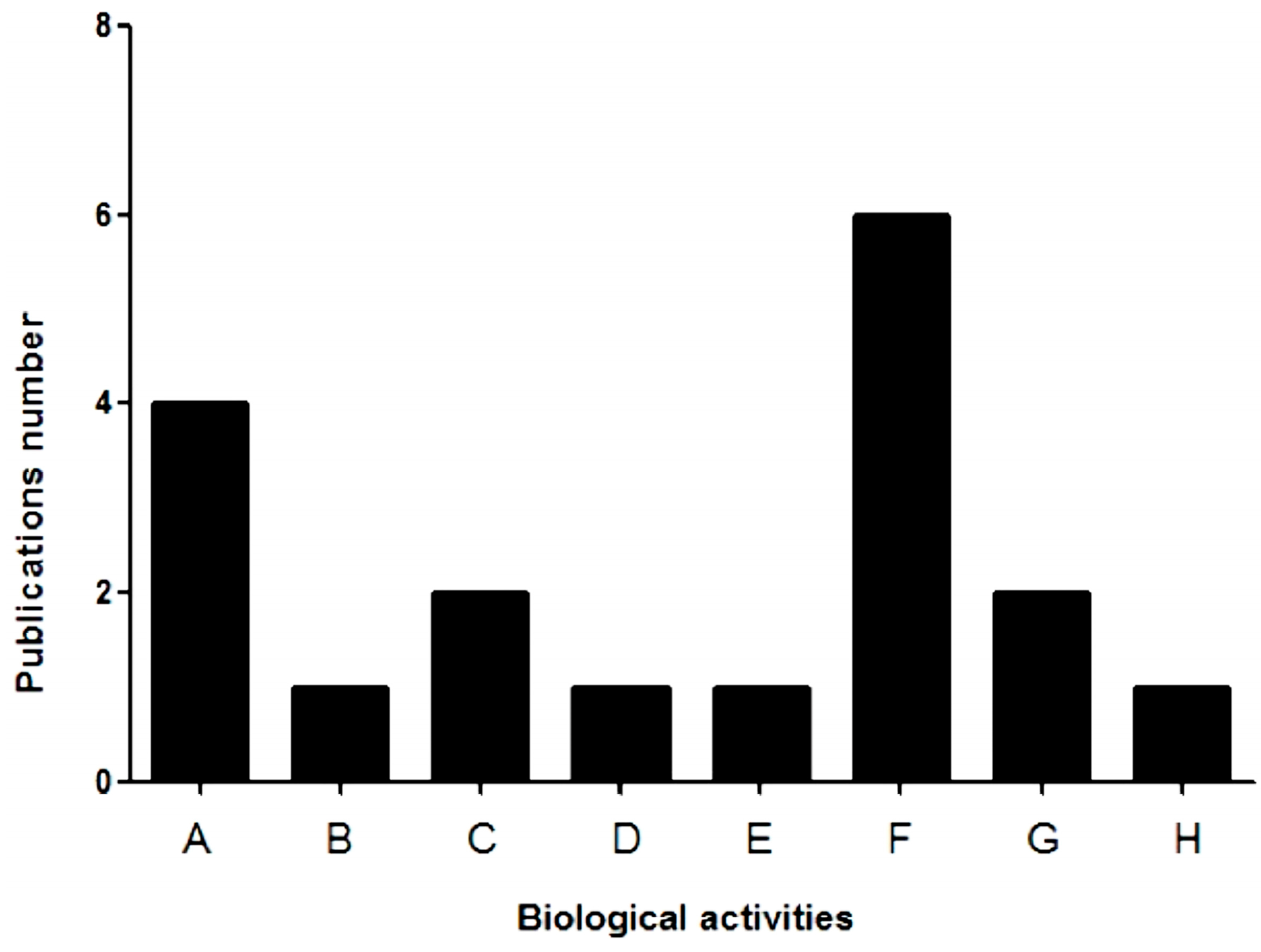
3. Biological Applications of Lectins from Marine Sponge

4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bakus, G.J.; Green, G. Toxicity in sponges and holothurians: A geographic pattern. Science 1974, 185, 951–953. [Google Scholar] [CrossRef] [PubMed]
- Sepcic, K.; Kauferstein, S.; Mebs, D.; Turk, T. Biological activities of aqueous and organic extracts from tropical marine sponges. Mar. Drugs 2010, 8, 1550–1566. [Google Scholar] [CrossRef] [PubMed]
- Berlinck, R.G.S.; Hajdu, E.; Rocha, R.M.; Oliveira, J.H.H.L.; Hernandez, I.L.C.; Seleghim, M.H.R.; Granato, A.C.; Almeida, E.V.R.; Nunez, C.V.; Muricy, G.; et al. Challenges and rewards of research in marine natural products chemistry in Brazil. J. Nat. Prod. 2004, 67, 510–522. [Google Scholar] [CrossRef] [PubMed]
- Carte, B.K. Biomedical potential of marine natural products. Bioscience 1996, 46, 271–286. [Google Scholar] [CrossRef]
- Cragg, G.M.; Newman, D.J. Discovery and development of antineoplastic agents from natural sources. Cancer Investig. 1999, 17, 153–163. [Google Scholar] [CrossRef]
- Peumans, W.J.; van Damme, E.J. Lectins as plant defense proteins. Plant Physiol. 1995, 109, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Lakhtin, V.; Lakhtin, M.; Alyoshkin, V. Lectins of living organisms. The overview. Anaerobe 2011, 17, 452–455. [Google Scholar] [CrossRef] [PubMed]
- Sumner, J.B.; Howell, S.F. Identification of Hemagglutinin of Jack Bean with Concanavalin A. J. Bacteriol. 1936, 32, 227–237. [Google Scholar] [PubMed]
- Dodd, R.Y.; MacLennan, A.P.; Hawkins, D.C. Haemagglutinins from marine sponges. Vox Sanguinis 1968, 15, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Mebs, D.; Weiler, I.; Heinke, H.F. Bioactive proteins from marine sponges: Screening of sponge extracts for hemagglutinating, hemolytic, ichthyotoxic and lethal properties and isolation and characterization of hemagglutinins. Toxicon 1985, 23, 955–962. [Google Scholar] [CrossRef] [PubMed]
- Miarons, P.B.; Fresno, M. Lectins from tropical sponges. Purification and characterization of lectins from genus Aplysina. J. Biol. Chem. 2000, 275, 29283–29289. [Google Scholar] [CrossRef] [PubMed]
- Kawsar, S.M.A.; Mamun, S.M.A.; Rahman, M.D.S.; Hidetaro, Y.; Ozeki, Y. In vitro antibacterial and antifungal effects of a 30 kDa d-galactoside-specific lectin from the desmosponge Helichondria okadai. Int. J. Biol. Life Sci. 2011, 7, 31–37. [Google Scholar]
- Queiroz, A.; Silva, R.; Moura, R.; Dreyfuss, J.; Paredes-Gamero, E.; Souza, A.; Tersariol, I.; Santos, E.; Nader, H.; Justo, G.; et al. Growth inhibitory activity of a novel lectin from Cliona varians against K562 human erythroleukemia cells. Cancer Chemother. Pharmacol. 2009, 63, 1023–1033. [Google Scholar] [CrossRef] [PubMed]
- Rüdiger, H.; Gabius, H.J. Plant lectins: Occurrence, biochemistry, functions and applications. Glycoconjugate J. 2001, 18, 589–613. [Google Scholar] [CrossRef]
- Gabius, H.J. Animal lectins. Eur. J. Biochem. 1997, 243, 543–576. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Bhaglal, P.; Bhullar, S. Wheat germ agglutinin (WGA) gene expression and ABA accumulation in the developing embryos of wheat (Triticum aestivum) in response to drought. Plant Growth Regul. 2000, 30, 145–150. [Google Scholar] [CrossRef]
- Bretting, H.; Kabat, E.A. Purification and characterization of the agglutinins from the sponge Axinella polypoides and a study of their combining sites. Biochemistry 1976, 15, 3228–3236. [Google Scholar] [CrossRef] [PubMed]
- Dresch, R.R.; Zanetti, G.D.; Lerner, C.B.; Mothes, B.; Trindade, V.M.T.; Henriques, A.T.; Vozári-Hampe, M.M. ACL-I, a lectin from the marine sponge Axinella corrugata: Isolation, characterization and chemotactic activity. Comp. Biochem. Physiol. Toxicol. Pharmacol. 2008, 148, 23–30. [Google Scholar] [CrossRef]
- Müller, W.E.G.; Conrad, J.; Schröder, C.; Zahn, R.K.; Kurelec, B.; Dreesbach, K.; Uhlenbruck, G. Characterization of the Trimeric, Self-Recognizing Geodia cydonium Lectin I. Eur. J. Biochem. 1983, 133, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, D.S.; Medeiros, T.L.; Ribeiro, J.K.C.; Monteiro, N.K.V.; Migliolo, L.; Uchoa, A.F.; Vasconcelos, I.M.; Oliveira, A.S.; de Sales, M.P.; Santos, E.A. A lactose specific lectin from the sponge Cinachyrella apion: Purification, characterization, N-terminal sequences alignment and agglutinating activity on Leishmania promastigotes. Comp. Biochem. Physiol. Biochem. Mol. Biol. 2010, 155, 211–216. [Google Scholar] [CrossRef]
- Moura, R.M.; Queiroz, A.F.S.; Fook, J.M.S.L.L.; Dias, A.S.F.; Monteiro, N.K.V.; Ribeiro, J.K.C.; Moura, G.E.D.D.; Macedo, L.L.P.; Santos, E.A.; Sales, M.P. CvL, a lectin from the marine sponge Cliona varians: Isolation, characterization and its effects on pathogenic bacteria and Leishmania promastigotes. Comp. Biochem. Physiol. Mol. Integr. Physiol. 2006, 145, 517–523. [Google Scholar] [CrossRef]
- Xiong, C.; Li, W.; Liu, H.; Zhang, W.; Dou, J.; Bai, X.; Du, Y.; Ma, X. A normal mucin-binding lectin from the sponge Craniella australiensis. Comp. Biochem. Physiol. Toxicol. Pharmacol. 2006, 143, 9–16. [Google Scholar] [CrossRef]
- Atta, A.M.; Barral-Netto, M.; Peixinho, S.; Sousa-Atta, M.L. Isolation and functional characterization of a mitogenic lectin from the marine sponge Cinachyrella alloclada. Braz. J. Med. Biol. Res. 1989, 22, 379–385. [Google Scholar] [PubMed]
- Ueda, T.; Nakamura, Y.; Smith, C.M.; Copits, B.A.; Inoue, A.; Ojima, T.; Matsunaga, S.; Swanson, G.T.; Sakai, R. Isolation of novel prototype galectins from the marine ball sponge Cinachyrella sp. guided by their modulatory activity on mammalian glutamate-gated ion channels. Glycobiology 2013, 23, 412–425. [Google Scholar] [CrossRef] [PubMed]
- Pajic, I.; Kljajic, Z.; Dogovic, N.; Sladic, D.; Juranic, Z.; Gasic, M.J. A novel lectin from the sponge Haliclona cratera: Isolation, characterization and biological activity. Comp. Biochem. Physiol. Toxicol. Pharmacol. 2002, 132, 213–221. [Google Scholar] [CrossRef]
- Carneiro, R.F.; de Melo, A.A.; Nascimento, F.E.P.D.; Simplicio, C.A.; Nascimento, K.S.d.; Rocha, B.A.M.D.; Saker-Sampaio, S.; Moura, R.D.M.; Mota, S.S.; Cavada, B.S.; et al. Halilectin 1 (H-1) and Halilectin 2 (H-2): Two new lectins isolated from the marine sponge Haliclona caerulea. J. Mol. Recognit. 2013, 26, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Engel, M.; Bachmann, M.; Schröder, H.C.; Rinkevich, B.; Kljajic, Z.; Uhlenbruck, G.; Müller, W.E.G. A novel galactose- and arabinose-specific lectin from the sponge Pellina semitubulosa: Isolation, characterization and immunobiological properties. Biochimie 1992, 74, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Freymann, D.M.; Nakamura, Y.; Focia, P.J.; Sakai, R.; Swanson, G.T. Structure of a tetrameric galectin from Cinachyrella sp. (ball sponge). Acta Crystallogr. Biol. Crystallogr. 2012, 68, 1163–1174. [Google Scholar] [CrossRef]
- Sharon, N.; Lis, H. The structural basis for carbohydrate recognition by lectins. Adv. Exp. Med. Biol. 2001, 491, 1–16. [Google Scholar] [PubMed]
- Dresch, R.R.; Lerner, C.B.; Mothes, B.; Trindade, V.M.T.; Henriques, A.T.; Vozári-Hampe, M.M. Biological activities of ACL-I and physicochemical properties of ACL-II, lectins isolated from the marine sponge Axinella corrugata. Comp. Biochem. Physiol. Biochem. Mol. Biol. 2012, 161, 365–370. [Google Scholar] [CrossRef]
- Dresch, R.R.; Zanetti, G.D.; Irazoqui, F.J.; Sendra, V.G.; Zlocowski, N.; Bernardi, A.; Rosa, R.M.; Battastini, A.M.O.; Henriques, A.T.; Vozári-Hampe, M.M. Staining tumor cells with biotinylated ACL-I, a lectin isolated from the marine sponge, Axinella corrugata. Biotech. Histochem. 2013, 88, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rabelo, L.; Monteiro, N.; Serquiz, R.; Santos, P.; Oliveira, R.; Oliveira, A.; Rocha, H.; Morais, A.H.; Uchoa, A.; Santos, E. A lactose-binding lectin from the marine sponge Cinachyrella Apion (cal) induces cell death in human cervical adenocarcinoma cells. Mar. Drugs 2012, 10, 727–743. [Google Scholar] [CrossRef] [PubMed]
- Kazanjian, A.; Fariñas, M. Actividades biológicas del extracto acuoso de la esponja Aplysina lacunosa (Porifera: Aplysinidae). Rev. Biol. Trop. 2006, 54, 189–200. [Google Scholar] [PubMed]
- Leite, J.F.; Assreuy, A.M.; Mota, M.R.; Bringel, P.H.; Lacerda, R.R.; Gomes Vde, M.; Cajazeiras, J.B.; Nascimento, K.S.; Pessoa Hde, L.; Gadelha, C.A.; et al. Antinociceptive and anti-inflammatory effects of a lectin-like substance from Clitoria fairchildiana R. Howard seeds. Molecules 2012, 17, 3277–3290. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, A.F.S.; Moura, R.M.; Ribeiro, J.K.C.; Lyra, I.L.; Cunha, D.C.S.; Santos, E.A.; de-Sales, M.P. Pro-inflammatory effect in mice of CvL, a lectin from the marine sponge Cliona varians. Comp. Biochem. Physiol. Toxicol. Pharmacol. 2008, 147, 216–221. [Google Scholar] [CrossRef]
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes Filho, S.M.; Cardoso, J.D.; Anaya, K.; Silva do Nascimento, E.; De Lacerda, J.T.J.G.; Mioso, R.; Santi Gadelha, T.; De Almeida Gadelha, C.A. Marine Sponge Lectins: Actual Status on Properties and Biological Activities. Molecules 2015, 20, 348-357. https://doi.org/10.3390/molecules20010348
Gomes Filho SM, Cardoso JD, Anaya K, Silva do Nascimento E, De Lacerda JTJG, Mioso R, Santi Gadelha T, De Almeida Gadelha CA. Marine Sponge Lectins: Actual Status on Properties and Biological Activities. Molecules. 2015; 20(1):348-357. https://doi.org/10.3390/molecules20010348
Chicago/Turabian StyleGomes Filho, Sandro Mascena, Juscélio Donizete Cardoso, Katya Anaya, Edilza Silva do Nascimento, José Thalles Jucelino Gomes De Lacerda, Roberto Mioso, Tatiane Santi Gadelha, and Carlos Alberto De Almeida Gadelha. 2015. "Marine Sponge Lectins: Actual Status on Properties and Biological Activities" Molecules 20, no. 1: 348-357. https://doi.org/10.3390/molecules20010348
APA StyleGomes Filho, S. M., Cardoso, J. D., Anaya, K., Silva do Nascimento, E., De Lacerda, J. T. J. G., Mioso, R., Santi Gadelha, T., & De Almeida Gadelha, C. A. (2015). Marine Sponge Lectins: Actual Status on Properties and Biological Activities. Molecules, 20(1), 348-357. https://doi.org/10.3390/molecules20010348