
Trypanocidal Activity of Long Chain Diamines and Aminoalcohols

 and
and
Abstract
:1. Introduction
2. Results and Dicussion
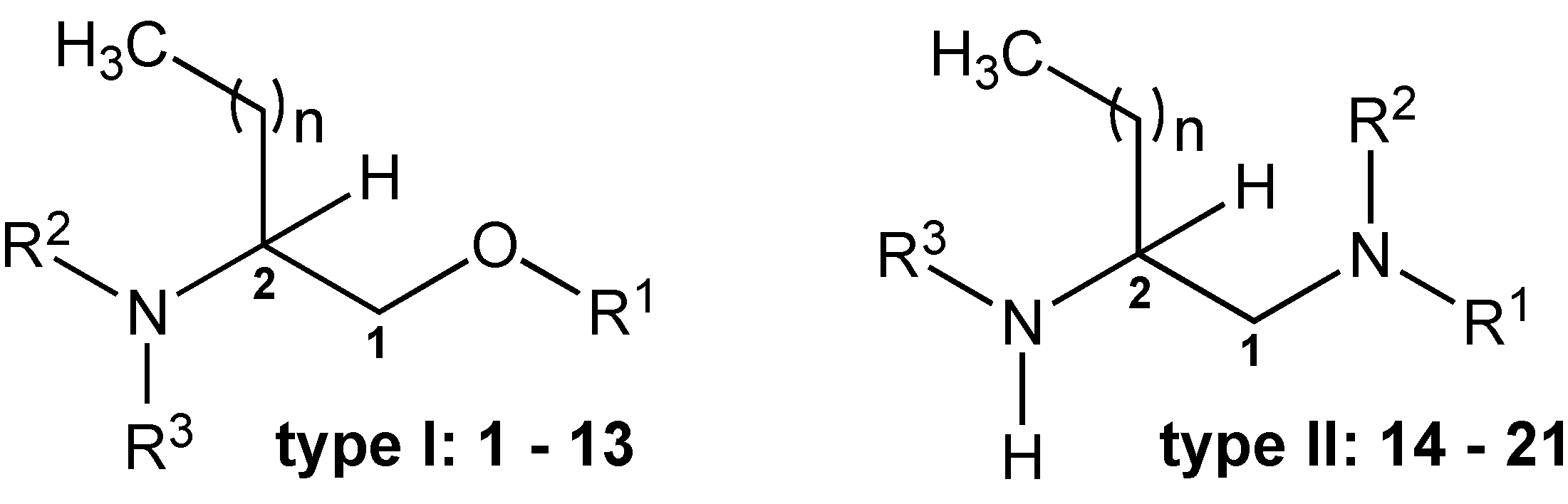
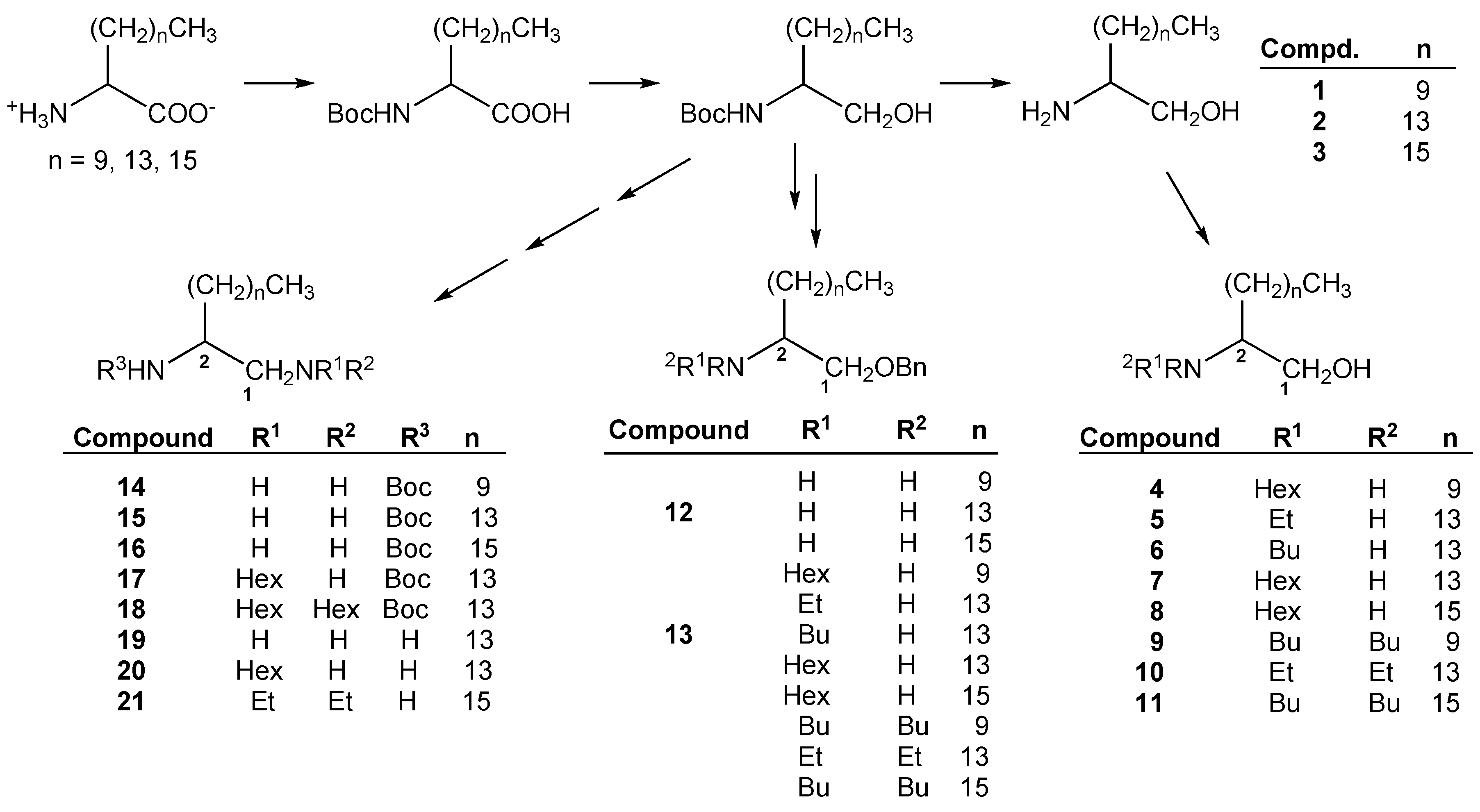
2.1. Chemistry

2.2. Biological Activity


{kind=link}
{kind=link}

| Compound | R1 | R2 | R3 | n | MG Strain | JEM Strain | Macrophages | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| IC50 (µM) | PNFX | SI | IC50 (µM) | PNFX | SI | LC50 (µM) | |||||
| 1 | H | H | H | 9 | 14.5 ± 3.15 | 1.5 | 1.0 | 33.0 ± 1.70 | 0.4 | 0.4 | 14.0 ± 0.90 |
| 2 | H | H | H | 13 | 23.7 ± 3.15 | 0.9 | 3.3 | 3.0 ± 1.50 * | 4.9 | 26 | 77.8 ± 0.35 |
| 3 | H | H | H | 15 | 46.2 ± 2.50 | 0.5 | 0.1 | 9.9 ± 1.65 * | 1.5 | 0.3 | 3.0 ± 0.24 |
| 4 | H | H | Hex | 9 | 21.1 ± 1.25 | 1.0 | 1.7 | 37.2 ± 1.25 | 0.4 | 1.0 | 36.1 ± 2.30 |
| 5 | H | H | Et | 13 | 21.6 ± 2.95 | 1.0 | 2.0 | 19.3 ± 1.35 | 0.8 | 2.3 | 43.4 ± 0.50 |
| 6 | H | H | Bu | 13 | 3.2 ± 0.20 * | 6.6 | 20 | 2.8 ± 0.75 * | 5.3 | 23 | 63.9 ± 3.50 |
| 7 | H | H | Hex | 13 | 6.1 ± 0.15 * | 3.5 | 1.3 | na | - | - | 8.2 ± 1.55 |
| 8 | H | H | Hex | 15 | na | - | - | na | - | - | 26.0 ± 0.20 |
| 9 | H | Bu | Bu | 9 | 39.2 ± 0.20 | 0.5 | 1.6 | 50.5 ± 0.70 | 0.3 | 1.3 | 63.7 ± 0.73 |
| 10 | H | Et | Et | 13 | na | - | - | na | - | - | 5.6 ± 0.27 |
| 11 | H | Bu | Bu | 13 | na | - | - | 50.3 ± 0.10 | 0.3 | - | nt |
| 12 | Bn | H | H | 13 | 34.4 ± 1.80 | 0.6 | 0.8 | 28.8 ± 1.20 | 0.5 | 1.0 | 27.7 ± 0.10 |
| 13 | Bn | H | Bu | 13 | na | - | - | na | - | - | nt |
| NFX | 21.2 ± 1.16 | 1.0 | 2.3 | 14.8 ± 1.24 | 1.0 | 3.3 | 49.1 ± 1.15 | ||||

| Compound | R1 | R2 | R3 | n | MG Strain | JEM Strain | Macrophages | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| IC50 (µM) | PNFX | SI | IC50 (µM) | PNFX | SI | LC50 (µM) | |||||
| 14 | H | H | Boc | 9 | 10.8 ± 0.0 * | 2.0 | 4.4 | 14.5 ± 0.05 | 1.0 | 3.3 | 47.1 ± 0.35 |
| 15 | H | H | Boc | 13 | 12.0 ± 2.15 | 1.8 | 4.0 | 20.4 ± 0.30 | 0.7 | 2.4 | 48.1 ± 0.15 |
| 16 | H | H | Boc | 15 | 13.9 ± 1.00 | 1.5 | 1.7 | 6.7 ± 2.10 | 2.2 | 3.6 | 24.1 ± 0.20 |
| 17 | Hex | H | Boc | 13 | 26.7 ± 0.25 | 0.8 | 0.2 | 30.6 ± 0.65 | 0.5 | 0.1 | 4.2 ± 0.05 |
| 18 | Hex | Hex | Boc | 13 | na | - | na | - | - | nt | |
| 19 | H | H | H | 13 | 19.2 ± 2.90 | 1.1 | 4.1 | 26.6 ± 0.70 | 0.6 | 2.9 | 78.1 ± 0.85 |
| 20 | Hex | H | H | 13 | 32.9 ± 1.25 | 0.6 | 0.2 | 48.8 ± 1.05 | 0.3 | 0.1 | 5.0 ± 0.10 |
| 21 | Et | Et | H | 13 | 42.2 ± 1.4 | 0.5 | 0.2 | na | - | - | 9.3 ± 0.50 |
| NFX | 21.2 ± 1.16 | 1.0 | 2.3 | 14.8 ± 1.24 | 1.0 | 3.3 | 49.1 ± 1.15 | ||||

| Compound | Epimastigotes | Amastigotes | Cytotoxicity | ||||
|---|---|---|---|---|---|---|---|
| CL-B5 Clone | CL-B5 Clone | NCTC-929 | |||||
| IC50 (µM) | PNFX | SI | IC50 (µM) | PNFX | SI | LC50 (µM) | |
| 2 | 3.5 ± 0.01 * | 3.0 | 7.2 | 1.8 ± 0.12 | 0.3 | 14.0 | 25.2 ± 0.12 |
| 5 | 3.3 ± 0.10 * | 3.1 | 9.8 | 0.6 ± 0.25 | 1.0 | 54.0 | 32.4 ± 0.02 |
| 6 | 4.2 ± 0.52 * | 2.5 | 22.7 | 2.0 ± 0.27 | 0.3 | 47.8 | 95.6 ± 1.19 |
| 14 | 2.5 ± 0.03 * | 4.2 | 11.8 | 3.0 ± 0.21 | 0.2 | 9.9 | 29.6 ± 0.01 |
| 15 | 2.4 ± 0.01 * | 4.3 | 10.6 | 2.6 ± 0.23 | 0.2 | 9.8 | 25.6 ± 0.06 |
| NFX | 10.4 ± 1.3 | 1.0 | 6.7 | 0.6 ± 0.01 | 1.00 | 116.0 | 69.6 ± 0.17 |
3. Experimental Section
3.1. General Information
3.2. Chemistry
3.2.1. 2-Aminohexadecanol (2)
3.2.2. 2-(Ethylamino)hexadecanol (5)
3.2.3. 2-(Butylamino)hexadecanol (6)
3.2.4. 2-(tert-Butoxycarbonyl)-dodecan-1,2-diamine (14)
3.2.5. 2-tert-Butoxycarbonyl-hexadecan-1,2-diamine (15)
3.3. In Vitro Assays
3.3.1. Parasite Strains, Culture and Assays Procedures
3.3.2. Macrophage and Fibroblast Cultures
3.3.3. Epimastigote Susceptibility Assay
3.3.4. Amastigote Susceptibility Assay
3.3.5. Cytotoxicity Assays
3.3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Coura, J.R.; Viñas, P.A. Chagas disease: A new worldwide challenge. Nature 2010, 465, S6–S7. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, D.E.; Benchimol, M.; Crepaldi, P.H.; de Souza, W. Interactive multimedia to teach the life cycle of Trypanosoma cruzi, the causative agent of Chagas disease. PLoS Negl. Trop. Dis. 2012, 6, e1749. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO): Geneva, Switzerland, Fact Sheet No 340. Available online: http:// www.who.int/mediacentre/factsheets/fs340/en/ (accessed on 16 April 2015).
- Rassi, A.; Rassi, A.; Marin-Neto, J.A. Chagas disease. Lancet 2010, 375, 1388–1402. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Molina, J.A.; Rodríguez-Guardado, A.; Soriano, A.; Pinazo, M.J.; Carrilero, B.; García-Rodríguez, M.; Salas, J.; Torrús, D.; Soler-Ferrer, C.; Puente, S.; et al. Guidelines on the treatment of chronic coinfection by Trypanosoma cruzi and HIV outside endemic areas. HIV Clin. Trials 2011, 12, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Bern, C. Chagas disease in the immunosuppressed host. Curr. Opin Infect Dis. 2012, 25, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Coura, J.R.; Borges-Pereira, J. What is known and what should be improved: A systemic review. Rev. Soc. Bras. Med. Trop. 2012, 45, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Matta Guedes, P.M.; Gutierrez, F.R.; Nascimento, M.S.; Do-Valle-Matta, M.A.; Silva, J.S. Antiparasitical chemotherapy in Chagas’ disease cardiomyopathy: current evidence. Trop. Med. Int. Health 2012, 17, 1057–1065. [Google Scholar] [CrossRef] [PubMed]
- Sosa-Estani, S.; Cura, E.; Velazquez, E.; Yampotis, C.; Segura, E.L. Etiological treatment of young women infected with Trypanosoma cruzi, and prevention of congenital transmission. Rev. Soc. Bras. Med. Trop. 2009, 42, 484–487. [Google Scholar] [CrossRef] [PubMed]
- Benaim, G.; Paniz Mondolfi, A.E. The emerging role of amiodarone and dronedarone in Chagas disease. Nat. Rev. Cardiol. 2012, 9, 605–609. [Google Scholar] [CrossRef] [PubMed]
- Urbina, J.A. Specific chemotherapy of Chagas disease: Relevance, current limitations and new approaches. Acta Trop. 2010, 115, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, S.; Milstien, S. Sphingosine-1-phosphate: Signaling inside and out. FEBS Lett. 2000, 476, 55–57. [Google Scholar] [CrossRef]
- Kester, M.; Kolesnick, R. Sphingolipids as therapeutics. Pharmacol. Res. 2003, 47, 365–371. [Google Scholar] [CrossRef]
- Lauer, S.A.; Ghori, N.; Haldar, K. Sphingolipid synthesis as a target for chemotherapy against malaria parasites. Proc. Natl. Acad. Sci. USA 1995, 92, 9181–9185. [Google Scholar] [CrossRef] [PubMed]
- Howarth, J.; Lloyd, D.G. Simple 1,2-aminoalcohols as strain-specific antimalarial agents. J. Antimicrob. Chemother. 2000, 46, 625–628. [Google Scholar] [CrossRef] [PubMed]
- Calas, M.; Ancelin, M.L.; Cordina, G.; Portefaix, P.; Piquet, G.; Vidal-Sailhan, V.; Vial, H. Antimalarial activity of compounds interfering with Plasmodium falciparum phospholipid metabolism: Comparison between mono- and bisquaternary ammonium salts. J. Med. Chem. 2000, 43, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Del Olmo, E.; Alves, M.; López, J.L.; Inchaustti, A.; Yaluff, G.; Rojas de Arias, A.; San Feliciano, A. Leishmanicidal activity of some aliphatic diamines and amino-alcohols. Bioorg. Med. Chem. Lett. 2002, 12, 659–662. [Google Scholar] [CrossRef]
- Rebollo, O.; del Olmo, E.; Ruiz, G.; López-Pérez, J.L.; Giménez, A.; San Feliciano, A. Leishmanicidal and trypanocidal activities of 2-aminocyclohexanol and 1,2-cyclohexanediamine derivatives. Bioorg. Med. Chem. Lett. 2008, 18, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Del Olmo, E.; Diaz-Gonzalez, R.; Escarcena, R.; Carvalho, L.; Bustos, L.A.; Navarro, M.; Feliciano, A.S. Diamine and aminoalcohol derivatives active against Trypanosoma brucei. Bioorg. Med. Chem. Lett. 2012, 22, 440–443. [Google Scholar] [CrossRef] [PubMed]
- Del Olmo, E.; Molina-Salinas, G.M.; Escarcena, R.; Alves, M.; Lopez-Perez, J.L.; Hernandez-Pando, R.; Said-Fernandez, S.; Feliciano, A.S. Simple dihydrosphyngosine analogues with potent activity against MDR-Mycobacterium tuberculosis. Bioorg. Med. Chem. Lett. 2009, 19, 5764–5768. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.D.; Duque, M.C.; Guhl, F. Phylogenetic reconstruction based on Cytochrome b (Cytb) gene sequences reveals distinct genotypes within Colombian Trypanosoma cruzi I populations. Acta Trop. 2011, 119, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Buckner, F.S.; Verlinde, C.L.; la Flamme, A.C.; van Voorhis, W.C. Efficient technique for screening drugs for activity against Trypanosoma cruzi using parasites expressing beta-galactosidase. Antimicrob. Agents Chemother. 1996, 40, 2592–2597. [Google Scholar]
- Tran, S.L.; Puhar, A.; Ngo-Camus, M.; Ramarao, N. Trypan blue dye enters viable cells incubated with the pore-forming toxin HlyII of Bacillus cereus. PLoS ONE 2011, 6, e22876. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Ayala, S.; Castrillón, J.A.; Palma, A.; Leal, S.M.; Escobar, P.; Bahsas, A. Synthesis, structural elucidation and in vitro antiparasitic activity against Trypanosoma cruzi and Leishmania chagasi parasites of novel tetrahydro-1-benzazepine derivatives. Bioorg. Med. Chem. 2010, 18, 4721–4739. [Google Scholar] [CrossRef] [PubMed]
- Fonseca-Berzal, C.; Escario, J.A.; Arán, V.J.; Gómez-Barrio, A. Further insights into biological evaluation of new anti-Trypanosoma cruzi 5-nitroindazoles. Parasitol. Res. 2014, 113, 1049–1056. [Google Scholar] [CrossRef] [PubMed]
- Muelas-Serrano, S.; Le-Senne, A.; Fernandez-Portillo, C.; Nogal, J.J.; Ochoa, C.; Gomez-Barrio, A. In vitro and in vivo anti-Trypanosoma cruzi activity of a novel nitro-derivative. Mem. Inst. Oswaldo Cruz 2002, 97, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Moraes, C.B.; Giardini, M.; Kim, H.; Franco, C.H.; Araujo-Junior, A.M.; Schenkman, S.; Chatelain, E.; Freitas-Junior, L.H. Nitroheterocyclic compounds are more efficacious than CYP51 inhibitors against Trypanosoma cruzi: Implications for Chagas disease drug discovery and development. Sci. Rep. 2014, 4, 4703–4714. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the reported compounds are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Legarda-Ceballos, A.L.; Del Olmo, E.; López-Abán, J.; Escarcena, R.; Bustos, L.A.; Fonseca-Berzal, C.; Gómez-Barrio, A.; Dib, J.C.; Feliciano, A.S.; Muro, A. Trypanocidal Activity of Long Chain Diamines and Aminoalcohols. Molecules 2015, 20, 11554-11568. https://doi.org/10.3390/molecules200611554
Legarda-Ceballos AL, Del Olmo E, López-Abán J, Escarcena R, Bustos LA, Fonseca-Berzal C, Gómez-Barrio A, Dib JC, Feliciano AS, Muro A. Trypanocidal Activity of Long Chain Diamines and Aminoalcohols. Molecules. 2015; 20(6):11554-11568. https://doi.org/10.3390/molecules200611554
Chicago/Turabian StyleLegarda-Ceballos, Ana L., Esther Del Olmo, Julio López-Abán, Ricardo Escarcena, Luis A. Bustos, Cristina Fonseca-Berzal, Alicia Gómez-Barrio, Juan C. Dib, Arturo San Feliciano, and Antonio Muro. 2015. "Trypanocidal Activity of Long Chain Diamines and Aminoalcohols" Molecules 20, no. 6: 11554-11568. https://doi.org/10.3390/molecules200611554
APA StyleLegarda-Ceballos, A. L., Del Olmo, E., López-Abán, J., Escarcena, R., Bustos, L. A., Fonseca-Berzal, C., Gómez-Barrio, A., Dib, J. C., Feliciano, A. S., & Muro, A. (2015). Trypanocidal Activity of Long Chain Diamines and Aminoalcohols. Molecules, 20(6), 11554-11568. https://doi.org/10.3390/molecules200611554