
Identification and Expression Analysis of Zebrafish (Danio rerio) E-Selectin during Embryonic Development



Abstract
:1. Introduction
2. Results and Discussion
2.1. Cloning of Zebrafish E-Selectin
2.2. Nucleotide Sequence Analysis of Zebrafish E-Selectin
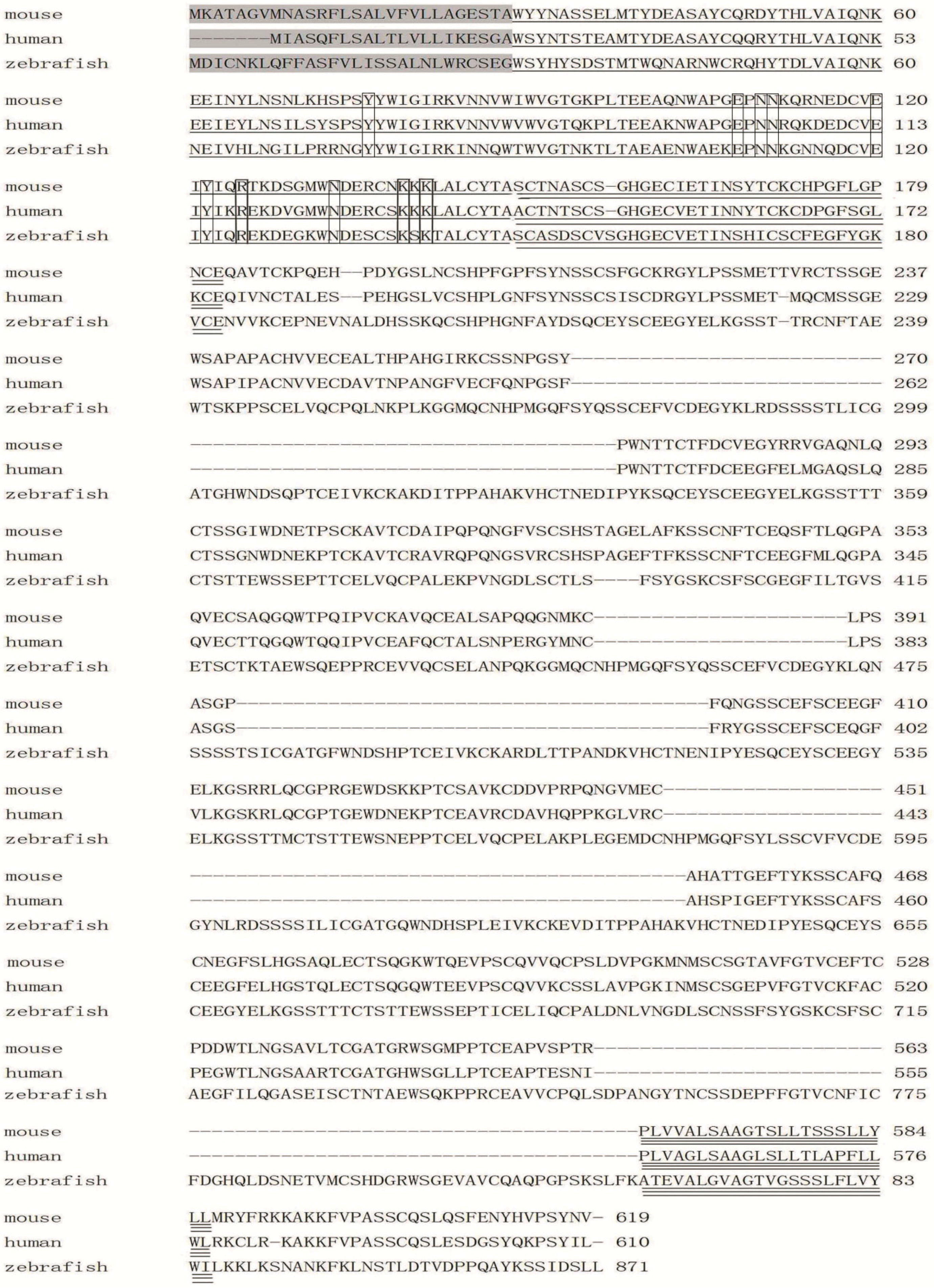
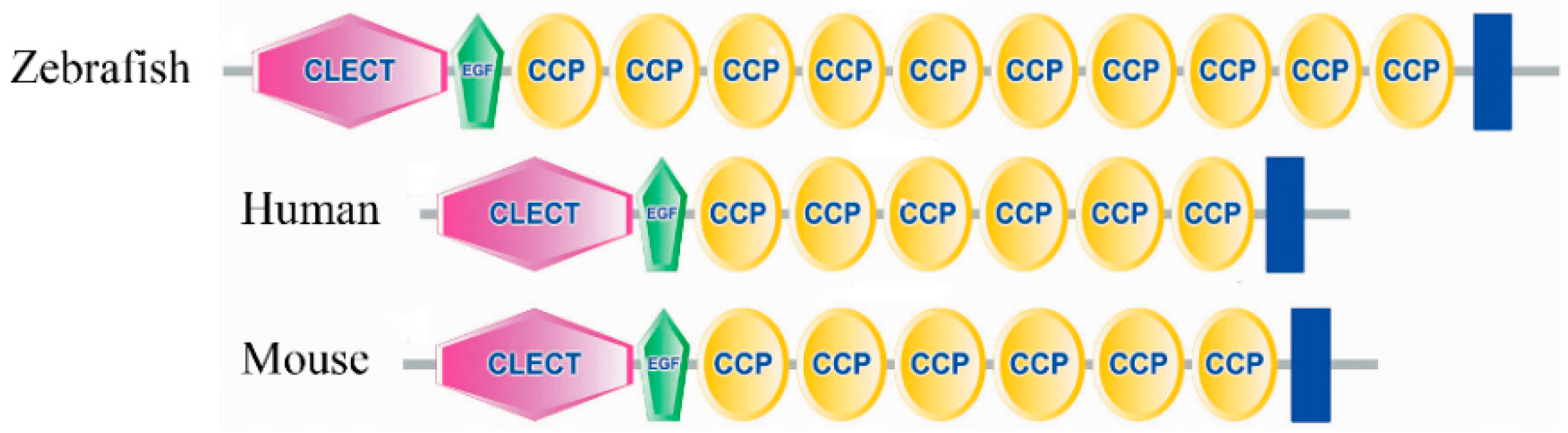
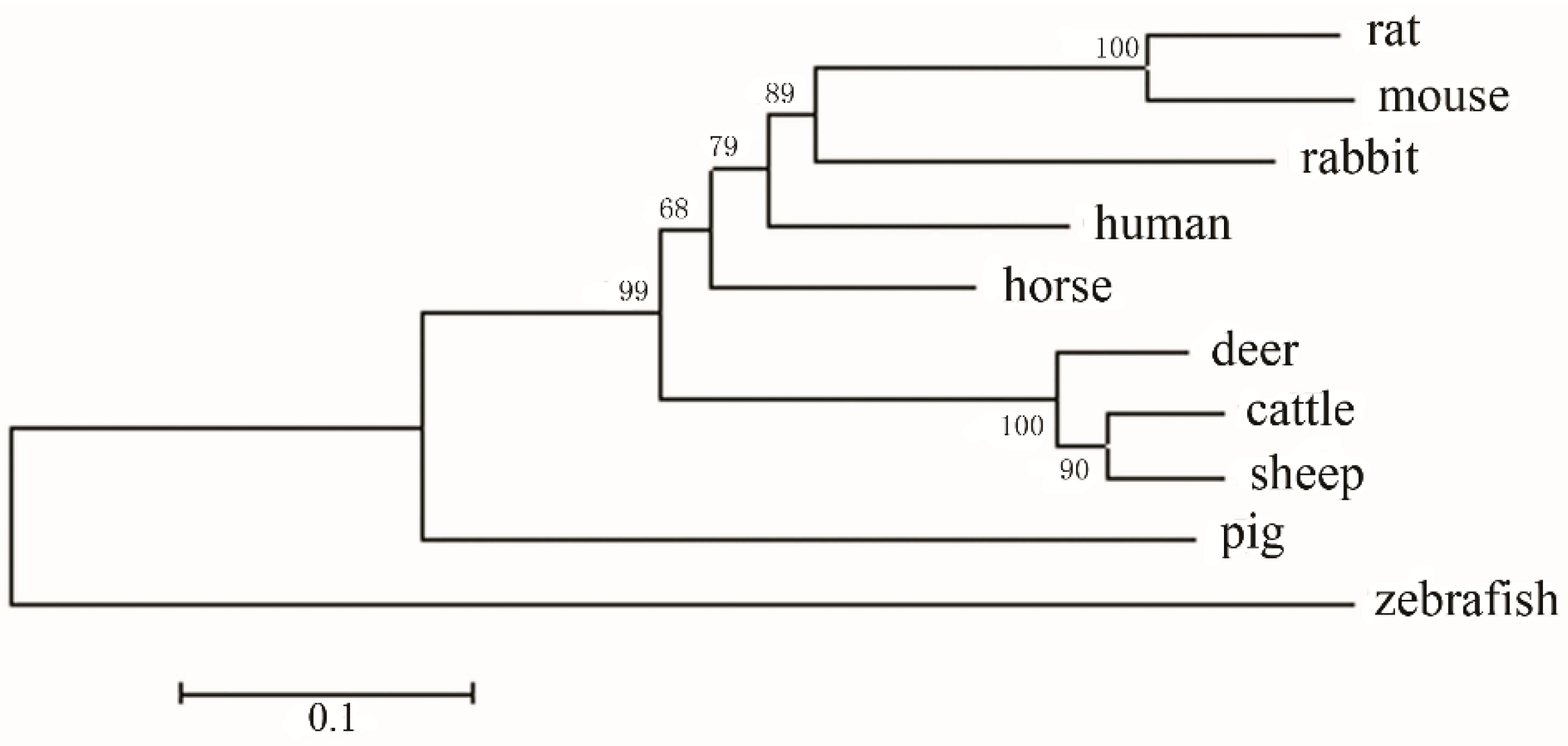
2.3. Amino Acid Sequence Analysis of Zebrafish E-Selectin



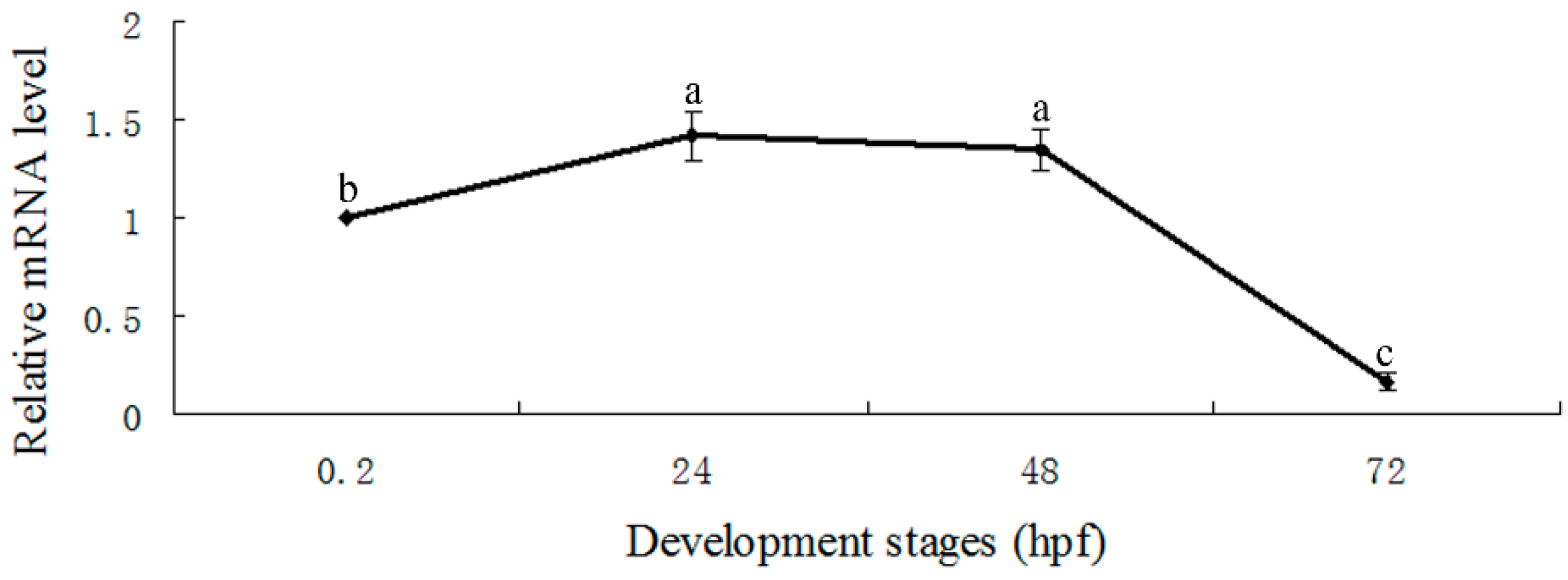
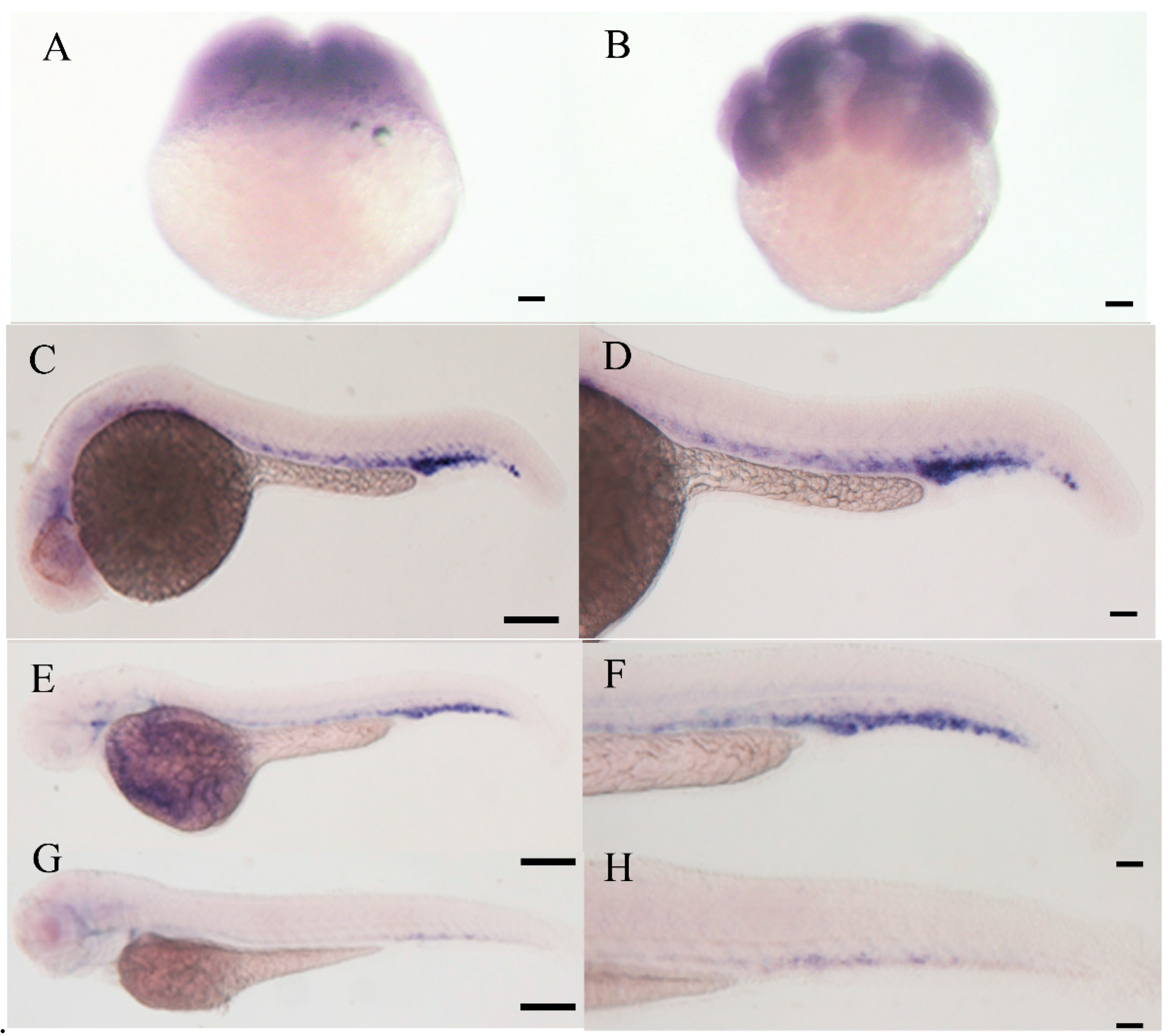
2.4. Expression Pattern of E-Selectin during Zebrafish Embryonic Development


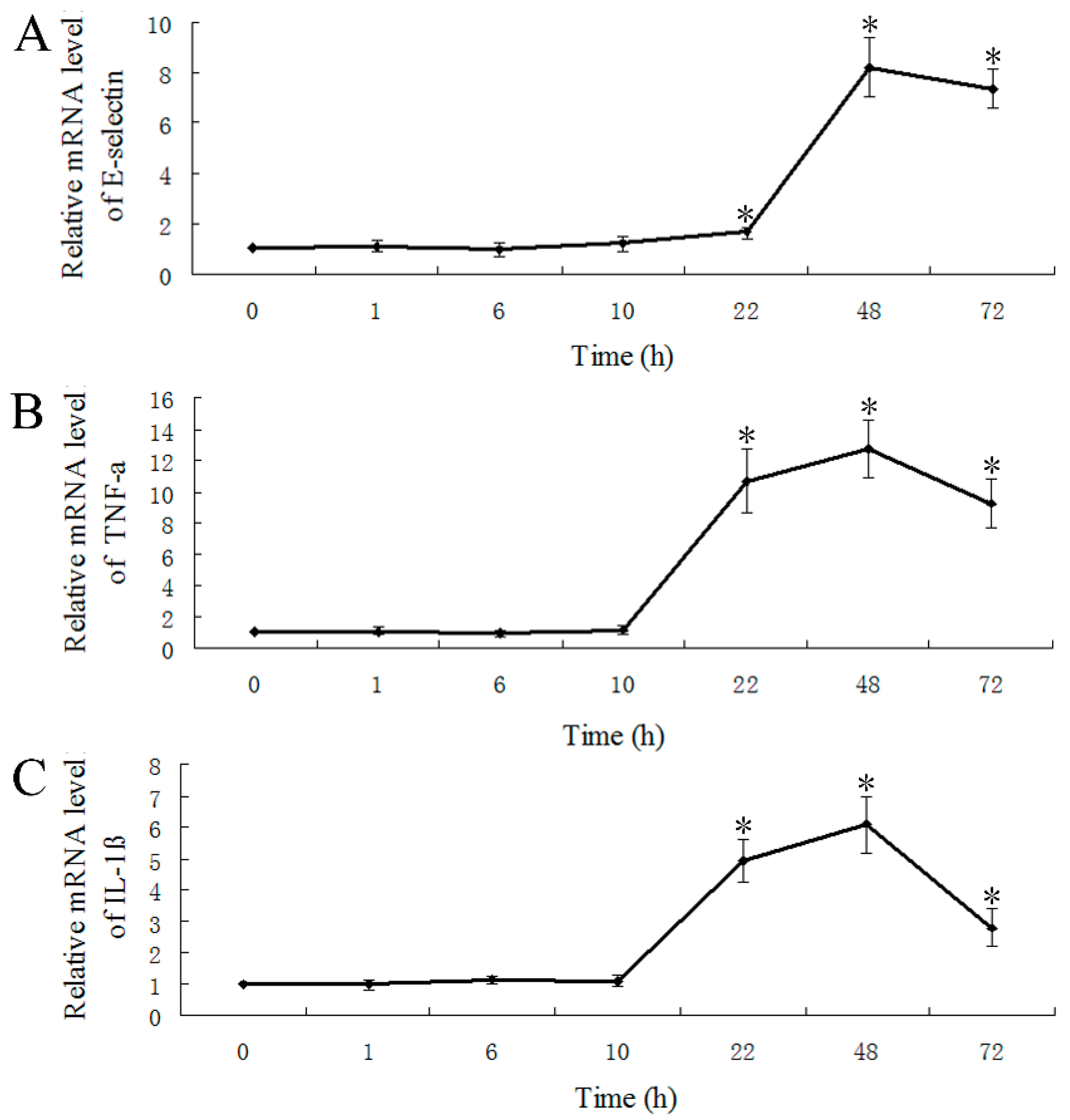
2.5. LPS-Induced Expression of E-Selectin, TNF-α and IL-1β

3. Experimental Section
3.1. Cloning of Zebrafish E-Selectin cDNA
3.2. Bioinformatics Analysis of Zebrafish E-Selectin
3.3. LPS Treatment
3.4. Real-Time Quantitative RT-PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (Acession Number) | Prime Sequences (5′–3′) | Product Size |
|---|---|---|
| β-actin (AF057040) | Forward: ATGGATGAGGAAATCGCTG | 130 |
| Reverse: ATGCCAACCATCACTCCCTG | ||
| E-selectin (KC488324) | Forward: GCAGCACTACACCGACTTGG | 196 |
| Reverse: TCCTTTGTTATTGGGCTCCTT | ||
| TNF-α (AY427649) | Forward: GCTGGATCTTCAAAGTCGGGTGTA | 138 |
| Reverse: TGTGAGTCTCAGCACACTTCCATC | ||
| IL-1β (AY340959) | Forward: TGGACTTCGCAGCACAAAATG | 124 |
| Reverse: GTTCACTTCACGCTCTTGGATG |
3.5. Whole-Mount in Situ Hybridization
3.6. Statistical Analysis
4. Conclusion
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
References
- Barthel, S.R.; Gavino, J.D.; Descheny, L.; Dimitroff, C.J. Targeting selectins and selectin ligands in inflammation and cancer. Expert Opin. Ther. Targets 2007, 11, 1473–1491. [Google Scholar] [CrossRef] [PubMed]
- Okanishi, H.; Kagawa, Y.; Watari, T. Expression of selectins and P-selectin glycoprotein ligand-1 in dogs with lymphocytic-plasmacytic enteritis. Vet. Immunol. Immunopathol. 2014, 161, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Bevilacqua, M.P.; Nelson, R.M. Selectins. J. Clin. Invest. 1993, 91, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Jebali, J.; Jeanneau, C.; Bazaa, A.; Mathieu, S.; El Ayeb, M.; Luis, J.; El Battari, A.; Marrakchi, N. Selectins as adhesion molecules and potential therapeutic target. Arch. Inst. Pasteur Tunis 2011, 88, 3–18. [Google Scholar]
- Impellizzeri, D.; Cuzzocrea, S. Targeting selectins for the treatment of inflammatory diseases. Expert Opin. Ther. Targets 2014, 18, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Bevilacqua, M.P.; Pober, J.S.; Mendrick, D.L.; Cotran, R.S.; Gimbrone, M.A., Jr. Identification of an inducible endothelial-leukocyte adhesion molecule. Proc. Natl. Acad. Sci. USA 1987, 84, 9238–9242. [Google Scholar] [CrossRef] [PubMed]
- Tsang, Y.T.; Stephens, P.E.; Licence, S.T.; Haskard, D.O.; Binns, R.M.; Robinson, M.K. Porcine E-selectin: Cloning and functional characterization. Immunology 1995, 85, 140–145. [Google Scholar] [PubMed]
- Huang, H.; Lavoie-Lamoureux, A.; Moran, K.; Lavoie, J.P. IL-4 stimulates the expression of CXCL-8, E-selectin, VEGF, and inducible nitric oxide synthase mRNA by equine pulmonary artery endothelial cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2007, 292, L1147–L1154. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.M.; Chapman, S.M.; Brown, A.A.; Frenette, P.S.; Hynes, R.O.; Wagner, D.D. The combined role of P- and E-selectins in atherosclerosis. J. Clin. Invest. 1998, 102, 145–152. [Google Scholar] [CrossRef]
- Ley, K. The role of selectins in inflammation and disease. Trends Mol. Med. 2003, 9, 263–268. [Google Scholar] [CrossRef]
- Liu, Z.J.; Tian, R.; An, W.; Zhuge, Y.; Li, Y.; Shao, H.; Habib, B.; Livingstone, A.S.; Velazquez, O.C. Identification of E-selectin as a novel target for the regulation of postnatal neovascularization: Implications for diabetic wound healing. Ann. Surg. 2010, 252, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Xu, X.; Bi, Y.; Xu, J.; Qin, C.; Han, M. Systemic inflammation promotes lung metastasis via E-selectin upregulation in mouse breast cancer model. Cancer Biol. Ther. 2014, 15, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Shirure, V.S.; Reynolds, N.M.; Burdick, M.M. Mac-2 binding protein is a novel E-selectin ligand expressed by breast cancer cells. PLoS ONE. 2012, 7, e44529. [Google Scholar] [CrossRef]
- Shirure, V.S.; Liu, T.; Delgadillo, L.F.; Cuckler, C.M.; Tees, D.F.; Benencia, F.; Goetz, D.J.; Burdick, M.M. CD44 variant isoforms expressed by breast cancer cells are functional E-selectin ligands under flow conditions. Am. J. Physiol. Cell Physiol. 2015, 308, C68–C78. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, P.L.; Huot, J.; Auger, F.A. Mechanisms by which E-selectin regulates diapedesis of colon cancer cells under flow conditions. Cancer Res. 2008, 68, 5167–5176. [Google Scholar] [CrossRef] [PubMed]
- Gout, S.; Tremblay, P.L.; Huot, J. Selectins and selectin ligands in extravasation of cancer cells and organ selectivity of metastasis. Clin. Exp. Metastasis 2008, 25, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Dooley, K.; Zon, L.I. Zebrafish: A model system for the study of human disease. Curr. Opin. Genet. Dev. 2000, 10, 252–256. [Google Scholar] [CrossRef]
- Painter, C.A.; Ceol, C.J. Zebrafish as a platform to study tumor progression. Methods Mol. Biol. 2014, 1176, 143–155. [Google Scholar] [PubMed]
- Asnani, A.; Peterson, R.T. The zebrafish as a tool to identify novel therapies for human cardiovascular disease. Dis. Model. Mech. 2014, 7, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, R.N.; Jopling, C.; van Eeden, F.J. Zebrafish as a model of cardiac disease. Prog. Mol. Biol. Transl. Sci. 2014, 124, 65–91. [Google Scholar] [PubMed]
- Larigan, J.D.; Tsang, T.C.; Rumberger, J.M.; Burns, D.K. Characterization of cDNA and genomic sequences encoding rabbit ELAM-1: Conservation of structure andfunctional interactions with leukocytes. DNA Cell Biol. 1992, 11, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Weller, A.; Isenmann, S.; Vestweber, D. Cloning of the mouse endothelial selectins. Expression of both E- and P-selectin is inducible by tumor necrosis factor alpha. J. Biol. Chem. 1992, 267, 15176–15183. [Google Scholar]
- Fries, J.W.; Williams, A.J.; Atkins, R.C.; Newman, W.; Lipscomb, M.F.; Collins, T. Expression of VCAM-I and E-selectin in an in vivo model of endothelial activation. Am. J. Parhol. 1993, 143, 725–737. [Google Scholar]
- Nguyen, M.; Strubel, N.A.; Bischoff, J. A role for sialyl Lewis-X/A glycoconjugates in capillary morphogenesis. Nature 1993, 365, 267–269. [Google Scholar] [CrossRef] [PubMed]
- Hedges, J.F.; Demaula, C.D.; Moore, B.D.; McLaughlin, B.E.; Simon, S.I.; MacLachlan, N.J. Characterization of equine E-selectin. Immunology 2001, 103, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Graves, B.J.; Crowther, R.L.; Chandran, C.; Rumberger, J.M.; Li, S.; Huang, K.S.; Presky, D.H.; Familletti, P.C.; Wolitzky, B.A.; Burns, D.K. Insight into E-selectin/ligand interaction from the crystal structure and mutagenesis of the lec/EGF domains. Nature 1994, 367, 532–538. [Google Scholar] [CrossRef]
- Kansas, G.S.; Saunders, K.B.; Ley, K.; Zakrzewicz, A.; Gibson, R.M.; Furie, B.C.; Furie, B.; Tedder, T.F. A role for the epidermal growth factor-like domain of P-selectin in ligand recognition and cell adhesion. J. Cell Biol. 1994, 124, 609–618. [Google Scholar] [CrossRef]
- McEver, R.P. Selectins: Novel receptors that mediate leukocyte adhesion during inflammation. Thromb. Haemost. 1991, 65, 223–228. [Google Scholar]
- Zhang, R.L.; Chopp, M.; Zhang, Z.G.; Phillips, M.L.; Rosenbloom, C.L.; Cruz, R.; Manning, A. E-selectin in focal cerebral ischemia and reperfusion in the rat. J. Cereb. Blood Flow Metab. 1996, 16, 1126–1136. [Google Scholar] [CrossRef]
- Bird, S.; Wang, T.; Zou, J.; Cunningham, C.; Secombes, C.J. The first cytokine sequence within cartilaginous fish: IL-1 beta in the small spotted catshark (Scyliorhinus canicula). J. Immunol. 2002, 168, 3329–3340. [Google Scholar] [CrossRef] [PubMed]
- Pressley, M.E.; Phelan, P.E.; Witten, P.E.; Mellon, M.T.; Kim, C.H. Pathogenesis and inflammatory response to Edwardsiella tarda infection in the zebrafish. Dev. Comp. Immunol. 2005, 29, 501–513. [Google Scholar] [CrossRef] [PubMed]
- Roca, F.J.; Mulero, I.; López-Muñoz, A.; Sepulcre, M.P.; Renshaw, S.A.; Meseguer, J.; Mulero, V. Evolution of the inflammatory response in vertebrates: Fish TNF-α is a powerful activator of endothelial cells but hardly activates phagocytes. J. Immunol. 2008, 181, 5071–5081. [Google Scholar] [CrossRef]
- BLAST server-Ensembl. Available online: http://www.ensembl.org/Danio_rerio/blastview (accessed on 8 October 2015).
- SMART. Available online: http://smart.embl-heidelberg.de (accessed on 8 October 2015).
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Thisse, C.; Thisse, B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nat. Protoc. 2008, 3, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are not available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, G.; Liu, K.; Wang, X.; Liu, X.; He, Q.; Hsiao, C.-D. Identification and Expression Analysis of Zebrafish (Danio rerio) E-Selectin during Embryonic Development. Molecules 2015, 20, 18539-18550. https://doi.org/10.3390/molecules201018539
Sun G, Liu K, Wang X, Liu X, He Q, Hsiao C-D. Identification and Expression Analysis of Zebrafish (Danio rerio) E-Selectin during Embryonic Development. Molecules. 2015; 20(10):18539-18550. https://doi.org/10.3390/molecules201018539
Chicago/Turabian StyleSun, Guijin, Kechun Liu, Xue Wang, Xiuhe Liu, Qiuxia He, and Chung-Der Hsiao. 2015. "Identification and Expression Analysis of Zebrafish (Danio rerio) E-Selectin during Embryonic Development" Molecules 20, no. 10: 18539-18550. https://doi.org/10.3390/molecules201018539