
Assessment of Chemical Impact of Invasive Bryozoan Pectinatella magnifica on the Environment: Cytotoxicity and Antimicrobial Activity of P. magnifica Extracts

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Elemental Analysis of P. magnifica Gel
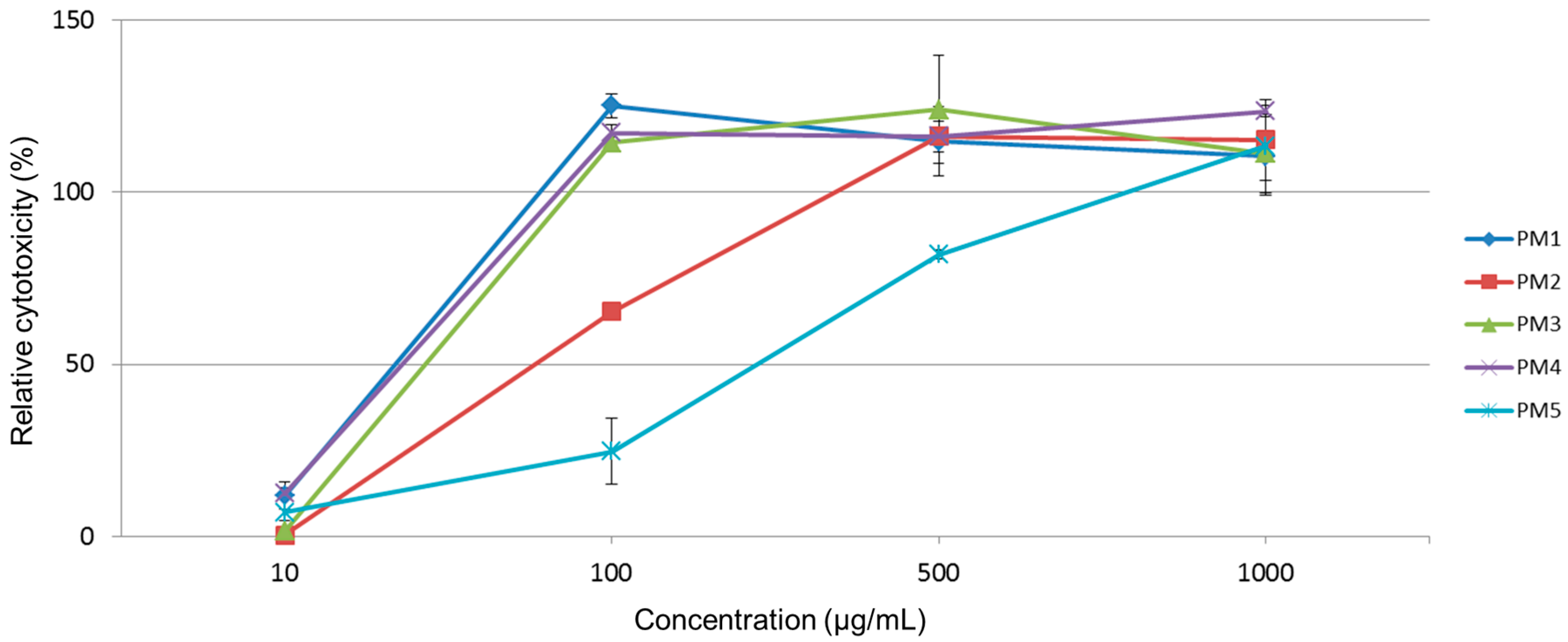
2.2. Cytotoxicity of Extracts
2.3. Antimicrobial Activity of Extracts
2.4. Cyanobacterial Toxins Determination
3. Discussion
3.1. Elemental Analysis of P. magnifica Gel
3.2. Cytotoxicity Tests
3.3. Antimicrobial Activity
3.4. Cyanobacterial Toxins Determination
4. Materials and Methods
4.1. Elemental Analysis of P. magnifica Gel
4.2. Cytotoxicity Tests
4.3. Antimicrobial Activity
4.4. Cyanobacterial Toxins Determination
4.4.1. Sample Preparation
4.4.2. MC Determination
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Carroget, P.; Carroget, L.; Gruet, Y.; Baudet, J.; Dutertre, M. Presence of colonies of the bryozoan Pectinatella magnifica Leidy 1851 in the Loire and the Nantes channel at Brest (Loire-Atlantique). Bull. Soc. Sci. Nat. Ouest. Fr. 2005, 27, 19–29. [Google Scholar]
- Wood, T.S. Bryozoans. In Ecology and Classification of North American Freshwater Invertebrates; Thorp, J.H., Covich, A.P., Eds.; Academic Press: New York, NY, USA, 2001; pp. 505–525. [Google Scholar]
- Kraepelin, K. Zur Biologie und Fauna der Süßwasserbryozoen. Zool. Anz. 1884, 7, 319–321. [Google Scholar]
- Balounová, Z.; Rajchard, J.; Švehla, J.; Šmahel, L. The onset of invasion of bryozoan Pectinatella magnifica in South Bohemia (Czech Republic). Biologia 2011, 66, 1091–1096. [Google Scholar] [CrossRef]
- Rodriguez, S.; Vergon, J.P. Pectinatella magnifica Leidy 1851 (Phylactolaemates), a species of bryozoa introduced in the north of Franche-Comté. Bull. Fr. Pech. Piscic. 2002, 365/366, 281–296. [Google Scholar] [CrossRef]
- Devin, S.; Bollache, L.; Noël, P.Y.; Beisel, J.-N. Patterns of biological invasions in French freshwater systems by non-indigenous macroinvertebrates. Hydrobiologia 2005, 551, 137–146. [Google Scholar] [CrossRef]
- Balounová, Z.; Pechoušková, E.; Rajchard, J.; Joza, V.; Šinko, J. World-Wide distribution of the Bryozoan Pectinatella magnifica (Leidy 1851). Eur. J. Environ. Sci. 2013, 3, 96–100. [Google Scholar]
- Szekeres, J.; Akác, A.; Csányi, B. First record of Pectinatella magnifica (Leidy 1851) in Hungary. Water Res. Manag. 2013, 3, 47–49. [Google Scholar]
- Notteghem, P. Évolution de la distribution de la Pectinatelle, Pectinatella magnifica (Leidy, 1851), Bryozoaire d’eau douce, en France et en Europe. Rev. Sci. Bourgogne-Nat. 2009, 9/10, 188–197. [Google Scholar]
- Hyunbin, J.; Gea-Jae, J.; Myeoungseop, B.; Dong-Gyun, H.; Jung-Soo, G.; Ji-Yoon, K.; Jong-Yun, C. Distribute pattern of Pectinatella magnifica (Leidy, 1851), an invasive species, in the Geum River and the Nakdong River, South Korea. J. Ecol. Environ. 2014, 37, 217–223. [Google Scholar]
- Oda, S. Pectinatella magnifica occurring in Lake Shoji, Japan. Proc. Jpn. Soc. Syst. Zool. 1974, 10, 31–39. [Google Scholar]
- Brown, C.J.D. A limnological study of certain fresh-water Polyzoa with special reference to their statoblasts. Trans. Am. Micros. Soc. 1933, 52, 271–314. [Google Scholar] [CrossRef]
- Seo, J.E. Taxonomy of the freshwater bryozoans from Korea. Korean J. Syst. Zool. 1998, 14, 371–378. [Google Scholar]
- Borg, F. Moostierchen oder Bryozoen (Ectoprocten). In Muschelinge oder Moluscocieda und Manteltiere oder Tunicata (Kamptozoa, Phoronidea, Bryozoa, Tunicata, Ascidiae). Die Tierwelt Deutschlands und der angrenzenden Meersteile nach ihren Merkmalen und nach ihrer Lebensweise; Bischoff, H., Ed.; Jena: Gustav Fisher, Germany, 1930; pp. 25–142. [Google Scholar]
- Prinsep, M.R.; Yao, B.; Nicholson, B.K.; Gordon, D.P. The pterocellins, bioactive alkaloids from the marine bryozoan Pterocella vesiculosa. Phytochem. Rev. 2004, 3, 325–331. [Google Scholar] [CrossRef]
- Kollar, P.; Rajchard, J.; Balounová, Z.; Pazourek, J. Marine natural products: Bryostatins in preclinical and clinical studies. Pharm. Biol. 2014, 52, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Dunlap, W.C.; Battershill, C.N.; Liptrot, C.H.; Coob, R.E.; Bourne, D.G.; Jaspars, M.; Long, P.F.; Newman, D.J. Biomedicinals from the phytosymbionts of marine invertebrates: A molecular approach. Methods 2007, 42, 358–376. [Google Scholar] [CrossRef] [PubMed]
- Figuerola, B.; Sla-Comorera, L.; Angulo-Preckler, C.; Vázquez, J.; Montes, M.J.; García-Aljaro, C.; Mercadé, E.; Blanch, A.R.; Avila, C. Antimicrobial activity of Antarctic bryozoans: An ecological perspective with potential for clinical applications. Mar. Environ. Res. 2014, 101, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Peters, L.; König, M.; Wright, A.D.; Pukall, R.; Stackebrandt, E.; Eberl, L.; Riede, K. Secondary metabolites of Flustra foliacea and their influence on bacteria. Appl. Environ. Microbiol. 2003, 69, 3469–3475. [Google Scholar] [CrossRef] [PubMed]
- Walls, J.T.; Ritz, D.A.; Blackman, A.J. Fouling, surface bacteria and antibacterial agents of four bryozoan species found in Tasmania, Australia. J. Exp. Mar. Biol. Ecol. 1993, 169, 1–13. [Google Scholar] [CrossRef]
- Sharp, K.H.; Davidson, S.K.; Haygood, M.G. Localization of ‘Candidatus Endobugula Sertula’ and the bryostatins throughout the life cycle of the bryozoan Bugula neritina. ISME J. 2007, 1, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Molinski, T.F.; Dalisay, D.S.; Lievens, S.L.; Saludes, J.P. Drug development from marine natural products. Nat. Rev. Drug Discov. 2009, 8, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Paul, V.J.; Arthur, K.E.; Ritson-Williams, R.; Ross, C.; Sharp, K. Chemical defenses: From Compounds to Communities. Biol. Bull. 2007, 213, 226–251. [Google Scholar] [CrossRef] [PubMed]
- Lopanik, N.; Gufson, K.R.; Lindquist, N. Structure of bryostatin 20: A symbiont-produced chemical. Defense for larvae of the host bryozoan, Bugula neritina. J. Nat. Prod. 2004, 67, 1412–1414. [Google Scholar] [CrossRef] [PubMed]
- Pejin, B.; Glamoclija, J.; Ciric, A.; Radotic, K.; Vajs, V.; Tesevic, V.; Hegedis, A.; Karaman, I.; Horvatovic, M.; Sokovic, M. Antimicrobial activity of the freshwater bryozoan Hyalinella punctata (Hancock, 1850). Dig. J. Nanomater. Biostruct. 2012, 7, 1021–1026. [Google Scholar]
- Pejin, B.; Ciric, A.; Horvatovic, M.; Jurca, T.; Glamoclija, J.; Nikolic, M.; Sokovic, M. An insight into antimicrobial activity of the freshwater bryozoan Pectinatella magnifica. Nat. Prod. Res. 2016, 30, 1839–1843. [Google Scholar] [CrossRef] [PubMed]
- Pejin, B.; Ciric, A.; Karaman, I.; Horvatovic, M.; Glamoclija, J.; Nikolic, M.; Sokovic, M. In vitro antibiofilm activity of the freshwater bryozoan Hyalinella punctata: A case study of Pseudomonas aeruginosa PAO1. Nat. Prod. Res. 2016, 30, 1847–1850. [Google Scholar] [CrossRef] [PubMed]
- Gad, S.C. Alternatives to in vivo studies in toxicology. In General and Applied Toxicology; Ballantyne, B., Marrs, T., Syversen, T., Eds.; Grove’s Dictionaries Inc.: New York, NY, USA, 1999; Volume 1, p. 178. [Google Scholar]
- Šetlíková, I.; Skácelová, O.; Šinko, J.; Rajchard, J.; Balounová, Z. Ecology of Pectinatella magnifica and associated algae and cyanobacteria. Biologia 2013, 68, 1136–1141. [Google Scholar] [CrossRef]
- Shine, R. The ecological impact of invasive cane toads (Bufo marinus) in Australia. Q. Rev. Biol. 2010, 85, 253–291. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.A.; Smith, F.M.; Heath, M.W.; Palfroy, T.; Gaw, S.; Young, R.G.; Ryan, K.G. Within-mat variability in anatoxin-a and homoanatoxin-a production among benthic Phormidium. (cyanobacteria) strains. Toxins 2012, 4, 900–912. [Google Scholar] [CrossRef] [PubMed]
- Morse, W. The chemical constitution of Pectinatella. Science (New York) 1930, 71, 265. [Google Scholar] [CrossRef] [PubMed]
- Pazourek, J.; Šmejkal, K.; Kollár, P.; Rajchard, J.; Šinko, J.; Balounová, Z.; Vlková, E.; Salmonová, H. Invasion of Pectinatella magnifica in Freshwater Resources of the Czech Republic. Int. J. Environ. Chem. Ecol. Geol. Geophys. Eng. 2016, 111, 278–285. [Google Scholar]
- Fajkusová, D.; Pesko, M.; Keltošová, S.; Guo, J.; Oktabec, Z.; Vejsová, M.; Kollár, P.; Coffey, A.; Csollei, J.; Králová, K.; et al. Anti-Infective and herbicidal activity of N-substituted 2-aminobenzothiazoles. Bioorg. Med. Chem. 2012, 20, 7059–7068. [Google Scholar] [CrossRef] [PubMed]
- Kos, J.; Zadražilová, I.; Peško, M.; Keltošová, S.; Tengler, J.; Goněc, T.; Bobáľ, P.; Kauerová, T.; Oravec, M.; Kollár, P.; et al. Antibacterial and herbicidal activity of ring-substituted 3-Hydroxynaphthalene-2-carboxanilides. Molecules 2013, 18, 7977–7997. [Google Scholar] [CrossRef] [PubMed]
- Bhunia, A.K. Foodborne Microbial Pathogens: Mechanisms and Pathogenesis; Springer: New York, NY, USA, 2008; p. 134. [Google Scholar]
- Silver, A.C.; Williams, D.; Faucher, J.; Horneman, A.J.; Gogarten, J.P.; Graf, J. Complex evolutionary history of the Aeromonas veronii group revealed by host interaction and DNA sequence data. PLoS ONE 2011, 6, e16751. [Google Scholar] [CrossRef] [PubMed]
- Sreedharan, K.; Philip, R.; Singh, I.S. Characterization and virulence potential of phenotypically diverse Aeromonas veronii isolates recovered from moribund freshwater ornamental fishes of Kerala, India. Anton Leeuw. 2013, 103, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Heindl, H.; Wiese, J.; Thiel, V.; Imhoff, J.F. Phylogenetic diversity and antimicrobial activities of bryozoan-associated bacteria isolated from Mediterranean and Baltic Sea habitats. Syst. Appl. Microbiol. 2010, 33, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Cronberg, G.; Annadotter, H. Manual on Aquatic Cyanobacteria. A Photo Guide and a Synopsis of Their Toxicology; ISSHA: Copenhagen, Denmark, 2006; p. 106. [Google Scholar]
- Glowacka, J.; Szefel-Markowska, M.; Waleron, M.; Łojkowska, E.; Waleron, K. Detection and identification of potentially toxic cyanobacteria in Polish water bodies. Acta Biochim. Pol. 2011, 58, 321–333. [Google Scholar] [PubMed]
- Quiblier, C.; Wood, S.; Echenique-Subiabre, I.; Heath, M.; Villeneuve, A.; Humbert, J.-F. A review of current knowledge on toxic benthic freshwater cyanobacteria—Ecology, toxin production and risk management. Water Res. 2013, 47, 5464–5479. [Google Scholar]
- Babica, P.; Kohoutek, J.; Bláha, L.; Adamovský, O.; Maršálek, B. Evaluation of extraction approaches linked to ELISA and HPLC for analyses of microcystin-LR, -RR and -YR in freshwater sediments with different organic material contents. Anal. Bioanal. Chem. 2006, 385, 1545–1551. [Google Scholar] [CrossRef] [PubMed]
- Faltermann, S.; Pretot, R.; Pernthaler, J.; Fent, K. Comparative effects of nodularin and microcystin-LR in zebrafish: 1. Uptake by organic anion transporting polypeptide Oatp1d1 (Slco1d1). Aquat. Toxicol. 2016, 171, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Chen, J.; Zhang, X.Z.; Xie, P. A review of reproductive toxicity of microcystins. J. Hazard. Mater. 2016, 301, 381–399. [Google Scholar] [CrossRef] [PubMed]
- Guidelines for Drinking Water Quality; World Health Organization: Geneva, Switzerland, 2004; p. 407. ISBN 92 4 154638 7.
- Kohoutek, J.; Adamovský, O.; Oravec, M.; Simek, Z.; Palíková, M.; Kopp, R.; Bláha, L. LC-MS analyses of microcystins in fish tissues overestimate toxin levels-critical comparison with LC-MS/MS. Anal. Bioanal. Chem. 2010, 398, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Bláhová, L.; Babica, P.; Maršálková, E.; Maršálek, B.; Bláha, L. Concentrations and seasonal trends of extracellular microcystins in freshwaters of the Czech Republic—Results of the national monitoring program. Clean-Soil Air Water 2007, 35, 348–354. [Google Scholar] [CrossRef]
- Bláhova, L.; Babica, P.; Adamovský, O.; Kohoutek, J.; Maršálek, B.; Bláha, L. Analyses of cyanobacterial toxins (microcystins, cylindrospermopsin) in the reservoirs of the Czech Republic and evaluation of health risks. Environ. Chem. Lett. 2008, 6, 223–227. [Google Scholar] [CrossRef]
- Jančula, D.; Straková, L.; Sadílek, J.; Maršálek, B.; Babica, P. Survey of cyanobacterial toxins in Czech water reservoirs - the first observation of neurotoxic saxitoxins. Environ. Sci. Pollut. Res. 2014, 21, 8006–8015. [Google Scholar] [CrossRef] [PubMed]
- Carey, C.C.; Haney, J.F.; Cottingham, K.L. First report of microcystin-LR in the cyanobacterium Gloeotrichia echinulata. Environ. Toxicol 2007, 22, 337–339. [Google Scholar] [CrossRef] [PubMed]
- Richardson, L.L.; Sekar, R.; Myers, J.L.; Gantar, M.; Voss, J.D.; Kaczmarsky, L.; Remily, E.R.; Boyer, G.L.; Zimba, P.V. The presence of the cyanobacterial toxin microcystin in black band disease of corals. FEMS Microbiol. Lett. 2007, 272, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Kaasalainen, U.; Fewer, D.P.; Jokela, J.; Wahlsten, M.; Sivonen, K.; Rikkinen, J. Cyanobacteria produce a high variety of hepatotoxic peptides in lichen symbiosis. Proc. Natl. Acad. Sci. USA 2012, 109, 5886–5891. [Google Scholar] [CrossRef] [PubMed]
- Bellinger, B.J.; Hagerthey, S.E. Presence and Diversity of Algal Toxins in Subtropical Peatland Periphyton: the Florida Everglades, USA. J. Phycol. 2010, 46, 674–678. [Google Scholar] [CrossRef]
- Testai, E.; Buratti, F.M.; Funari, E.; Manganelli, M.; Vichi, S.; Arnich, N.; Biré, R.; Fessard, V.; Sialehaamoa, A. Review and analysis of occurrence, exposure and toxicity of cyanobacteria toxins in food. EFSA Support. Publ. 2016, 13. [Google Scholar] [CrossRef]
- Ibelings, B.W.; Havens, K.E. Cyanobacterial toxins: A qualitative meta-analysis of concentrations, dosage and effects in freshwater, estuarine and marine biota. In Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs; Hudnell, H.K., Ed.; Springer: Berlin, Germany, 2008; pp. 675–732. [Google Scholar]
- Sieniawska, E.; Baj, T.; Dudka, J.; Gieroba, R.; Swiatek, L.; Rajtar, B.; Glowniak, K.; Polz-Dacewicz, M. Cytotoxicity, antioxidant activity and an effect on CYP3A4 and CYP2D6 of Mutellina purpurea L. extracts. Food Chem. Toxicol. 2013, 52, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Kmeť, V.; Drugdová, Z. Antimicrobial susceptibility of microflora from ovine cheese. Folia Microbiol. 2012, 57, 291–293. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not available.


{kind=link}
| Category | IC50 |
|---|---|
| Potentially very toxic | IC50 < 10 μg/mL |
| Potentially toxic | 10 μg/mL < IC50 < 100 μg/mL |
| Potentially harmful | 100 μg/mL < IC50 < 1000 μg/mL |
| Potentially non-toxic | IC50 ˃ 1000 μg/mL |
| Bacterial Strains | Minimal Inhibition Concentrations (mg/mL) | ||
|---|---|---|---|
| PM 1 * | PM 2 | PM 3 | |
| Micrococcus luteus ATCC 10240 | 10 | 4 | 4 |
| Acinetobacter parvus CCM 7030 | >20 | >20 | >20 |
| Bacillus cereus CCM 2010 | 5 | 4 | 2 |
| Bifidobacterium bifidum DSMZ 20215 | 10 | 10 | 5 |
| Clostridium difficile CCM 3593 | 1 | 0.5 | 0.5 |
| Clostridium perfringens CCM 4435 | >20 | 5 | 10 |
| Clostridium perfringens DSMZ 11778 | >20 | 5 | 2 |
| Enterobacter aerogenes CCM 7797 | >20 | >20 | >20 |
| Enterococcus faecalis DMND | >20 | >20 | >20 |
| Escherichia coli DMND | >20 | >20 | >20 |
| Escherichia coli O45 IS | >20 | >20 | >20 |
| Escherichia coli O55 IS | >20 | >20 | >20 |
| Lactobacillus brevis CCM 3805 | >20 | >20 | >20 |
| Listeria monocytogenes ATCC 7644 | 10 | 10 | 10 |
| Moraxella canis CCM 4590 | >20 | >20 | >20 |
| Propionibacterium acnes DSMZ 1893 | >20 | >20 | >20 |
| Pseudomonas aeruginosa CCM 1960 | >20 | >20 | 10 |
| Salmonella enterica Enteritidis ATCC 13076 | >20 | >20 | >20 |
| Salmonella enterica Typhimurium IS | >20 | >20 | >20 |
| Salmonella sp. DMND | >20 | >20 | >20 |
| Serratia marcescens DSMZ 30121 | >20 | >20 | >20 |
| Staphylococcus aureus ATCC 25923 | >20 | 10 | 5 |
| Location, Sampling Date * | MC–RR | MC–YR | MC–LR | MC–RR | MC–YR | MC–LR |
|---|---|---|---|---|---|---|
| P. magnifica Colony (ng/g d.w.) | Surrounding Water (ng/L) | |||||
| Veselí I, 23.7.2015 | 31.5 | 6.6 | 13.9 | 1.9 | <0.6 | 3.1 |
| Veselí I, 6.8.2015 | 6.1 | <0.6 | 10.6 | 38.3 | 17.9 | 70.4 |
| Hejtman, 23.7.2015 | 4.8 | <0.6 | 7.4 | 6.8 | 4.5 | 31.7 |
| Hejtman, 6.8.2015 | 4.0 | <0.6 | 2.6 | 121.6 | 27.5 | 159.5 |
| Hejtman, 6.8.2015 | 11.3 | 6.1 | 21.9 | n.a. | n.a. | n.a. |
| Hejtman, 9.10.2012 | 1.7 | <0.6 | 1.4 | n.a. | n.a. | n.a. |
| Hejtman, 31.10.2012 | 0.7 | <0.6 | 1.4 | n.a. | n.a. | n.a. |
| Hejtman, 9.10.2012 | <0.2 | <0.6 | <0.2 | n.a. | n.a. | n.a. |
| Hejtman, 9.10.2012 | <0.2 | <0.6 | <0.2 | n.a. | n.a. | n.a. |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kollar, P.; Šmejkal, K.; Salmonová, H.; Vlková, E.; Lepšová-Skácelová, O.; Balounová, Z.; Rajchard, J.; Cvačka, J.; Jaša, L.; Babica, P.; et al. Assessment of Chemical Impact of Invasive Bryozoan Pectinatella magnifica on the Environment: Cytotoxicity and Antimicrobial Activity of P. magnifica Extracts. Molecules 2016, 21, 1476. https://doi.org/10.3390/molecules21111476
Kollar P, Šmejkal K, Salmonová H, Vlková E, Lepšová-Skácelová O, Balounová Z, Rajchard J, Cvačka J, Jaša L, Babica P, et al. Assessment of Chemical Impact of Invasive Bryozoan Pectinatella magnifica on the Environment: Cytotoxicity and Antimicrobial Activity of P. magnifica Extracts. Molecules. 2016; 21(11):1476. https://doi.org/10.3390/molecules21111476
Chicago/Turabian StyleKollar, Peter, Karel Šmejkal, Hana Salmonová, Eva Vlková, Olga Lepšová-Skácelová, Zuzana Balounová, Josef Rajchard, Josef Cvačka, Libor Jaša, Pavel Babica, and et al. 2016. "Assessment of Chemical Impact of Invasive Bryozoan Pectinatella magnifica on the Environment: Cytotoxicity and Antimicrobial Activity of P. magnifica Extracts" Molecules 21, no. 11: 1476. https://doi.org/10.3390/molecules21111476