
Nutritional Regulators of Bcl-xL in the Brain

1
Department of Human Nutrition and Hospitality Management, College of Human Environmental Science, The University of Alabama, Tuscaloosa, AL 35487, USA
2
Department of Biological Sciences, College of Arts and Sciences, The University of Alabama, Tuscaloosa, AL 35487, USA
3
Department of Internal Medicine, Section of Endocrinology, Yale University, New Haven, CT 06520, USA
*
Author to whom correspondence should be addressed.
Molecules 2018, 23(11), 3019; https://doi.org/10.3390/molecules23113019
Submission received: 10 August 2018
/
Revised: 15 November 2018
/
Accepted: 18 November 2018
/
Published: 19 November 2018
(This article belongs to the Collection Neuroprotection Mediated by Natural Products and Their Chemical Derivatives)
Abstract
:B-cell lymphoma-extra large (Bcl-xL) is an anti-apoptotic Bcl-2 protein found in the mitochondrial membrane. Bcl-xL is reported to support normal brain development and protects neurons against toxic stimulation during pathological process via its roles in regulation of mitochondrial functions. Despite promising evidence showing neuroprotective properties of Bcl-xL, commonly applied molecular approaches such as genetic manipulation may not be readily applicable for human subjects. Therefore, findings at the bench may be slow to be translated into treatments for disease. Currently, there is no FDA approved application that specifically targets Bcl-xL and treats brain-associated pathology in humans. In this review, we will discuss naturally occurring nutrients that may exhibit regulatory effects on Bcl-xL expression or activity, thus potentially providing affordable, readily-applicable, easy, and safe strategies to protect the brain.
1. Introduction
The Bcl-2 family of proteins are best known for their role in apoptosis by means of mitochondrial membrane permeabilization and subsequent caspase regulation. Bcl-2 proteins are structurally similar in that they all share one or more of four Bcl-2 homology (BH) domains: BH2, BH1, BH3, and/or BH4. Generally, the Bcl-2 proteins are divided into two groups, anti-apoptotic and pro-apoptotic. Anti-apoptotic proteins such as Bcl-2 and Bcl-xL contain all four homologues. Among four BH domains, the BH4 domain has been shown to be essential for the anti-apoptotic properties of Bcl-2 and Bcl-xL by its ability to inactivate pro-apoptotic effector proteins such as Bax [1,2]. Mutation on the BH4 domain reduces stability of Bcl-2 protein and interferes occupation of Bax by Bcl-2 [3]. In addition to its ability to heterodimerize pro-apoptotic Bcl-2 proteins, the BH4 domain is reported to interact with the C-terminal of inositol 1,4,5-trisphosphate receptor (IP3R) inhibiting calcium-mediated apoptosis [4]. Pro-apoptotic proteins are further classified into two groups: multidomain and BH3-only. The multidomain pro-apoptotic proteins include Bax and Bak. Once activated by BH3-only proteins, Bax and Bak promote apoptosis by transitioning from inactive monomers to oligomers, leading to the permeabilization of the mitochondrial outer membrane. Additionally, Bax and Bak can be inhibited by the anti-apoptotic members of the Bcl-2 family [5]. BH3-only proteins are unique in their pro-apoptotic mechanisms; the activator such as Bid promotes oligomerization of Bax and Bak [6,7,8], whereas the sensitizers such as PUMA and Bad sequestrate anti-apoptotic Bcl-2 proteins favoring apoptotic signaling [9].
B-cell lymphoma-extra large (Bcl-xL) is a mitochondrially localized, anti-apoptotic member of the Bcl-2 protein family. Bcl-xL has been shown to bind with pro-apoptotic Bcl-2 proteins such as Bax, Bak and Bad, thereby preventing apoptotic signaling that leads to the opening of pro-apoptotic mitochondrial ion channels [10,11,12,13]. By doing so, Bcl-xL supports cell survival by inhibiting intrinsic death pathways such as cytochrome c release and apoptosome assembly [14,15,16]. Although Bcl-xL is best known for its pro-survival role in cancer cells, additional functions of Bcl-xL in the central nervous system (CNS) have been recently reported. Bcl-xL is highly expressed in various regions of the brain including hippocampus, cortex, and hypothalamus [17,18,19], and it is abundantly found in both neurons and glia [18,20,21,22]. During prenatal development, various Bcl-2 proteins, such as Bcl-2, Bax, and Bcl-xL are expressed in the brain. However, protein levels of Bcl-xL remain high after birth through adulthood, thus Bcl-xL may be the critical player to protect the adult brain [23]. Bcl-xL enhances intracellular energy metabolism by supporting the activity of the F1Fo ATP synthase [24]. Energy conservation by actions of Bcl-xL may prevent neuronal injury caused by oxygen or glucose deficits during pathological processes of the brain. Bcl-xL is also reported to promote formation of synapses and recycling of synaptic vesicles, both of which are necessary for neuronal plasticity and recovery [25,26,27]. Bcl-xL is required for the extension and branching of neurites which is essential for both normal brain development and brain recovery after a neurotoxic event [21]. Due to its importance in brain functions, alteration of Bcl-xL activities or expression levels is highly associated with pathological processes in the brain (Table 1) [28,29].
In contrast to its protective role, Bcl-xL is also responsible for causing neuronal death during neurotoxic challenge. Bcl-xL undergoes caspase 3-dependent N-terminal cleavage forming ∆N-Bcl-xL (Figure 1) [65]. Loss of CA1 hippocampal neurons after cerebral ischemia is highly associated with the accumulation of ∆N-Bcl-xL in vivo [18,66,67]. Additionally, excitotoxicity, the major neuronal death signal during cerebral stroke caused by a surge of glutamate, leads to an abundance of ∆N-Bcl-xL in primary neurons in vitro [68]. Approaches that inactivate ∆N-Bcl-xL or prevent the accumulation of ∆N-Bcl-xL using cleavage resistant animal models, delivering pharmacological inhibitors, and applying ischemic preconditioning protect neurons by improving mitochondrial function against neurotoxic stimuli [18,66,67,68,69,70].
In addition to its role regulating the function of brain cells, Bcl-xL participates in intracellular signaling by interacting with various protein partners during physiological and pathological processes in cells [29]. Bcl-xL directly binds to the subunit of F1Fo ATP synthase [14,24]. Formation of a Bcl-xL-F1Fo ATP synthase complex prevents a proton leak through the c-subunit ring of F1Fo ATP synthase and increases intracellular ATP production [24,71]. Maintaining a functional population of neurons is energy-demanding: neurotransmission consumes a large amount of energy. Actin and tubulin polymerization during neuronal outgrowth require ATP and GTP, respectively. Therefore, Bcl-xL-dependent augmentation of neuronal energy status favors neurite outgrowth [21], synapse formation [26], and protection against ischemia-induced energy deficit [18,66].
Bcl-xL also directly binds to the IP3R located on the endoplasmic reticulum (ER) [72,73]. Opening of IP3R releases calcium from the ER to mitochondria regulating mitochondrial energy metabolism. Mitochondrial calcium influx controls activities of dehydrogenases such as pyruvate dehydrogenase [74,75,76], thus regulating mitochondrial NADH. Abundance of mitochondrial NADH increases availability of electrons supporting operation of the electron transport chain and synthesis of ATP. Studies have reported that calcium directly binds to F1Fo ATP synthase and regulates ATP production [77,78,79], and binding between calcium in large quantities and ATP synthase may also produce pathological mitochondrial membrane depolarization [80]. Together, the proximity between mitochondria and the ER regulates mitochondrial energy metabolism which is in turn regulated by Bcl-xL. This may further influence synaptic transmission and neuronal development. In addition, augmentation of ATP synthesis via formation of a Bcl-xL-F1Fo ATP synthase complex may influence IP3R activities. Since IP3R contains ATP binding sites and opening of the IP3R is regulated by different concentrations of ATP [81,82], Bcl-xL may also be responsible for controlling IP3R activation through enhanced production of ATP at sites closely apposed to the ER in the mitochondrial associated membrane (MAM).
In addition to Bcl-xL and IP3R interaction, mitochondrial calcium influx may be regulated by Bcl-xL dependent voltage-dependent anion channel (VDAC) activities [83,84,85]. VDAC is expressed on the mitochondrial outer membrane, and it is reported to have multiple protein binding partners such as the adenine nucleotide translocator (ANT), hexokinase, and Bcl-2 family proteins. Formation of a multiprotein complex or oligomerization with other VDAC channels allows transport of ATP, ADP, NADH, metabolites (e.g., pyruvate, succinate), ions (e.g., Mg2+, and Ca2+), and cytochrome c [86,87]. Various groups have shown an interaction between Bcl-xL and VDAC [83,85,88,89], and formation of a Bcl-xL-VDAC complex can either enhance or inhibit activity of VDAC [90,91]. GST pulldown assay showed that VDAC 1 and 3 directly bind to both recombinant and endogenous Bcl-xL protein, and this interaction was associated with increases of mitochondrial calcium influx [83], whereas binding of the Bcl-xL BH4 domain with VDAC inhibited mitochondrial calcium uptake [85] via a reduction of channel conductance which may be associated with protection against apoptotic signaling [88].
Beclin-1, a critical player initiating autophagy [92,93,94], contains a BH3 domain. The crystal structure demonstrated a direct interaction between Beclin-1 and Bcl-xL, and occupation of Beclin-1 in the hydrophobic groove of Bcl-xL [95]. Dissociation of Bcl-xL and Beclin-1 induces autophagy in cancer cells [96], and mutation of the BH3 domain in Beclin-1 alters Bcl-xL-mediated autophagy [97] indicating regulatory roles of Bcl-xL in cell degradation. Interestingly, co-immunoprecipitation studies reveal that Bcl-xL also serves as a binding partner of PTEN-induced putative kinase 1 (PINK1), a mitochondrial serine/threonine kinase responsible for inducing mitochondrial autophagy (mitophagy) [98]. PINK1 phosphorylates ubiquitin, promotes recruitment of PARKIN and LC3, thus PINK1 facilitates formation of the autophagosome [98,99,100]. Alteration of PINK1 is highly associated with Parkinson’s disease [101,102] due to failure of mitochondrial quality control such as accumulation of impaired mitochondria. Although it is unknown how formation of a Bcl-xL-PINK1 complex directly influences mitophagy, PINK1 exhibits neuroprotective effects by phosphorylating serine 62 of Bcl-xL under apoptotic stimulation. PINK1-mediated phosphorylation of Bcl-xL inhibits N-terminal cleavage of Bcl-xL, thus preventing accumulation of neurotoxic ΔN-Bcl-xL [103].
Given these findings, Bcl-xL may be a necessary molecular player to maintain normal brain cell function, and it may be an important contributor to the delay and prevention of neurodegenerative processes [28,29]. Although it is necessary to pinpoint specific pathways of Bcl-xL-mediated neuroprotection using molecular approaches, currently reported strategies to manipulate Bcl-xL gene or protein may have limitations to translate into human subjects: Application of the pharmacological inhibitor (e.g., ABT-737) showed strong neuroprotective effects by blocking accumulation of ΔN-Bcl-xL but this drug does not cross the blood-brain barrier [18,68,104]. Overexpression of Bcl-xL improves neuronal survival [30,105] and Bcl-xL cleavage resistant animals protect the brain against ischemic injury [18], but genetic modification is still challenging to apply in human subjects. Currently, there are no FDA approved medications or treatments that specifically prevent Bcl-xL loss or promote Bcl-xL retention in human brain. In this review, we aim to introduce applicable strategies, nutritional intervention, that potentially regulate Bcl-xL protein, gene, or activity in the brain. Although limited information is available to explain a direct relationship between nutrients and Bcl-xL at the current time (Table 2), nutritional intervention may be an important extension in the field of Bcl-xL research due to its advantage in availability, low-toxicity, and cost-effectiveness. Here, we will discuss nutritional candidates that may participate in intracellular signaling pathways to delay degeneration of neurons or to promote recovery after neurotoxic insults via controlling expression of Bcl-xL.
2. Soy and Soy Isoflavones
Isoflavones are found predominantly in the legume family of flowering plants. In the human diet, soy isoflavones are often consumed in the form of soy or soy products, with the primary isoflavones being genistein, daidzein, and glycitein [128,129]. Soy isoflavones are called phytoestrogens due to chemical similarity with 17β-estradiol [130]. Studies have shown that soy isoflavones exert moderate levels of hormone-like effects or antagonize the interaction between estrogen and the estrogen receptor [131,132].
Soy isoflavones are reported to cross the blood-brain barrier [133]. Various research groups have reported improvement of the anti-apoptotic Bcl-2 protein profile in the brain after consumption of a soy diet or soy isoflavones in neurodegenerative models such as stroke, Alzheimer’s, and Parkinson’s disease [107,108,134,135,136]. Supplementation with soy isoflavones supports neuronal survival via enhancing Bcl-xL mRNA or protein expression in rodent brains [106,107,108,137]. Lovekamp-Swan et al. showed that rats supplemented with 600 µg/g soy isoflavones for 2 weeks prior to induction of middle carotid artery occlusion (MCAO), an experimental model to mimic focal ischemia, reduced MCAO-induced infarct size and protected brain cells from apoptotic death [107]. This group also showed that animals fed with a high soy diet had significantly enhanced mRNA and protein levels of Bcl-xL, without increasing Bcl-2 expression [107]. Interestingly, a high soy diet downregulated active caspase 3, the major contributor to the conversion of anti-apoptotic Bcl-xL to pro-apoptotic ∆N-Bcl-xL. Thus, oral supplementation of soy may prevent apoptotic pathways by preventing accumulation of pro-death ΔN-Bcl-xL and enhancing production of pro-survival Bcl-xL. Although limited information exists explaining the direct mechanisms of soy isoflavone-mediated Bcl-xL upregulation, soy isoflavone may interact with estrogen response elements (ERE) controlling Bcl-xL transcription. The bcl-x gene, BCL2L1, has been determined to contain an ERE at the 3′ untranslated region, indicating the target of transcriptional regulation via estrogen [138]. As soy isoflavones and 17β-estradiol both bind to estrogen receptor β [131,139], soy isoflavones may enhance Bcl-xL expression through transcriptional regulation by targeting EREs on BCL2L1.
In addition, Jiang et al. orally delivered various concentrations (20, 40, and 80 mg/kg body weight) of genistein, an abundantly found isoflavone in legumes including soybeans, to rat pups for 12 days and studied the effects of genistein in isoflurane-induced neurodegeneration [137]. Isoflurane inhalation increased fluoro-jade positive degenerating neurons in the hippocampal CA1, CA3, and dentate gyrus regions whereas genistein treatment significantly protected from this neuronal loss in dose dependent manner. Animals supplemented with genistein showed upregulation of Bcl-xL and Bcl-2, and down regulation of Bad and Bax protein levels in the brain leading to improvement in cognitive behaviors [137]. This group further found that genistein treatment significantly increased AKT and phosphorylated AKT protein levels [137]. AKT, protein kinase B, is a serine/threonine kinase that is responsible for the phosphorylation of pro-apoptotic Bad and Bax [140,141,142]. The phosphorylation status of Bad and Bax influences heterodimerization with anti-apoptotic Bcl-2 proteins such as Bcl-xL [141,143].
Orally delivered soy isoflavones prevented loss of Bcl-xL protein in the rat brain after intraperitoneal injection of amyloid-β peptides (Aβ) indicating the therapeutic potential of soy isoflavones in the treatment of patients with Alzheimer’s disease [108]. In this study, rats were gavaged with soybean isoflavone (80 mg/kg/day) for 14 days, then Bcl-xL protein levels were quantified using brain tissue. Soybean isoflavone fed group were resistant against Aβ-mediated Bcl-xL loss. Although molecular mechanisms of soy isoflavone-mediated neuroprotection against Aβ-induced oxidative damage are unclear, upregulation of Bcl-xL protein may be beneficial to block activation of Aβ. Conversion of amyloid precursor protein (APP) to active Aβ peptides requires proteolytic cleavage by proteases such as secretases and caspases [144,145,146,147]. Since Bcl-xL binds to the apoptosome and blocks activation of down-stream caspases in the intrinsic apoptotic pathway [148,149], increasing Bcl-xL protein expression by soy isoflavone may further arrest caspase-dependent post-translational modification of APP.
3. Ginseng and Ginsenosides
Panax ginseng has been used as a traditional medicine in Asia. Ginseng contains carbohydrates, oils, and amino acids. Most importantly, steroid derivatives of ginseng, ginsenosides (also called ginseng saponin), are reported to be the pharmaceutically functional molecule [150,151]. Antioxidant, anti-inflammatory, and anticancer effects of ginseng have been reported by various laboratories [152,153,154,155]. Moreover, neuroprotective effects of ginseng have been widely documented using both in vivo and in vitro models. Panaxadiol saponins of gisenoside Rb1 have been shown to decrease the loss of interneurons, astrocytes, and microglia in rats treated with kainic acid, an agonist of the AMPA/kainate glutamate receptor, thus preventing kainite-induced experimental seizures [156]. Further research suggests that ginsenoside Rg1 also decreased the MCAO-induced infarct volume in rat brains, indicating therapeutic effects of ginseng to prevent cerebral ischemia [157]. Interestingly, ginseng exerts neuroprotective properties in a concentration dependent manner. A low concentration of panaxatriol saponin treatment attenuated the loss of dopaminergic neurons in zebrafish whereas a high concentration caused neurotoxicity [158]. In vitro approaches using SH-SY5Y cell lines showed similar findings [159]. Ginsenoside Rd significantly increased cell viability at both 1 and 10 µM concentrations against in Parkinson’s Disease models, but it failed to do so at a higher concentration of 50 µM. At both lower concentrations, a decrease in oxidative stress and an increase in superoxide dismutase activity was recorded [159]. Therefore, it is possible that a low or a high concentration of ginsenosides may protect brain cells or facilitate death in malignant cells, respectively.
Upregulation of Bcl-xL protein by ginseng administration has been reported in various brain-associated disease models including bacterial inflammation, cerebral ischemia, and spinal cord injury [109,110,111,112,113]. Zhang et al. has reported that ginsenoside Rb1 protects the brain against ischemic insult in a Bcl-xL dependent manner [109]. Rats underwent intravenous infusion of ginsenoside Rb1 (6 or 60 µg/µL) for 4 weeks after induction of MCAO. Ginsenoside Rb1 treated animals showed an upregulation of Bcl-xL mRNA and protein in the stroke-affected cortex which was associated with a decrease in cortical infarct size, attenuated apoptosis, and improved navigational ability. This group identified the signal transducer and activator of transcription 5 (STAT5) as being under the control of ginsenoside Rb1-mediated Bcl-xL gene regulation. STAT5 contains a DNA binding domain and acts as a transcription factor; its target genes include Bcl-xL [160]. A Bcl-xL gene with a mutation in the STAT5 response element (STRE) failed to be activated by ginsenoside Rb1, whereas ginsenoside Rb1 treatment increased wild-type Bcl-xL activity by greater than 2 fold. In addition to prevention of cerebral ischemia, Bcl-xL-mediated neuroprotective effects of ginsenoside have been observed in spinal cord injury models [110,111,113] where red ginseng extract administration attenuated the expression of inflammatory cytokines, prevented morphological alteration of the spinal cord and improved rearing and locomotion activity. This group further showed that the neuroprotective properties of red ginseng extract are associated with its ability to upregulate protein levels of Bcl-xL and vascular endothelial growth factor (VEGF) [111,113]. Although the detailed mechanisms were not tested in these studies, VEGF has been reported to be a downstream target of Bcl-xL via the mitogen-activate protein kinase (MAPK) signaling pathway [161]; thus Bcl-xL may promote recovery after CNS injuries by supporting angiogenesis. In addition to transcriptional and translational regulation, ginsenosides may directly bind to Bcl-xL and control its activity. An in silico docking study predicted the interaction between Bcl-xL and ginsenosides, especially with Rg1, Rg2, Rg3, and Rd [162], and Ala142, Glu153, Thr109, Lys16, and Lys20 are reported as the predicted docking sites. In order to show biological significance of ginsenoside docking, verification of the interaction between Bcl-xL and ginsenosides using cells or in tissues will be required in future studies.
4. Omega-3 Fatty Acids
Docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA) are omega-3 (ω-3) fatty acids with 22-carbons and 20-carbons, respectively. Both fatty acids are found in fatty fish such as salmon, trout, and tuna. EPA and DHA are typically available in food or appropriate supplements, but they are also principle metabolites of the essential fatty acid α-linolenic acid. DHA and EPA are structural components of neuronal membranes, thus sufficient intake of omega-3 supports normal brain development [163,164], whereas deficiencies impair neuronal growth and plasticity [165,166,167].
Both in in vitro and in vivo models, application of omega-3 fatty acids has been noted to exert neuroprotection by enhancing the levels of Bcl-xL protein and modifying other Bcl-2 family protein levels [114,115,116,117,118,119,120,168,169]. Fat-1 gene encodes omega-3 fatty acid desaturase, the enzyme responsible for conversion of omega-6 fatty acids to omega-3 fatty acids. Tissues from Fat-1 mice are rich in omega-3 fatty acids such as EPA and DHA [170,171]. Shi et al. reported that cortical neurons isolated from Fat-1 mice were resistant to oxygen glucose deprivation (OGD)-mediated oxidative stress because Bcl-xL levels did not decline [118]. This group also showed that treatment with DHA (10 µM) significantly enhanced Bcl-xL protein in primary neuronal cultures indicating a regulatory effect of both endogenous and exogenous omega-3 fatty acids on Bcl-xL expression [118]. In addition, Fat-1 mice were protected from neuronal loss caused by infusion of amyloid beta (Aβ) 1-42, and primary cortical neurons isolated from Fat-1 embryos retained significantly higher levels of Bcl-xL protein under Aβ challenge [120].
Deficiency of thyroid hormone causes neuronal apoptosis associated with downregulation of Bcl-xL protein [116,172,173]. In a model of experimental hypothyroidism in pregnant rats, animals were protected from neuronal apoptosis by prior supplementation with 300 mg of omega-3 fatty acids [116]. Hypothyroid pups born from omega-3 fatty acid fed dams retained cerebellar weight and Bcl-xL protein in the cerebellum [116]. This group further showed that supplementation of omega-3 fatty acid inactivates c-Jun N-terminal kinase (JNK) and enhances phosphorylation of AKT and ERK [116]. AKT and ERK may enhance phosphorylation of pro-apoptotic Bcl-2 proteins [140,143] favoring survival by preventing dimerization of Bcl-xL. Interestingly, treatment with neuroprotectin D1, a derivative of omega-3 fatty acids, reverses phosphorylation of Bcl-xL by enhancing protein phosphatase 2A, an enzyme responsible for the dephosphorylation of Bcl-xL [174,175]. Therefore, omega-3 fatty acids may both decrease the functional population of pro-apoptotic Bcl-2 proteins and increase Bcl-xL in the brain favoring neuroprotection.
DHA treatment is reported to protect the brain in in vivo models of Parkinson’s disease [117]. Mice fed with DHA for 2 weeks were protected from dopaminergic cell loss induced by the dopaminergic toxin, 1-methyl-4-phenyl-1,2,3,6-tetra-hydropyridine (MPTP) [117]. Supplementation of DHA significantly increased the mRNA expression of Bcl-xL, leading to the neuroprotective effect. This group also showed downregulation of phosphorylated JNK in the DHA fed group. Although both the JNK pathway and Bcl-xL are known to play a role in apoptosis, it is unclear if DHA independently controls JNK and Bcl-xL expression, or it intervenes with JNK-induced Bcl-xL regulation. Since phosphorylation is an important modification to change the function of Bcl-xL by blocking post-translational cleavage or interaction with pro-apoptotic proteins [103,176,177,178,179], alteration of upstream kinase activities by DHA may regulate apoptosis.
Moreover, DHA application has been highly effective against oxidative stress-associated damage which is a common process during neurodegeneration [115,119]. Although mechanisms of DHA-mediated epigenetic control were not tested in these reports, DHA-induced augmentation of mRNA expression under oxidative stress is known to occur for the Bcl-xL gene, BCL2L1 in HT22 hippocampal cell line, without changes in Bcl-2 nor Bax gene expression [115]. Thus, DHA-dependent anti-apoptotic effect during oxidative stress may be primarily controlled by the Bcl-xL gene. Finally, treatment with DHA is reported to enhance neurite outgrowth in hippocampal neurons and to prevent loss of neurites during neurotoxicity [120,180]. Since Bcl-xL is required for neurite elongation and branching [21], DHA may be a potent candidate to promote rehabilitation or recovery of the brain after injury.
5. Resveratrol
Resveratrol, or 3,4,5-trihydroxystilbene, is part of a class of polyphenolic compounds called stilbenes. Resveratrol is in foods such as cocoa, various berries, and peanuts, but the most prominent sources in the human diet are grapes and grape products, such as wine. Although both red and white wines contain this compound, resveratrol content is markedly higher in red wines [181]. Resveratrol is notable for its role as a potent antioxidant by scavenging free radicals and increasing the activity of endogenous antioxidant enzymes, such as superoxide dismutase [182,183]. Additionally, it has been found to have additional benefits such as cardioprotection, anti-inflammatory, and anti-tumor properties [184,185,186,187].
Neuroprotective properties of resveratrol have been reported by various research groups. Administration of resveratrol attenuates brain damage caused by MCAO-induced ischemic stroke in rodents [188,189,190]. Resveratrol also exhibits protective effects in neurodegenerative diseases such as Alzheimer’s and Parkinson’s disease [191,192]. Resveratrol treatment regulated Bcl-xL and NF-κB protein levels in an in vitro Alzheimer’s disease model, and protected rat pheochromocytoma cells, PC12, against Aβ-induced oxidative stress [122]. Moreover, involvement of Bcl-xL in resveratrol-mediated neuroprotection in a cerebral ischemia model was reported [121]. Lanzillotta et al. exposed mouse primary cortical neurons to oxygen-glucose deprivation (OGD) for 3 h, and found that OGD decreased the activity of the Bcl-xL promoter. Interestingly, mutation of the nuclear factor-kappaB (NF-κB) binding site at the Bcl-xL promoter downregulated the basal activity of the Bcl-xL gene indicating NF-κB is an important transcriptional modulator of Bcl-xL gene expression [121]. RelA (also called p65), a subunit of NF-κB, is responsible for inducing Bcl-xL gene expression, but not that of Bcl-2 [193]. Lanzillotta’s group previously reported that acetylation of nuclear factor-kappaB (NF-κB) p50/RelA at Lys310 occurs during ischemia stroke, and deacetylation by a pharmacological inhibitor of acetylase and application of Lys310 mutation attenuates MCAO-induced neuronal injury [194]. Resveratrol activates sirtuin 1, an NAD-dependent deacetylase encoded by SIRT1 gene [195,196], thus resveratrol was applied to reverse acetylation of NF-κB RelA. Consistently, resveratrol enhanced Bcl-xL protein and protected SH-SY5Y cells from prion protein-induced cell death [123]. Seo et al. confirmed that resveratrol attenuates acetylation of p65, which enhances Bcl-xL promoter activity [193]. This group also showed that SH-SY5Y cells transduced with SIRT1 siRNA abolished resveratrol-mediated protection indicating that SIRT1 is the primary target of resveratrol in this model. These studies report that manipulating acetylation state using resveratrol supports recruitment of transcription factors at the Bcl-xL promoter enhancing the ratio of anti-apoptotic to pro-apoptotic gene products; this protects the brain against various apoptotic insults.
Although there are limited reports indicating a relationship between Bcl-xL and resveratrol in CNS disease models, numerous studies have reported that Bcl-2 and other members of Bcl-2 proteins are under the control of resveratrol [189,197,198,199,200,201]. Since apoptotic pathways are orchestrated by a series of highly coordinated actions of Bcl-xL and other Bcl-2 family proteins, it may be critical to dissect functions of Bcl-xL during resveratrol-mediated neuroprotection in future research.
6. Alcohol
Alcohol, specifically ethanol, is a substance known to have varying effects on the brain. Although alcohol sensitivities may vary by dosage, time, cell type, and individual genetic differences [202], it is well-reported that aggressive alcohol usage leads to neurological damage, which is causative of behavioral and cognitive changes [203,204]. In contrast to its neurotoxic effects, moderate consumption of alcohol can be neuroprotective, lowering the risk of ischemic stroke and enhancing cognitive function in human subjects [205,206,207,208]. Similarly, treatment with ethanol during or after MCAO procedure improved energy metabolism and protected rodent brain from ischemic injury [209,210].
Despite limited information regarding the underlying mechanisms of alcohol-mediated neuroprotection, Bcl-2 proteins including Bcl-xL have been reported as important molecular targets [124,125,211,212]. Fu et al. showed upregulation of Bcl-xL protein in ethanol treated animals in an ischemic stroke model [124]. Rats undergoing MCAO-induced transient ischemia followed by reperfusion, had significantly decreased pro-apoptotic protein levels such as Bax, caspase 3, and apoptosis inducing factor (AIF) after administration of 1.5 g/kg ethanol, and the ethanol treatment enhanced anti-apoptotic Bcl-xL and Bcl-2 protein levels. Bcl-xL directly binds to Bax to inhibit Bax-mediated membrane permeabilization [213] and interaction between Bcl-xL and Bax prevents AIF release. Sequestration of pro-apoptotic Bax by Bcl-xL also prevents formation of the apoptosome blocking activation of caspase 3. Since caspase 3 is a key contributor in the conversion of anti-apoptotic Bcl-xL to pro-apoptotic fragmented ∆N-Bcl-xL during cerebral stroke [18,65,68], the ability of ethanol to prevent caspase 3 induction and to increase Bcl-xL protein levels directly prevents activation of apoptotic pathways. The same research team has reported that intraperitoneal injection of ethanol (1 g/kg) with normobaric oxygen therapy significantly increased Bcl-xL protein levels and attenuated stroke-induced neurological deficits [125]. However, it is well studied that alcohol consumption can also lead to apoptotic death via decreases in levels of Bcl-2 proteins in the brain [214,215,216]. Ethanol-induced Bcl-xL mRNA expression in brain cells is highly dependent on its concentration [124] and on the length of time of miRNA expression [217], thus determination of the therapeutic window of protection after ethanol treatment will be needed in future projects.
7. Conclusions
In this review, we have discussed food sources, nutrients, and phytochemicals that exert neuroprotection by regulating Bcl-xL (Table 2). Unlike surgical procedures or pharmacological approaches, nutritional therapy may not directly cure CNS disease or immediately relieve symptoms. However, nutritional intervention may delay propagation of neurotoxic signaling in the brain, facilitate recovery of damaged neurons, or support medical treatment by controlling an important molecular target that is essential for mitochondrial energy production, synaptic transmission, neurite outgrowth, and neuronal survival. Here, we suggest that nutrients are participants in molecular events and pathways, and are capable of regulating the protein target Bcl-xL and directly or indirectly other Bcl-2 family members via controlling transcription, translation, post-translational modification, and direct binding. Nutrients with regulatory potential may act synergistically with existing medications to reduce the risk of CNS disease.
Author Contributions
Writing-review & editing: H.-A.P.; K.B.; A.S.; S.K.; E.A.J.
Funding
This research was funded by Summer Research Support by the College of Human Environmental Sciences, the University of Alabama.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Barclay, L.A.; Wales, T.E.; Garner, T.P.; Wachter, F.; Lee, S.; Guerra, R.M.; Stewart, M.L.; Braun, C.R.; Bird, G.H.; Gavathiotis, E.; et al. Inhibition of pro-apoptotic bax by a noncanonical interaction mechanism. Mol. Cell 2015, 57, 873–886. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.C.S. The conserved N-terminal bh4 domain of Bcl-2 homologues is essential for inhibition of apoptosis and interaction with ced-4. EMBO J. 1998, 17, 1029–1039. [Google Scholar] [CrossRef] [PubMed]
- Monaco, G.; La Rovere, R.; Karamanou, S.; Welkenhuyzen, K.; Ivanova, H.; Vandermarliere, E.; Di Martile, M.; Del Bufalo, D.; De Smedt, H.; Parys, J.B.; et al. A double point mutation at residues Ile14 and Val15 of Bcl-2 uncovers a role for the BH4 domain in both protein stability and function. FEBS J. 2018, 285, 127–145. [Google Scholar] [CrossRef] [PubMed]
- Rong, Y.-P.; Bultynck, G.; Aromolaran, A.S.; Zhong, F.; Parys, J.B.; De Smedt, H.; Mignery, G.A.; Roderick, H.L.; Bootman, M.D.; Distelhorst, C.W. The bh4 domain of Bcl-2 inhibits er calcium release and apoptosis by binding the regulatory and coupling domain of the ip3 receptor. Proc. Nat. Acad. Sci. USA 2009, 106, 14397–14402. [Google Scholar] [CrossRef] [PubMed]
- Westphal, D.; Kluck, R.M.; Dewson, G. Building blocks of the apoptotic pore: How bax and bak are activated and oligomerize during apoptosis. Cell Death Differ. 2013, 21, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Desagher, S.; Osen-Sand, A.; Nichols, A.; Eskes, R.; Montessuit, S.; Lauper, S.; Maundrell, K.; Antonsson, B.; Martinou, J.-C. Bid-induced conformational change of bax is responsible for mitochondrial cytochrome c release during apoptosis. J. Cell Biol. 1999, 144, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.C.; Lindsten, T.; Mootha, V.K.; Weiler, S.; Gross, A.; Ashiya, M.; Thompson, C.B.; Korsmeyer, S.J. tBID, a membrane-targeted death ligand, oligomerizes BAK to release cytochrome c. Gene. Dev. 2000, 14, 2060–2071. [Google Scholar] [PubMed]
- Wei, M.C. Proapoptotic bax and bak: A requisite gateway to mitochondrial dysfunction and death. Science 2001, 292, 727–730. [Google Scholar] [CrossRef] [PubMed]
- Certo, M.; Moore, V.D.G.; Nishino, M.; Wei, G.; Korsmeyer, S.; Armstrong, S.A.; Letai, A. Mitochondria primed by death signals determine cellular addiction to antiapoptotic Bcl-2 family members. Cancer Cell 2006, 9, 351–365. [Google Scholar] [CrossRef] [PubMed]
- Sattler, M. Structure of Bcl-xl-bak peptide complex: Recognition between regulators of apoptosis. Science 1997, 275, 983–986. [Google Scholar] [CrossRef] [PubMed]
- Jurgensmeier, J.M.; Xie, Z.; Deveraux, Q.; Ellerby, L.; Bredesen, D.; Reed, J.C. Bax directly induces release of cytochrome c from isolated mitochondria. Proc. Natl. Acad. Sci. USA 1998, 95, 4997–5002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petros, A.M.; Nettesheim, D.G.; Wang, Y.; Olejniczak, E.T.; Meadows, R.P.; Mack, J.; Swift, K.; Matayoshi, E.D.; Zhang, H.; Fesik, S.W.; et al. Rationale for Bcl-xl/bad peptide complex formation from structure, mutagenesis, and biophysical studies. Protein Sci. 2000, 9, 2528–2534. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.; Zha, J.; Jockel, J.; Boise, L.H.; Thompson, C.B.; Korsmeyer, S.J. Bad, a heterodimeric partner for Bcl-xl and Bcl-2, displaces bax and promotes cell death. Cell 1995, 80, 285–291. [Google Scholar] [CrossRef]
- Chen, Y.B.; Aon, M.A.; Hsu, Y.T.; Soane, L.; Teng, X.; McCaffery, J.M.; Cheng, W.-C.; Qi, B.; Li, H.; Alavian, K.N.; et al. Bcl-xL regulates mitochondrial energetics by stabilizing the inner membrane potential. J. Cell Biol. 2011, 195, 263–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertini, I.; Chevance, S.; Del Conte, R.; Lalli, D.; Turano, P. The anti-apoptotic Bcl-x(L.) protein, a new piece in the puzzle of cytochrome c interactome. PLoS ONE 2011, 6, e18329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kharbanda, S.; Pandey, P.; Schofield, L.; Israels, S.; Roncinske, R.; Yoshida, K.; Bharti, A.; Yuan, Z.-M.; Saxena, S.; Weichselbaum, R.; et al. Role for Bcl-xl as an inhibitor of cytosolic cytochrome c accumulation in dna damage-induced apoptosis. Proc. Natl. Acad. Sci. USA 1997, 94, 6939–6942. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Swahari, V.; Plestant, C.; Smith, I.; McCoy, E.; Smith, S.; Moy, S.S.; Anton, E.S.; Deshmukh, M. Bcl-xl is essential for the survival and function of differentiated neurons in the cortex that control complex behaviors. J. Neurosci. 2016, 36, 5448–5461. [Google Scholar] [CrossRef] [PubMed]
- Ofengeim, D.; Chen, Y.; Miyawaki, T.; Li, H.; Sacchetti, S.; Flannery, R.J.; Alavian, K.N.; Pontarelli, F.; Roelofs, B.A.; Hickman, J.A.; et al. N-terminally cleaved Bcl-xl mediates ischemia-induced neuronal death. Nat. Neurosci. 2012, 15, 574–580. [Google Scholar] [CrossRef] [PubMed]
- Naumenko, V.S.; Kulikov, A.V.; Kondaurova, E.M.; Tsybko, A.S.; Kulikova, E.A.; Krasnov, I.B.; Shenkman, B.S.; Sychev, V.N.; Bazhenova, E.Y.; Sinyakova, N.A.; et al. Effect of actual long-term spaceflight on bdnf, trkb, p75, bax and Bcl-xl genes expression in mouse brain regions. Neuroscience 2015, 284, 730–736. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, Y.; Liu, J.Q.; Nakano, Y.; Ueno, S.; Ohmori, S.; Fueta, Y.; Ishidao, T.; Kunugita, N.; Yamashita, U.; Hori, H. 1-bp inhibits nf-κb activity and Bcl-xl expression in astrocytes in vitro and reduces Bcl-xl expression in the brains of rats in vivo. NeuroToxicology 2007, 28, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-A.; Licznerski, P.; Alavian, K.N.; Shanabrough, M.; Jonas, E.A. Bcl-xl is necessary for neurite outgrowth in hippocampal neurons. Antioxid. Redox 2015, 22, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kannagi, M.; Ferrante, R.J.; Kowall, N.W.; Ryu, H. Activation of ets-2 by oxidative stress induces Bcl-xl expression and accounts for glial survival in amyotrophic lateral sclerosis. FASEB J. 2009, 23, 1739–1749. [Google Scholar] [CrossRef] [PubMed]
- Krajewska, M.; Mai, J.K.; Zapata, J.M.; Ashwell, K.W.; Schendel, S.L.; Reed, J.C.; Krajewski, S. Dynamics of expression of apoptosis-regulatory proteins bid, Bcl-2, Bcl-x, bax and bak during development of murine nervous system. Cell Death Differ. 2002, 9, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Alavian, K.N.; Li, H.; Collis, L.; Bonanni, L.; Zeng, L.; Sacchetti, S.; Lazrove, E.; Nabili, P.; Flaherty, B.; Graham, M.; et al. Bcl-xl regulates metabolic efficiency of neurons through interaction with the mitochondrial f1fo atp synthase. Nat. Cell Biol. 2011, 13, 1224–1233. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Alavian, K.N.; Lazrove, E.; Mehta, N.; Jones, A.; Zhang, P.; Licznerski, P.; Graham, M.; Uo, T.; Guo, J.; et al. A Bcl-xl–drp1 complex regulates synaptic vesicle membrane dynamics during endocytosis. Nat. Cell Biol. 2013, 15, 773–785. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chen, Y.; Jones, A.F.; Sanger, R.H.; Collis, L.P.; Flannery, R.; McNay, E.C.; Yu, T.; Schwarzenbacher, R.; Bossy, B.; et al. Bcl-xl induces drp1-dependent synapse formation in cultured hippocampal neurons. Proc. Natl. Acad. Sci. USA 2008, 105, 2169–2174. [Google Scholar] [CrossRef] [PubMed]
- Jonas, E. BCL-xL regulates synaptic plasticity. Mol. Interv. 2006, 6, 208–222. [Google Scholar] [CrossRef] [PubMed]
- Jonas, E.; Porter, G.A.; Beutner, G.; Mnatsakanyan, N.; Park, H.A.; Mehta, N.; Chen, R.; Alavian, K.N. The mitochondrial permeability transition pore: Molecular structure and function in health and disease. In Molecular Basis for Mitochondrial Signaling; Rostovtseva, T.K., Ed.; Springer: New York, NY, USA, 2017; pp. 69–105. [Google Scholar]
- Park, H.A.; Jonas, E. Mitochondrial regulators of synaptic plasticity in the ischemic brain. In Synaptic Plasticity; Heinbockel, T., Ed.; INTECH: London, UK, 2017; pp. 39–67. [Google Scholar]
- Cao, G.; Pei, W.; Ge, H.; Liang, Q.; Luo, Y.; Sharp, F.R.; Lu, A.; Ran, R. In vivo delivery of a Bcl-xl fusion protein containing the tat protein transduction domain protects against ischemic brain injury and neuronal apoptosis. J. Neurosci. 2002, 22, 5423–5431. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Fujihara, H.; Yao, J.; Qi, S.; Li, H.; Shimoji, K. Different expression patterns of Bcl-2, Bcl-xl, and Bax proteins after sublethal forebrain ischemia in C57Black/Crj6 mouse striatum. Stroke A J. Cerebr. Circ. 2003, 34, 1803–1808. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, S.; Eguchi, Y.; Kosaka, H.; Kamiike, W.; Matsuda, H.; Tsujimoto, Y. Prevention of hypoxia-induced cell death by Bcl-2 and Bcl-xl. Nature 1995, 374, 811–813. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Graham, S.H.; Nakayama, M.; Zhu, R.L.; Jin, K.; Stetler, R.A.; Simon, R.P. Apoptosis repressor genes Bcl-2 and Bcl-x-long are expressed in the rat brain following global ischemia. J. Cereb. Blood Flow Metab. 1997, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Wiessner, C.; Allegrini, P.R.; Rupalla, K.; Sauer, D.; Oltersdorf, T.; McGregor, A.L.; Bischoff, S.; Böttiger, B.W.; van der Putten, H. Neuron-specific transgene expression of Bcl-xl but not Bcl-2 genes reduced lesion size after permanent middle cerebral artery occlusion in mice. Neurosci. Lett. 1999, 268, 119–122. [Google Scholar] [CrossRef]
- Kilic, E.; Dietz, G.P.; Hermann, D.M.; Bahr, M. Intravenous TAT-Bcl-Xl is protective after middle cerebral artery occlusion in mice. Ann Neurol. 2002, 52, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Nesic, O.; Ye, Z.; Rea, H.; Westlund, K.N.; Xu, G.Y.; McAdoo, D.; Hulsebosch, C.E.; Perez-Polo, J.R. Bcl-xL expression after contusion to the rat spinal cord. J. Neurotrauma 2001, 18, 1267–1278. [Google Scholar] [CrossRef] [PubMed]
- Nesic-Taylor, O.; Cittelly, D.; Ye, Z.; Xu, G.Y.; Unabia, G.; Lee, J.C.; Svrakic, N.M.; Liu, X.H.; Youle, R.J.; Wood, T.G.; et al. Exogenous Bcl-xl fusion protein spares neurons after spinal cord injury. J. Neurosci. Res. 2005, 79, 628–637. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.I.; Kim, B.G.; Hwang, D.H.; Kim, H.M.; Kim, S.U. Overexpression of Bcl-xlin human neural stem cells promotes graft survival and functional recovery following transplantation in spinal cord injury. J. Neurosci. Res. 2009, 87, 3186–3197. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Peng, X.; Insolera, R.; Fink, D.J.; Mata, M. IL-10 promotes neuronal survival following spinal cord injury. Exp Neurol. 2009, 220, 183–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadota, R.; Koda, M.; Kawabe, J.; Hashimoto, M.; Nishio, Y.; Mannoji, C.; Miyashita, T.; Furuya, T.; Okawa, A.; Takahashi, K.; et al. Granulocyte colony-stimulating factor (g-csf) protects oligpdendrocyte and promotes hindlimb functional recovery after spinal cord injury in rats. PLoS ONE 2012, 7, e50391. [Google Scholar] [CrossRef] [PubMed]
- Pike, C.J. Estrogen modulates neuronal Bcl-xL expression and beta-amyloid-induced apoptosis: Relevance to Alzheimer’s disease. J. Neurochem. 1999, 72, 1552–1563. [Google Scholar] [CrossRef] [PubMed]
- Keil, U.; Bonert, A.; Marques, C.A.; Scherping, I.; Weyermann, J.; Strosznajder, J.B.; Müller-Spahn, F.; Haass, C.; Czech, C.; Pradier, L.; et al. Amyloid β-induced changes in nitric oxide production and mitochondrial activity lead to apoptosis. J. Biol. Chem. 2004, 279, 50310–50320. [Google Scholar] [CrossRef] [PubMed]
- Hauptmann, S.; Scherping, I.; Dröse, S.; Brandt, U.; Schulz, K.L.; Jendrach, M.; Leuner, K.; Eckert, A.; Müller, W.E. Mitochondrial dysfunction: An early event in alzheimer pathology accumulates with age in ad transgenic mice. Neurobiol. Aging 2009, 30, 1574–1586. [Google Scholar] [CrossRef] [PubMed]
- Vohra, B.P.S.; Sasaki, Y.; Miller, B.R.; Chang, J.; DiAntonio, A.; Milbrandt, J. Amyloid precursor protein cleavage-dependent and -independent axonal degeneration programs share a common nicotinamide mononucleotide adenylyltransferase 1-sensitive pathway. J. Neurosci. 2010, 30, 13729–13738. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, A.; Mouatt-Prigent, A.; Vila, M.; Abbas, N.; Perier, C.; Faucheux, B.A.; Vyas, S.; Hirsch, E.C. Increased expression and redistribution of the antiapoptotic molecule Bcl-xL in Parkinson’s disease. Neurobiol. Dis. 2002, 10, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Liste, I. The generation of dopaminergic neurons by human neural stem cells is enhanced by Bcl-xl, both in vitro and in vivo. J. Neurosci. 2004, 24, 10786–10795. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.-W. Enhanced in vitro midbrain dopamine neuron differentiation, dopaminergic function, neurite outgrowth, and 1-methyl-4-phenylpyridium resistance in mouse embryonic stem cells overexpressing Bcl-xl. J. Neurosci. 2004, 24, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Krabbe, C.; Courtois, E.; Jensen, P.; Jørgensen, J.R.; Zimmer, J.; Martínez-Serrano, A.; Meyer, M. Enhanced dopaminergic differentiation of human neural stem cells by synergistic effect of Bcl-xland reduced oxygen tension. J. Neurochem. 2009, 110, 1908–1920. [Google Scholar] [CrossRef] [PubMed]
- Courtois, E.T.; Castillo, C.G.; Seiz, E.G.; Ramos, M.; Bueno, C.; Liste, I.; Martinez-Serrano, A. In vitro and in vivo enhanced generation of human a9 dopamine neurons from neural stem cells by Bcl-xl. J. Biol. Chem. 2010, 285, 9881–9897. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Fu, K.; Wang, D.; Mu, C.; Wang, G. Oxidized dj-1 interacts with the mitochondrial protein Bcl-xl. J. Biol. Chem. 2011, 286, 35308–35317. [Google Scholar] [CrossRef] [PubMed]
- Chiou, S.H.; Ku, H.H.; Tsai, T.H.; Lin, H.L.; Chen, L.H.; Chien, C.S.; Ho, L.L.-T.; Lee, C.-H.; Chang, Y.-L. Moclobemide upregulated Bcl-2 expression and induced neural stem cell differentiation into serotoninergic neuron via extracellular-regulated kinase pathway. Br. J. Pharmacol. 2006, 148, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Kosten, T.A.; Galloway, M.P.; Duman, R.S.; Russell, D.S.; D’Sa, C. Repeated unpredictable stress and antidepressants differentially regulate expression of the Bcl-2 family of apoptotic genes in rat cortical, hippocampal, and limbic brain structures. Neuropsychopharmacology 2007, 33, 1545–1558. [Google Scholar] [CrossRef] [PubMed]
- Shishkina, G.T.; Kalinina, T.S.; Berezova, I.V.; Dygalo, N.N. Stress-induced activation of the brainstem Bcl-xl gene expression in rats treated with fluoxetine: Correlations with serotonin metabolism and depressive-like behavior. Neuropharmacology 2012, 62, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Li, G.; Zhang, Y.; Su, X.; Hang, C. The potential role of jak2/stat3 pathway on the anti-apoptotic effect of recombinant human erythropoietin (rhepo) after experimental traumatic brain injury of rats. Cytokine 2011, 56, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Lončarević-Vasiljković, N.; Milanović, D.; Pešić, V.; Tešić, V.; Brkić, M.; Lazić, D.; Avramović, V.; Kanazir, S. Dietary restriction suppresses apoptotic cell death, promotes Bcl-2 and Bcl-xl mrna expression and increases the Bcl-2/bax protein ratio in the rat cortex after cortical injury. Neurochem. Int. 2016, 96, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Pang, A.L.; Xiong, L.L.; Xia, Q.J.; Liu, F.; Wang, Y.C.; Liu, F.; Zhang, P.; Meng, B.-L.; Tan, S.; Wang, T.-H. Neural stem cell transplantation is associated with inhibition of apoptosis, Bcl-xL upregulation, and recovery of neurological function in a rat model of traumatic. brain injury. Cell Transpl. 2017, 26, 1262–1275. [Google Scholar] [CrossRef] [PubMed]
- Soler-Botija, C.; Ferrer, I.; Alvarez, J.L.; Baiget, M.; Tizzano, E.F. Downregulation of Bcl-2 proteins in type i spinal muscular atrophy motor neurons during fetal development. J. Neuropathol. Exp. Neurol. 2003, 62, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Tsai, L.K.; Tsai, M.S.; Ting, C.H.; Li, H. Multiple therapeutic effects of valproic acid in spinal muscular atrophy model mice. J. Mol. Med. 2008, 86, 1243–1254. [Google Scholar] [CrossRef] [PubMed]
- Tsai, L.K.; Tsai, M.S.; Ting, C.H.; Wang, S.H.; Li, H. Restoring Bcl-x(L) levels benefits a mouse model of spinal muscular atrophy. Neurobiol. Dis. 2008, 31, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Garcera, A.; Mincheva, S.; Gou-Fabregas, M.; Caraballo-Miralles, V.; Lladó, J.; Comella, J.X.; Soler, R.M. A new model to study spinal muscular atrophy: Neurite degeneration and cell death is counteracted by Bcl-xl overexpression in motoneurons. Neurobiol. Dis. 2011, 42, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Anderton, R.S.; Price, L.L.; Turner, B.J.; Meloni, B.P.; Mitrpant, C.; Mastaglia, F.L.; Goh, C.; Wilton, S.D.; Boulos, S. Co-regulation of survival of motor neuron and Bcl-xl expression: Implications for neuroprotection in spinal muscular atrophy. Neuroscience 2012, 220, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Kaufmann, J.A.; Sanchez-Ross, M.G.; Johnson, K.M. Mechanisms of N-methyl-d-aspartate-induced apoptosis in phencyclidine-treated cultured forebrain neurons. J. Pharmacol. Exp. Ther. 2000, 294, 287–295. [Google Scholar] [PubMed]
- Wang, C.; McInnis, J.; Ross-Sanchez, M.; Shinnick-Gallagher, P.; Wiley, J.; Johnson, K. Long-term behavioral and neurodegenerative effects of perinatal phencyclidine administration: Implications for schizophrenia. Neuroscience 2001, 107, 535–550. [Google Scholar] [CrossRef]
- He, J.; Xu, H.; Yang, Y.; Rajakumar, D.; Li, X.; Li, X.-M. The effects of chronic administration of quetiapine on the phencyclidine-induced reference memory impairment and decrease of Bcl-xl/Bax ratio in the posterior cingulate cortex in rats. Behav. Brain Res. 2006, 168, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Clem, R.J.; Cheng, E.H.-Y.; Karp, C.L.; Kirsch, D.G.; Ueno, K.; Takahashi, A.; Kastan, M.B.; Griffin, D.E.; Earnshaw, W.C.; Veliuona, M.A.; et al. Modulation of cell death by Bcl-xl through caspase interaction. Proc. Natl. Acad. Sci. USA 1998, 95, 554–559. [Google Scholar] [CrossRef] [PubMed]
- Miyawaki, T.; Mashiko, T.; Ofengeim, D.; Flannery, R.J.; Noh, K.-M.; Fujisawa, S.; Bonanni, L.; Bennett, M.V.L.; Zukin, R.S.; Jonas, E.A. Ischemic preconditioning blocks bad translocation, Bcl-xl cleavage, and large channel activity in mitochondria of postischemic hippocampal neurons. Proc. Natl. Acad. Sci. USA 2008, 105, 4892–4897. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Yokota, H.; Jover, T.; Cappuccio, I.; Calderone, A.; Simionescu, M.; Bennett, M.V.; Zukin, R.S. Ischemic preconditioning: Neuronal survival in the face of caspase-3 activation. J. Neurosci. 2004, 24, 2750–2759. [Google Scholar] [CrossRef] [PubMed]
- Tornero, D.; Posadas, I.; Ceña, V. Bcl-xl blocks a mitochondrial inner membrane channel and prevents ca2+ overload-mediated cell death. PLoS ONE 2011, 6, e20423. [Google Scholar] [CrossRef] [PubMed]
- Park, H.A.; Jonas, E.A. ΔN-Bcl-xL, a therapeutic target for neuroprotection. Neural Regen Res. 2017, 12, 1791–1794. [Google Scholar] [CrossRef] [PubMed]
- Jonas, E.A.; Hickman, J.A.; Chachar, M.; Polster, B.M.; Brandt, T.A.; Fannjiang, Y.; Ivanovska, I.; Basanez, G.; Kinnally, K.W.; Zimmerberg, J.; et al. Proapoptotic n-truncated Bcl-xl protein activates endogenous mitochondrial channels in living synaptic terminals. Proc. Natl. Acad. Sci. USA 2004, 101, 13590–13595. [Google Scholar] [CrossRef] [PubMed]
- Alavian, K.N.; Beutner, G.; Lazrove, E.; Sacchetti, S.; Park, H.-A.; Licznerski, P.; Li, H.; Nabili, P.; Hockensmith, K.; Graham, M.; et al. An uncoupling channel within the c-subunit ring of the f1fo atp synthase is the mitochondrial permeability transition pore. Proc. Natl. Acad. Sci. USA 2014, 111, 10580–10585. [Google Scholar] [CrossRef] [PubMed]
- White, C.; Li, C.; Yang, J.; Petrenko, N.B.; Madesh, M.; Thompson, C.B.; Foskett, J.K. The endoplasmic reticulum gateway to apoptosis by Bcl-xl modulation of the insp3r. Nat. Cell Biol. 2005, 7, 1021–1028. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, X.; Vais, H.; Thompson, C.B.; Foskett, J.K.; White, C. Apoptosis regulation by Bcl-xl modulation of mammalian inositol 1,4,5-trisphosphate receptor channel isoform gating. Proc. Natl. Acad. Sci. USA 2007, 104, 12565–12570. [Google Scholar] [CrossRef] [PubMed]
- Denton, R.M. Regulation of mitochondrial dehydrogenases by calcium ions. Biochim. Biophys. Acta 2009, 1787, 1309–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, J.C.K.; DiLorenzo, J.C.; Sheu, K.-F.R. Pyruvate dehydrogenase complex is inhibited in calcium-loaded cerebrocortical mitochondria. Neurochem. Res. 1988, 13, 1043–1048. [Google Scholar] [CrossRef] [PubMed]
- Contreras, L.; Satrustegui, J. Calcium signaling in brain mitochondria: Interplay of malate aspartate nadh shuttle and calcium uniporter/mitochondrial dehydrogenase pathways. J. Biol. Chem. 2009, 284, 7091–7099. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, M.J.; McHugh, N.J. Mitochondrial ATP synthase F1-beta-subunit is a calcium-binding protein. FEBS Lett. 1996, 391, 323–329. [Google Scholar] [CrossRef]
- Territo, P.R.; Mootha, V.K.; French, S.A.; Balaban, R.S. Ca2+ activation of heart mitochondrial oxidative phosphorylation: Role of the F(0)/F(1)-ATPase. Am. J. Physiol. Cell Physiol. 2000, 278, C423–C435. [Google Scholar] [CrossRef] [PubMed]
- Boerries, M.; Most, P.; Gledhill, J.R.; Walker, J.E.; Katus, H.A.; Koch, W.J.; Aebi, U.; Schoenenberger, C.-A. Ca2+-dependent interaction of s100a1 with f1-atpase leads to an increased atp content in cardiomyocytes. Mol. Cell. Biol. 2007, 27, 4365–4373. [Google Scholar] [CrossRef] [PubMed]
- Giorgio, V.; Burchell, V.; Schiavone, M.; Bassot, C.; Minervini, G.; Petronilli, V.; Argenton, F.; Forte, M.; Tosatto, S.; Lippe, G.; et al. Ca2+ binding to F-ATP synthase β subunit triggers the mitochondrial permeability transition. EMBO Rep. 2017, 18, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Bezprozvanny, I.; Ehrlich, B.E. Atp modulates the function of inositol 1,4,5-trisphosphate-gated channels at two sites. Neuron 1993, 10, 1175–1184. [Google Scholar] [CrossRef]
- Wagner, L.E.; Betzenhauser, M.J.; Yule, D.I. Atp binding to a unique site in the type-1 s2- inositol 1,4,5-trisphosphate receptor defines susceptibility to phosphorylation by protein kinase a. J. Biol. Chem. 2006, 281, 17410–17419. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Hu, X.; Eno, C.O.; Zhao, G.; Li, C.; White, C. An interaction between Bcl-xland the voltage-dependent anion channel (vdac) promotes mitochondrial Ca2+ uptake. J. Biol. Chem. 2013, 288, 19870–19881. [Google Scholar] [CrossRef] [PubMed]
- Shoshan-Barmatz, V.; Krelin, Y.; Shteinfer-Kuzmine, A. VDAC1 functions in Ca2+ homeostasis and cell life and death in health and disease. Cell Calcium 2018, 69, 81–100. [Google Scholar] [CrossRef] [PubMed]
- Monaco, G.; Decrock, E.; Arbel, N.; van Vliet, A.R.; La Rovere, R.M.; De Smedt, H.; Parys, J.B.; Agostinis, P.; Leybaert, L.; Shoshan-Barmatz, V.; et al. The bh4 domain of anti-apoptotic Bcl-xl, but not that of the related Bcl-2, limits the voltage-dependent anion channel 1 (vdac1)-mediated transfer of pro-apoptotic ca2+signals to mitochondria. J. Biol. Chem. 2015, 290, 9150–9161. [Google Scholar] [CrossRef] [PubMed]
- Madesh, M.; Hajnoczky, G. VDAC-dependent permeabilization of the outer mitochondrial membrane by superoxide induces rapid and massive cytochrome c release. J. Cell Biol. 2001, 155, 1003–1015. [Google Scholar] [CrossRef] [PubMed]
- Lemasters, J.J.; Holmuhamedov, E. Voltage-dependent anion channel (vdac) as mitochondrial governator—Thinking outside the box. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2006, 1762, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Arbel, N.; Ben-Hail, D.; Shoshan-Barmatz, V. Mediation of the antiapoptotic activity of Bcl-xl protein upon interaction with vdac1 protein. J. Biol. Chem. 2012, 287, 23152–23161. [Google Scholar] [CrossRef] [PubMed]
- Malia, T.J.; Wagner, G. Nmr structural investigation of the mitochondrial outer membrane protein vdac and its interaction with antiapoptotic Bcl-xl†. Biochemistry 2007, 46, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Vander Heiden, M.G. Bcl-xl promotes the open configuration of the voltage-dependent anion channel and metabolite passage through the outer mitochondrial membrane. J. Biol. Chem. 2001, 276, 19414–19419. [Google Scholar] [CrossRef] [PubMed]
- Vander-Heiden, M.G.; Chandel, N.S.; Li, X.X.; Schumacker, P.T.; Colombini, M.; Thompson, C.B. Outer mitochondrial membrane permeability can regulate coupled respiration and cell survival. Proc. Nat. Acad. Sci. USA 2000, 97, 4666–4671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishida, Y.; Arakawa, S.; Fujitani, K.; Yamaguchi, H.; Mizuta, T.; Kanaseki, T.; Komatsu, M.; Otsu, K.; Tsujimoto, Y.; Shimizu, S. Discovery of atg5/atg7-independent alternative macroautophagy. Nature 2009, 461, 654–658. [Google Scholar] [CrossRef] [PubMed]
- Kang, R.; Zeh, H.J.; Lotze, M.T.; Tang, D. The Beclin 1 network regulates autophagy and apoptosis. Cell Death Diff. 2011, 18, 571–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, S.; Levine, B. The autophagy effector Beclin 1: A novel BH3-only protein. Oncogene 2008, 27, S137–S148. [Google Scholar] [CrossRef] [PubMed]
- Oberstein, A.; Jeffrey, P.D.; Shi, Y. Crystal structure of the Bcl-xl-beclin 1 peptide complex: Beclin 1 is a novel bh3-only protein. J. Biol. Chem. 2007, 282, 13123–13132. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-Y.; Song, X.; Zhang, L.; Bartlett, D.L.; Lee, Y.J. Role of Bcl-xl/beclin-1 in interplay between apoptosis and autophagy in oxaliplatin and bortezomib-induced cell death. Biochem. Pharmacol. 2014, 88, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Maiuri, M.C.; Le Toumelin, G.; Criollo, A.; Rain, J.-C.; Gautier, F.; Juin, P.; Tasdemir, E.; Pierron, G.; Troulinaki, K.; Tavernarakis, N.; et al. Functional and physical interaction between Bcl-xl and a bh3-like domain in beclin-1. EMBO J. 2007, 26, 2527–2539. [Google Scholar] [CrossRef] [PubMed]
- Lazarou, M.; Sliter, D.A.; Kane, L.A.; Sarraf, S.A.; Wang, C.; Burman, J.L.; Sideris, D.P.; Fogel, A.I.; Youle, R.J. The ubiquitin kinase pink1 recruits autophagy receptors to induce mitophagy. Nature 2015, 524, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Ordureau, A.; Sarraf, S.A.; Duda, D.M.; Heo, J.-M.; Jedrychowski, M.P.; Sviderskiy, V.O.; Olszewski, J.L.; Koerber, J.T.; Xie, T.; Beausoleil, S.A.; et al. Quantitative proteomics reveal a feedforward mechanism for mitochondrial parkin translocation and ubiquitin chain synthesis. Mol. Cell 2014, 56, 360–375. [Google Scholar] [CrossRef] [PubMed]
- McWilliams, T.G.; Muqit, M.M. PINK1 and parkin: Emerging themes in mitochondrial homeostasis. Curr. Opin. Cell Biol. 2017, 45, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Pickrell, A.M.; Youle, R.J. The roles of PINK1, parkin, and mitochondrial fidelity in Parkinson’s disease. Neuron 2015, 85, 257–273. [Google Scholar] [CrossRef] [PubMed]
- Geisler, S.; Holmström, K.M.; Treis, A.; Skujat, D.; Weber, S.S.; Fiesel, F.C.; Kahle, P.J.; Springer, W. The pink1/parkin-mediated mitophagy is compromised by pd-associated mutations. Autophagy 2010, 6, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Arena, G.; Gelmetti, V.; Torosantucci, L.; Vignone, D.; Lamorte, G.; De Rosa, P.; Cilia, E.; Jonas, E.A.; Valente, E.M. Pink1 protects against cell death induced by mitochondrial depolarization, by phosphorylating Bcl-xl and impairing its pro-apoptotic cleavage. Cell Death Differ. 2013, 20, 920–930. [Google Scholar] [CrossRef] [PubMed]
- Hickman, J.A.; Hardwick, J.M.; Kaczmarek, L.K.; Jonas, E.A. Bcl-xl inhibitor abt-737 reveals a dual role for Bcl-xl in synaptic transmission. J. Neurophysiol. 2008, 99, 1515–1522. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Perkins, G.A.; Poblenz, A.T.; Harris, J.B.; Hung, M.; Ellisman, M.H.; Fox, D.A. Bcl-xl overexpression blocks bax-mediated mitochondrial contact site formation and apoptosis in rod photoreceptors of lead-exposed mice. Proc. Nat. Acad. Sci. USA 2003, 100, 1022–1027. [Google Scholar] [CrossRef] [PubMed]
- Lovekamp-Swan, T.; Glendenning, M.L.; Schreihofer, D.A. A high soy diet enhances neurotropin receptor and Bcl-xl gene expression in the brains of ovariectomized female rats. Brain Res. 2007, 1159, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Lovekamp-Swan, T.; Glendenning, M.; Schreihofer, D.A. A high soy diet reduces programmed cell death and enhances Bcl-xl expression in experimental stroke. Neuroscience 2007, 148, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.-F.; He, L.; Li, D.; Yuan, L.-H.; Yu, H.-L.; Ma, W.-W.; Yang, Y.; Xi, Y.-D.; Ding, J.; Xiao, Y.-X.; et al. Antagonizing effects of soybean isoflavones on β-amyloid peptides- induced oxidative damage in neuron mitochondria of rats. Basic Clin. Pharmacol. Toxicol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Hata, R.; Zhu, P.; Sato, K.; Wen, T.-C.; Yang, L.; Fujita, H.; Mitsuda, N.; Tanaka, J.; Samukawa, K.; et al. Prevention of ischemic neuronal death by intravenous infusion of a ginseng saponin, ginsenoside rb1, that upregulates Bcl-xl expression. J. Cereb. Blood Flow Metab. 2006, 26, 708–721. [Google Scholar] [CrossRef] [PubMed]
- Sakanaka, M.; Zhu, P.; Zhang, B.; Wen, T.-C.; Cao, F.; Ma, Y.-J.; Samukawa, K.; Mitsuda, N.; Tanaka, J.; Kuramoto, M.; et al. Intravenous infusion of dihydroginsenoside rb1 prevents compressive spinal cord injury and ischemic brain damage through upregulation of vegf and Bcl-xL. J. Neurotrauma 2007, 24, 1037–1054. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Samukawa, K.; Fujita, H.; Kato, H.; Sakanaka, M. Oral administration of red ginseng extract promotes neurorestoration after compressive spinal cord injury in rats. Evid-Based Compl. Alt. Med. 2017, 2017, 1265464. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, S.O.; Kim, G.L.; Rhee, D.K. Estrogen receptor-beta of microglia underlies sexual differentiation of neuronal protection via ginsenosides in mice brain. CNS Neurosci. Ther. 2018, 24, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Hata, R.; Nakata, K.; Cao, F.; Samukawa, K.; Fujita, H.; Sakanaka, M. Intravenous infusion of ginsenoside Rb1 ameliorates compressive spinal cord injury through upregulation of Bcl-xL and VEGF. Int. J. Neurol. Neurother. 2015, 2, 1–6. [Google Scholar] [CrossRef]
- Lukiw, W.J. A role for docosahexaenoic acid-derived neuroprotectin d1 in neural cell survival and alzheimer disease. J. Clin. Investig. 2005, 115, 2774–2783. [Google Scholar] [CrossRef] [PubMed]
- Cieslik, M.; Pyszko, J.; Strosznajder, J.B. Docosahexaenoic acid and tetracyclines as promising neuroprotective compounds with poly(adp-ribose) polymerase inhibitory activities for oxidative/genotoxic stress treatment. Neurochem. Int. 2013, 62, 626–636. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.A.; Khare, P.; Rai, A.; Maurya, S.K.; Pathak, A.; Mohan, V.; Nagar, G.K.; Mudiam, M.K.R.; Godbole, M.M.; Bandyopadhyay, S. Anti-apoptotic role of omega-3-fatty acids in developing brain: Perinatal hypothyroid rat cerebellum as apoptotic model. Int. J. Dev. Neurosci. 2009, 27, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, D.; Xu, J.; Yanagita, T.; Xue, C.; Zhang, T.; Wang, W. DHA enriched phospholipids with different polar groups (PC and PS) had different improvements on MPTP-induced mice with Parkinson’s disease. J. Funct. Foods. 2018, 45, 414–426. [Google Scholar] [CrossRef]
- Shi, Z.; Ren, H.; Luo, C.; Yao, X.; Li, P.; He, C.; Kang, J.-X.; Wan, J.-B.; Yuan, T.-F.; Su, H. Enriched endogenous omega-3 polyunsaturated fatty acids protect cortical neurons from experimental ischemic injury. Mol. Neurobiol. 2015, 53, 6482–6488. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wu, P.; Zhang, J.H.; Li, Y.; Xu, S.; Wang, C.; Wang, L.; Zhang, G.; Dai, J.; Zhu, S.; et al. Docosahexaenoic acid alleviates oxidative stress-based apoptosis via improving mitochondrial dynamics in early brain injury after subarachnoid hemorrhage. Cell Mol. Neurobiol. 2018, 38, 1413–1423. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Ren, H.; Shi, Z.; Yao, X.; He, C.; Kang, J.-X.; Wan, J.-B.; Li, P.; Yuan, T.-F.; Su, H. Endogenous docosahexaenoic acid (dha) prevents aβ1–42 oligomer-induced neuronal injury. Mol. Neurobiol. 2015, 53, 3146–3153. [Google Scholar] [CrossRef] [PubMed]
- Lanzillotta, A.; Pignataro, G.; Branca, C.; Cuomo, O.; Sarnico, I.; Benarese, M.; Annunziato, L.; Spano, P.; Pizzi, M. Targeted acetylation of nf-kappab/rela and histones by epigenetic drugs reduces post-ischemic brain injury in mice with an extended therapeutic window. Neurobiol. Dis. 2013, 49, 177–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, J.H.; Surh, Y.J. Protective effect of resveratrol on beta-amyloid-induced oxidative PC12 cell death. Free Radic. Biol. Med. 2003, 34, 1100–1110. [Google Scholar] [CrossRef]
- Seo, J.-S.; Moon, M.-H.; Jeong, J.-K.; Seol, J.-W.; Lee, Y.-J.; Park, B.-H.; Park, S.-Y. Sirt1, a histone deacetylase, regulates prion protein-induced neuronal cell death. Neurobiol. Aging 2012, 33, 1110–1120. [Google Scholar] [CrossRef] [PubMed]
- Fu, P.; Peng, C.; Ding, J.Y.; Asmaro, K.; Sullivan, J.M.; Guthikonda, M.; Ding, Y. Acute administration of ethanol reduces apoptosis following ischemic stroke in rats. Neurosci. Res. 2013, 76, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Geng, X.; Parmar, S.; Li, X.; Peng, C.; Ji, X.; Chakraborty, T.; Li, W.A.; Du, H.; Tan, X.; Ling, F.; et al. Reduced apoptosis by combining normobaric oxygenation with ethanol in transient ischemic stroke. Brain Res. 2013, 1531, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Vlasova, Y.A.; Zakharova, I.O.; Avrova, N.F. The effects of alpha-tocoferol and H2O2 on the mitochondrial membrane potential and Bax/Bcl-xL ratio in PC12 cells. Neurochem. J. 2016, 10, 318–322. [Google Scholar] [CrossRef]
- Kerek, R.; Geoffroy, A.; Bison, A.; Martin, N.; Akchiche, N.; Pourié, G.; Helle, D.; Guéant, J.-L.; Bossenmeyer-Pourié, C.; Daval, J.-L. Early methyl donor deficiency may induce persistent brain defects by reducing stat3 signaling targeted by mir-124. Cell Death Dis. 2013, 4, e755. [Google Scholar] [CrossRef] [PubMed]
- Boue, S.M.; Wiese, T.E.; Nehls, S.; Burow, M.E.; Elliott, S.; Carter-Wientjes, C.H.; Shih, B.Y.; McLachlan, J.A. Evaluation of the estrogenic effects of legume extracts containing phytoestrogens. J. Agric. Food Chem. 2003, 51, 2193–2199. [Google Scholar] [CrossRef] [PubMed]
- Heinonen, S.-M.; Hoikkala, A.; Wähälä, K.; Adlercreutz, H. Metabolism of the soy isoflavones daidzein, genistein and glycitein in human subjects. J. Steroid Biochem. Mol. Biol. 2003, 87, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.; Fitzpatrick, L.A. Soy isoflavones: Are they useful in menopause? Mayo Clin. Proc. 2000, 75, 1174–1184. [Google Scholar] [CrossRef] [PubMed]
- Morito, K.; Hirose, T.; Kinjo, J.; Hirakawa, T.; Okawa, M.; Nohara, T.; Ogawa, S.; Inoue, S.; Muramatsu, M.; Masamune, Y. Interaction of phytoestrogens with estrogen receptors α and β. Biol. Pharm. Bull. 2001, 24, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Patisaul, H.B. Soy isoflavone supplements antagonize reproductive behavior and estrogen receptor and dependent gene expression in the brain. Endocrinology 2001, 142, 2946–2952. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.C.; Campana, A.; Lange, P.S.; Lee, H.-H.; Banerjee, K.; Bryson, J.B.; Mahishi, L.; Alam, S.; Giger, R.J.; Barnes, S.; et al. A large-scale chemical screen for regulators of the arginase 1 promoter identifies the soy isoflavone daidzeinas a clinically approved small molecule that can promote neuronal protection or regeneration via a camp-independent pathway. J. Neurosci. 2010, 30, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Schreihofer, D.A. High-soy diet decreases infarct size after permanent middle cerebral artery occlusion in female rats. AJP Regul. Integr. Comp. Physiol. 2005, 289, R103–R108. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.X.; Chen, W.F.; Xie, J.X.; Wong, M.S. Neuroprotective effects of genistein on dopaminergic neurons in the mice model of Parkinson’s disease. Neurosci Res. 2008, 60, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wei, H.; Cai, M.; Lu, Y.; Hou, W.; Yang, Q.; Dong, H.; Xiong, L. Genistein attenuates brain damage induced by transient cerebral ischemia through up-regulation of erk activity in ovariectomized mice. Int. J. Boil. Sci. 2014, 10, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Wang, X.Q.; Ding, C.; Du, X.L. Genistein attenuates isoflurane-induced neurotoxicity and improves impaired spatial learning and memory by regulating cAMP/CREB and BDNF-TrkB-PI3K/Akt signaling. Korean J. Physiol. Pharmacol. 2017, 21, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Grillot, D.A.; Gonzalez-Garcia, M.; Ekhterae, D.; Duan, L.; Inohara, N.; Ohta, S.; Seldin, M.F.; Nuñez, G. Genomic organization, promoter region analysis, and chromosome localization of the mouse Bcl-x gene. J. Immunol. 1997, 158, 4750–4757. [Google Scholar] [PubMed]
- Setchell, K.D.; Brown, N.M.; Zimmer-Nechemias, L.; Brashear, W.T.; Wolfe, B.E.; Kirschner, A.S.; Heubi, J.E. Evidence for lack of absorption of soy isoflavone glycosides in humans, supporting the crucial role of intestinal metabolism for bioavailability. Am. J. Clin. Nutr. 2002, 76, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.R.; Dudek, H.; Tao, X.; Masters, S.; Fu, H.; Gotoh, Y.; Greenberg, M.E. Akt phosphorylation of bad couples survival signals to the cell-intrinsic death machinery. Cell 1997, 91, 231–241. [Google Scholar] [CrossRef]
- Gardai, S.J.; Hildeman, D.A.; Frankel, S.K.; Whitlock, B.B.; Frasch, S.C.; Borregaard, N.; Marrack, P.; Bratton, D.L.; Henson, P.M. Xphosphorylation of bax ser184 by akt regulates its activity and apoptosis in neutrophils. J. Biol. Chem. 2004, 279, 21085–21095. [Google Scholar] [CrossRef] [PubMed]
- Koh, P.-O. Nicotinamide attenuates the ischemic brain injury-induced decrease of akt activation and bad phosphorylation. Neurosci. Lett. 2011, 498, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Demeter, M.R.; Ruan, H.; Comb, M.J. Bad ser-155 phosphorylation regulates bad/Bcl-xl interaction and cell survival. J. Biol. Chem. 2000, 275, 25865–25869. [Google Scholar] [CrossRef] [PubMed]
- Nunan, J.; Small, D.H. Regulation of APP cleavage by α-, β- and γ-secretases. FEBS Lett. 2000, 483, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Chow, V.W.; Mattson, M.P.; Wong, P.C.; Gleichmann, M. An overview of APP processing enzymes and products. Neuromol. Med. 2010, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Stone, J.R.; Okonkwo, D.O.; Singleton, R.H.; Mutlu, L.K.; Helm, G.A.; Povlishock, J.T. Caspase-3-mediated cleavage of amyloid precursor protein and formation of amyloid β peptide in traumatic axonal injury. J. Neurotrauma 2002, 19, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.C.; Soriano, S.; Bredesen, D.E.; Koo, E.H. Caspase cleavage of the amyloid precursor protein modulates amyloid β-protein toxicity. J. Neurochem. 2003, 87, 733–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carthy, C.M.; Yanagawa, B.; Luo, H.; Granville, D.J.; Yang, D.; Cheung, P.; Cheung, C.; Esfandiarei, M.; Rudin, C.M.; Thompson, C.B.; et al. Bcl-2 and Bcl-xL overexpression inhibits cytochrome c release, activation of multiple caspases, and virus release following coxsackievirus B3 infection. Virology 2003, 313, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Benedict, M.A.; Wu, D.; Inohara, N.; Nunez, G. Bcl-xl interacts with apaf-1 and inhibits apaf-1-dependent caspase-9 activation. Proc. Nat. Acad. Sci. USA 1998, 95, 4386–4391. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.P. The chemical constituents of ginseng plants. Comp. Med. East West. 1977, 5, 123–145. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.M.; Yao, Q.; Chen, C. Ginseng compounds: An update on their molecular mechanisms and medical applications. Curr. Vasc. Pharmacol. 2009, 7, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Hofseth, L.J.; Wargovich, M.J. Inflammation, cancer, and targets of ginseng. J Nutr. 2007, 37, 183S–185S. [Google Scholar] [CrossRef] [PubMed]
- Kitts, D.D.; Wijewickreme, A.N.; Hu, C. Antioxidant properties of a North American ginseng extract. Mol. Cell Biochem. 2000, 203, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.-Y.; Choi, I.-S.; Shim, J.-Y.; Yun, E.-K.; Yun, Y.-S.; Jeong, G.; Song, J.-Y. The immunomodulator ginsan induces resistance to experimental sepsis by inhibiting toll-like receptor-mediated inflammatory signals. Eur. J. Immunol. 2006, 36, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Keum, Y.-S.; Han, S.S.; Chun, K.-S.; Park, K.-K.; Park, J.-H.; Lee, S.K.; Surh, Y.-J. Inhibitory effects of the ginsenoside rg3 on phorbol ester-induced cyclooxygenase-2 expression, nf-κb activation and tumor promotion. Mutat. Res. Mol. Mech. Mutagen. 2003, 523–524, 75–85. [Google Scholar] [CrossRef]
- Xu, K.; Zhang, Y.; Wang, Y.; Ling, P.; Xie, X.; Jiang, C.; Zhang, Z.; Lian, X.-Y. Ginseng rb fraction protects glia, neurons and cognitive function in a rat model of neurodegeneration. PLoS ONE 2014, 9, e101077. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Sun, W.; Gong, W.; Ding, Y.; Zhuang, Y.; Hou, Q. Ginsenoside rg1 protects against transient focal cerebral ischemic injury and suppresses its systemic metabolic changes in cerabral injury rats. Acta Pharm. Sin. B 2015, 5, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, C.; Chen, S.; Li, Z.; Ma, L.; Jia, X.; Wang, K.; Bao, J.; Liang, Y.; Chen, M.W.; et al. Hormetic effect of panaxatriol saponins confers neuroprotection in PC12 cells and zebrafish through PI3K/AKT/mTOR and AMPK/SIRT1/FOXO3 pathways. Sci. Rep. 2017, 7, 41082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhang, R.-Y.; Zhao, J.; Dong, Z.; Feng, D.-Y.; Wu, R.; Shi, M.; Zhao, G. Ginsenoside rd protects sh-sy5y cells against 1-methyl-4-phenylpyridinium induced injury. Int. J. Mol. Sci. 2015, 16, 14395–14408. [Google Scholar] [CrossRef] [PubMed]
- Al Zaid Siddiquee, K.; Turkson, J. STAT3 as a target for inducing apoptosis in solid and hematological tumors. Cell Res. 2008, 18, 254–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karl, E.; Zhang, Z.; Dong, Z.; Neiva, K.G.; Soengas, M.S.; Koch, A.E.; Polverini, P.J.; Núñez, G.; Nör, J.E. Unidirectional crosstalk between Bcl-xl and Bcl-2 enhances the angiogenic phenotype of endothelial cells. Cell Death Differ. 2007, 14, 1657–1666. [Google Scholar] [CrossRef] [PubMed]
- Sathishkumar, N.; Sathiyamoorthy, S.; Ramya, M.; Yang, D.-U.; Lee, H.N.; Yang, D.-C. Molecular docking studies of anti-apoptotic Bcl-2, Bcl-xl, and mcl-1 proteins with ginsenosides frompanax ginseng. J. Enzym. Inhib. Med. Chem. 2011, 27, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Innis, S.M. Dietary (n-3) fatty acids and brain development. J. Nutr. 2007, 137, 855–859. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Kevala, K.; Kim, J.; Moon, H.S.; Jun, S.B.; Lovinger, D.; Kim, H.-Y. Docosahexaenoic acid promotes hippocampal neuronal development and synaptic function. J. Neurochem. 2009, 111, 510–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukiw, W.J.; Bazan, N.G. Docosahexaenoic acid and the aging brain. J. Nutr. 2008, 138, 2510–2514. [Google Scholar] [CrossRef] [PubMed]
- McNamara, R.K.; Carlson, S.E. Role of omega-3 fatty acids in brain development and function: Potential implications for the pathogenesis and prevention of psychopathology. Prostaglandins Leuk. Essent. Fat. Acids 2006, 75, 329–349. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, H.S.; Agrawal, R.; Sharma, S.; Huo, Y.-X.; Ying, Z.; Gomez-Pinilla, F. Omega-3 fatty acid deficiency during brain maturation reduces neuronal and behavioral plasticity in adulthood. PLoS ONE 2011, 6, e28451. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-L.; Chen, S.-J.; Kao, C.-L.; Hung, S.-C.; Ding, D.-C.; Yu, C.-C.; Chen, Y.-J.; Ku, H.-H.; Lin, C.-P.; Lee, K.-H.; et al. Docosahexaenoic acid promotes dopaminergic differentiation in induced pluripotent stem cells and inhibits teratoma formation in rats with parkinson-like pathology. Cell Transpl. 2012, 21, 313–332. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Bazan, N.G. Lipid-mediated cell signaling protects against injury and neurodegeneration. J Nutr. 2010, 140, 858–863. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.X.; Wang, J.; Wu, L.; Kang, Z.B. Transgenic mice: Fat-1 mice convert n-6 to n-3 fatty acids. Nature 2004, 427, 504. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.X. Fat-1 transgenic mice: A new model for omega-3 research. Prostaglandins Leuk. Essent. Fat. Acids 2007, 77, 263–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Upadhyay, G.; Kumar, S.; Kapoor, A.; Kumar, A.; Tiwari, M.; Godbole, M.M. Hypothyroidism alters the expression of Bcl-2 family genes to induce enhanced apoptosis in the developing cerebellum. J. Endocrinol. 2003, 176, 39–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Sinha, R.A.; Tiwari, M.; Pal, L.; Shrivastava, A.; Singh, R.; Kumar, K.; Kumar Gupta, S.; Godbole, M.M. Increased pro-nerve growth factor and p75 neurotrophin receptor levels in developing hypothyroid rat cerebral cortex are associated with enhanced apoptosis. Endocrinology 2006, 147, 4893–4903. [Google Scholar] [CrossRef] [PubMed]
- Antony, R.; Lukiw, W.J.; Bazan, N.G. Neuroprotectin d1 induces dephosphorylation of Bcl-xl in a pp2a-dependent manner during oxidative stress and promotes retinal pigment epithelial cell survival. J. Boil. Chem. 2010, 285, 18301–18308. [Google Scholar] [CrossRef] [PubMed]
- Bazan, N.G. Neuroprotectin d1-mediated anti-inflammatory and survival signaling in stroke, retinal degenerations, and alzheimer’s disease. J. Lipid Res. 2008, 50, S400–S405. [Google Scholar] [CrossRef] [PubMed]
- Megyesi, J.; Tarcsafalvi, A.; Seng, N.; Hodeify, R.; Price, P. Cdk2 phosphorylation of Bcl-xl after stress converts it to a pro-apoptotic protein mimicking bax/bak. Cell Death Discov. 2016, 2, 15066. [Google Scholar] [CrossRef] [PubMed]
- Cittelly, D.M.; Nesic-Taylor, O.; Perez-Polo, J.R. Phosphorylation of Bcl-xL after spinal cord injury. J. Neurosci. Res. 2007, 85, 1894–1911. [Google Scholar] [CrossRef] [PubMed]
- Veas-Perez de Tudela, M.; Delgado-Esteban, M.; Maestre, C.; Bobo-Jimenez, V.; Jimenez-Blasco, D.; Vecino, R.; Bolaños, J.P.; Almeida, A. Regulation of Bcl-xL-ATP synthase interaction by mitochondrial cyclin b1-cyclin-dependent kinase-1 determines neuronal survival. J. Neurosci. 2015, 35, 9287–9301. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; Goodwin, M.; Vu, T.; Brantley-Finley, C.; Gaarde, W.A.; Chambers, T.C. Vinblastine-induced phosphorylation of Bcl-2 and Bcl-xl is mediated by jnk and occurs in parallel with inactivation of the raf-1/mek/erk cascade. J. Biol. Chem. 2000, 275, 29980–29985. [Google Scholar] [CrossRef] [PubMed]
- Calderon, F.; Kim, H.Y. Docosahexaenoic acid promotes neurite growth in hippocampal neurons. J. Neurochem. 2004, 90, 979–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tosun, İ.; İnkaya, A.N. Resveratrol as a health and disease benefit agent. Food Rev. Int. 2010, 26, 85–101. [Google Scholar] [CrossRef]
- Khan, M.A.; Chen, H.; Wan, X.; Tania, M.; Xu, A.; Chen, F.; Zhang, D. Regulatory effects of resveratrol on antioxidant enzymes: A mechanism of growth inhibition and apoptosis induction in cancer cells. Mol. Cells 2013, 35, 219–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmatz, R.; Perreira, L.B.; Stefanello, N.; Mazzanti, C.; Spanevello, R.; Gutierres, J.; Bagatini, M.; Martins, C.C.; Abdalla, F.H.; da Silva Serres, J.D.; et al. Effects of resveratrol on biomarkers of oxidative stress and on the activity of delta aminolevulinic acid dehydratase in liver and kidney of streptozotocin-induced diabetic rats. Biochimie 2012, 94, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Kuršvietienė, L.; Stanevičienė, I.; Mongirdienė, A.; Bernatonienė, J. Multiplicity of effects and health benefits of resveratrol. Medicina 2016, 52, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.M.; Hsieh, T.C. Resveratrol: A cardioprotective substance. Ann. N. Y. Acad Sci. 2011, 1215, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Carter, L.G.; D’Orazio, J.A.; Pearson, K.J. Resveratrol and cancer: Focus on in vivo evidence. Endocr. Relat. Cancer 2014, 21, R209–R225. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Das, D.K. Anti-inflammatory responses of resveratrol. Inflamm. Allergy Drug Targets 2007, 6, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.A.; Lee, K.-E.; Kim, H.-S.; Park, E.-M. Acute Resveratrol treatment modulates multiple signaling pathways in the ischemic brain. Neurochem. Res. 2012, 37, 2686–2696. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Pang, L.; Fang, F.; Zhang, G.; Zhang, J.; Xie, M.; Wang, L. Resveratrol attenuates brain damage in a rat model of focal cerebral ischemia via up-regulation of hippocampal Bcl-2. Brain Res. 2012, 1450, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Koronowski, K.B.; Dave, K.R.; Saul, I.; Camarena, V.; Thompson, J.W.; Neumann, J.T.; Young, J.I.; Perez-Pinzon, M.A. Resveratrol preconditioning induces a novel extended window of ischemic tolerance in the mouse brain. Stroke 2015, 46, 2293–2298. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Gong, Q.; Dong, H.; Shi, J. Resveratrol, a neuroprotective supplement for Alzheimer’s disease. Curr. Pharm. Des. 2012, 18, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Ferretta, A.; Gaballo, A.; Tanzarella, P.; Piccoli, C.; Capitanio, N.; Nico, B.; Annese, T.; Di Paola, M.; Dell’Aquila, C.; De Mari, M.; et al. Effect of resveratrol on mitochondrial function: Implications in parkin-associated familiar Parkinson’s disease. Biochim. Biophys. Acta 2014, 1842, 902–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Edelstein, L.C.; Gelinas, C. The Rel/NF-kappaB family directly activates expression of the apoptosis inhibitor Bcl-x(L). Mol. Cell Biol. 2000, 20, 2687–2695. [Google Scholar] [CrossRef] [PubMed]
- Lanzillotta, A.; Sarnico, I.; Ingrassia, R.; Boroni, F.; Branca, C.; Benarese, M.; Faraco, G.; Blasi, F.; Chiarugi, A.; Spano, P.; et al. The acetylation of rela in lys310 dictates the nf-κb-dependent response in post-ischemic injury. Cell Death Dis. 2010, 1, e96. [Google Scholar] [CrossRef] [PubMed]
- Howitz, K.T.; Bitterman, K.J.; Cohen, H.Y.; Lamming, D.W.; Lavu, S.; Wood, J.G.; Zipkin, R.E.; Chung, P.; Kisielewski, A.; Zhang, L.-L.; et al. Small molecule activators of sirtuins extend saccharomyces cerevisiae lifespan. Nature 2003, 425, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Gertz, M.; Nguyen, G.T.; Fischer, F.; Suenkel, B.; Schlicker, C.; Franzel, B.; Tomaschewski, J.; Aladini, F.; Becker, C.; Wolters, D.; et al. A molecular mechanism for direct sirtuin activation by resveratrol. PLoS ONE 2012, 7, e49761. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Gao, H.; Zhang, W.; Zhang, W.; Wang, Y. Resveratrol alleviates nerve injury after cerebral ischemia and reperfusion in mice by inhibiting inflammation and apoptosis. Int. J. Clin. Exp. Med. 2015, 8, 3219–3226. [Google Scholar] [PubMed]
- Kizmazoglu, C.; Aydin, H.E.; Sevin, I.E.; Kalemci, O.; Yüceer, N.; Atasoy, M.A. Neuroprotective effect of resveratrol on acute brain ischemia reperfusion injury by measuring annexin v, p53, Bcl-2 levels in rats. J. Korean Neurosurg. Soc. 2015, 58, 508–512. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, Y.; Zhang, H.; Yan, H.; Wu, X.; Zhang, C. Administration of the resveratrol analogues isorhapontigenin and heyneanol-a protects mice hematopoietic cells against irradiation injuries. BioMed Res. Int. 2014, 2014, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, H.; Jia, G.; Li, L.; Chen, H.; Bi, J.; Wang, C.L. The Synergistic Neuroprotective Effects of Combined Rosuvastatin and Resveratrol Pretreatment against Cerebral Ischemia/Reperfusion Injury. J. Stroke Cerebrovasc. Dis. 2018, 27, 1697–1704. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, M.; Kumar, V.; Kashyap, M.P.; Khanna, V.K.; Randhawa, G.S.; Pant, A.B. Ischemic insult induced apoptotic changes in pc12 cells: Protection by trans resveratrol. Eur. J. Pharmacol. 2011, 666, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, J.S.; Mohr, C.; Rossi, D.J. Opposite actions of alcohol on tonic gabaa receptor currents mediated by nnos and pkc activity. Nat. Neurosci. 2013, 16, 1783–1793. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.; Fitzgerald, P.J.; MacPherson, K.P.; DeBrouse, L.; Colacicco, G.; Flynn, S.M.; Masneuf, S.; Pleil, K.E.; Li, C.; Marcinkiewcz, C.A.; et al. Chronic alcohol remodels prefrontal neurons and disrupts nmdar-mediated fear extinction encoding. Nat. Neurosci. 2012, 15, 1359–1361. [Google Scholar] [CrossRef] [PubMed]
- Taffe, M.A.; Kotzebue, R.W.; Crean, R.D.; Crawford, E.F.; Edwards, S.; Mandyam, C.D. Long-lasting reduction in hippocampal neurogenesis by alcohol consumption in adolescent nonhuman primates. Proc. Nat. Acad. Sci. USA 2010, 107, 11104–11109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stampfer, M.J.; Kang, J.H.; Chen, J.; Cherry, R.; Grodstein, F. Effects of moderate alcohol consumption on cognitive function in women. N. Engl. J. Med. 2005, 352, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Sacco, R.L.; Elkind, M.; Boden-Albala, B.; Lin, I.F.; Kargman, D.E.; Hauser, W.A.; Shea, S.; Paik, M.C. The protective effect of moderate alcohol consumption on ischemic stroke. JAMA 1999, 281, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Ruitenberg, A.; van Swieten, J.C.; Witteman, J.C.; Mehta, K.M.; van Duijn, C.M.; Hofman, A.; Breteler, M.M.B. Alcohol consumption and risk of dementia: The rotterdam study. Lancet 2002, 359, 281–286. [Google Scholar] [CrossRef]
- Ronksley, P.E.; Brien, S.E.; Turner, B.J.; Mukamal, K.J.; Ghali, W.A. Association of alcohol consumption with selected cardiovascular disease outcomes: A systematic review and meta-analysis. BMJ 2011, 342, d671. [Google Scholar] [CrossRef] [PubMed]
- Geng, X.; Fu, P.; Ji, X.; Peng, C.; Fredrickson, V.; Sy, C.; Meng, R.; Ling, F.; Du, H.; Tan, X.; et al. Synergetic neuroprotection of normobaric oxygenation and ethanol in ischemic stroke through improved oxidative mechanism. Stroke 2013, 44, 1418–1425. [Google Scholar] [CrossRef] [PubMed]
- Kochanski, R.; Peng, C.; Higashida, T.; Geng, X.; Hüttemann, M.; Guthikonda, M.; Ding, Y. Neuroprotection conferred by post-ischemia ethanol therapy in experimental stroke: An inhibitory effect on hyperglycolysis and nadph oxidase activation. J. Neurochem. 2013, 126, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Li, J.; Cao, X.; Chen, Y.; Ye, C.; Chen, K. Protective effect of n-butyl alcohol extracts from Rhizoma Pinelliae Pedatisectae against cerebral ischemia-reperfusion injury in rats. J. Ethnopharmacol. 2016, 188, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.-S.; Zhao, J.; Zheng, W.-P.; Zhao, Y. Neuroprotective effect of 4-hydroxybenzyl alcohol against transient focal cerebral ischemia via anti-apoptosis in rats. Brain Res. 2010, 1308, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Billen, L.P.; Kokoski, C.L.; Lovell, J.F.; Leber, B.; Andrews, D.W. Bcl-xl inhibits membrane permeabilization by competing with bax. PLoS Biol. 2008, 6, e147. [Google Scholar] [CrossRef] [PubMed]
- Siler-Marsiglio, K.I.; Madorsky, I.; Pan, Q.; Paiva, M.; Neeley, A.W.; Shaw, G.; Heaton, M.B. Effects of acute ethanol exposure on regulatory mechanisms of Bcl-2-associated apoptosis promoter, bad, in neonatal rat cerebellum: Differential effects during vulnerable and resistant developmental periods. Alcohol. Clin. Exp. Res. 2006, 30, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Han, J.Y.; Jeong, E.Y.; Kim, Y.S.; Roh, G.S.; Kim, H.J.; Kang, S.S.; Cho, G.J.; Choi, W.S. C-jun N-terminal kinase regulates the interaction between 14-3-3 and bad in ethanol-induced cell death. J. Neurosci. Res. 2008, 86, 3221–3229. [Google Scholar] [CrossRef] [PubMed]
- Bazovkina, D.V.; Tsybko, A.S.; Filimonova, E.A.; Ilchibaeva, T.V.; Naumenko, V.S. Influence of chronic alcohol treatment on the expression of the bdnf, bax, Bcl-xl, and casp3 genes in the mouse brain: Role of the c1473g polymorphism in the gene encoding tryptophan hydroxylase 2. Mol. Biol. 2016, 50, 262–269. [Google Scholar] [CrossRef]
- Lee, J.-H.; Tajuddin, N.F.; Druse, M.J. Effects of ethanol and ipsapirone on the expression of genes encoding anti-apoptotic proteins and an antioxidant enzyme in ethanol-treated neurons. Brain Res. 2009, 1249, 54–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Conversion of Bcl-xL to ∆N-Bcl-xL. During neurotoxic stimulation (e.g., cerebral ischemia), full length Bcl-xL undergoes caspase-dependent N-terminus cleavage to pro-apoptotic ∆N-Bcl-xL.
Figure 1.
Conversion of Bcl-xL to ∆N-Bcl-xL. During neurotoxic stimulation (e.g., cerebral ischemia), full length Bcl-xL undergoes caspase-dependent N-terminus cleavage to pro-apoptotic ∆N-Bcl-xL.


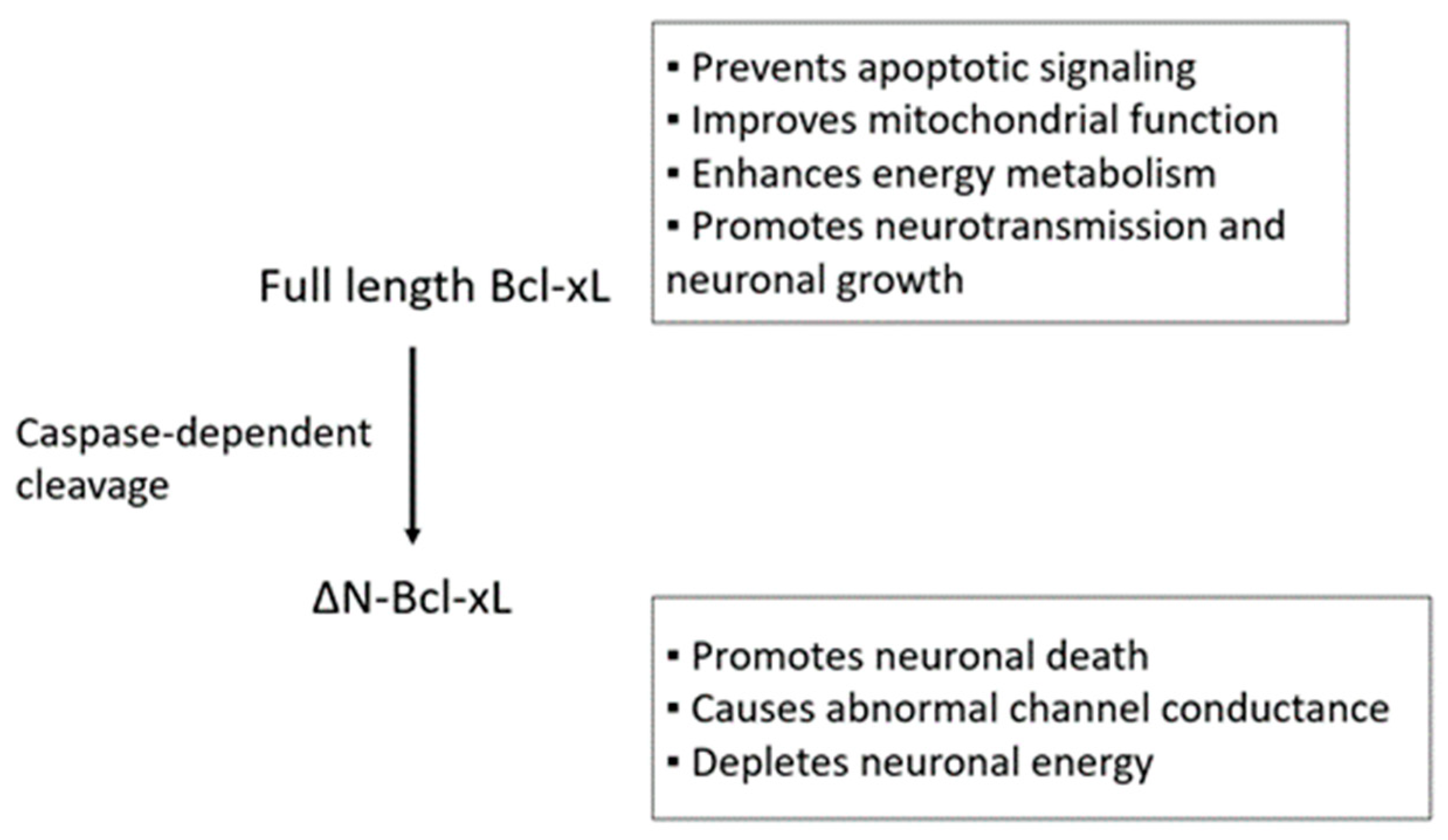{kind=link}
Table 1.
Neuroprotective properties of Bcl-xL in CNS-associated disease models.
| CNS-Associated Disorders | References |
|---|---|
| Ischemic stroke | [18,30,31,32,33,34,35] |
| Spinal cord injury | [36,37,38,39,40] |
| Alzheimer | [41,42,43,44] |
| Parkinson | [45,46,47,48,49,50] |
| Anxiety and Depression | [51,52,53] |
| Traumatic Brain injury | [54,55,56] |
| Spinal Muscular Dystrophy | [57,58,59,60,61] |
| Schizophrenia | [62,63,64] |
Table 2.
A list of dietary compounds that regulate Bcl-xL in the brain.
| Nutrients and Foods | Roles | References |
|---|---|---|
| High soy diet | Upregulation of Bcl-xL mRNA and protein in rats | [106] |
| High soy diet | Upregulation of Bcl-xL mRNA and protein in rats MCAO model | [107] |
| Orally gavaged soy isoflavones | Upregulation of Bcl-xL protein against Aβ-induced neurotoxicity in rats | [108] |
| Intravenous infusion of ginsenoside | STAT5-dependent upregulation of Bcl-xL protein in rodent ischemia models (rats and gerbils) and primary cortical neurons | [109] |
| Intravenous infusion of dihydroginsenoside Rb1 | Upregulation of Bcl-xL mRNA in spinal cord injury and cerebral ischemia models | [110] |
| Oral administration of red ginseng extract | Upregulation of Bcl-xL protein in rat spinal cord injury model | [111] |
| Oral administration and intravenous infusion of ginsenoside | Upregulation of Bcl-xL protein in mice during pneumococcal infection | [112] |
| Intravenous infusion of ginsenoside | Upregulation of Bcl-xL protein in rat spinal cord injury model | [113] |
| DHA or neuroprotectin D1 treatment in vitro | Upregulation of Bcl-xL gene in HN cell line | [114] |
| DHA treatment in vitro | Upregulation of Bcl-xL mRNA in HT22 cell line against MNNG-induced toxicity | [115] |
| Oral supplementation of DHA and EPA | Upregulation of Bcl-xL protein in hypothyroid rats | [116] |
| Oral supplementation of DHA | Upregulation of Bcl-xL mRNA in mice brains after challenge with dopaminergic toxin | [117] |
| Application with Fat-1 mouse model | Cortical neurons isolated from Fat-1 mice, genetically modified model with high endogenous omega-3 fatty acid levels, retained Bcl-xL protein against OGD | [118] |
| DHA treatment in vitro | Upregulation of Bcl-xL protein in primary cortical neurons against oxyhemoglobin treatment | [119] |
| Primary cortical neurons isolated from Fat-1 mice | Upregulation of Bcl-xL protein in primary cortical neurons against Aβ 1-42 treatment | [120] |
| Intraperitoneal delivery of resveratrol | Upregulation of acetylation of H3 histones on Bcl-xL promoter in mice | [121] |
| Resveratrol treatment in vitro | Upregulation of Bcl-xL protein in PC12 cells | [122] |
| Resveratrol treatment in vitro | Upregulation of Bcl-xL protein in SH-SY5Y cells | [123] |
| Intraperitoneal injection of ethanol | Upregulation of Bcl-xL protein in rat MCAO model | [124] |
| Intraperitoneal injection of ethanol | Upregulation of Bcl-xL protein in rat MCAO model | [125] |
| α-Tocopherol treatment in vitro | Downregulation of Bax/Bcl-xL protein ratio in PC 12 cells | [126] |
| Methyl donor deficient diet (no folate or vitamin B12) | Downregulation of Bcl-xL protein via miR-124-mediated Stat3 inhibition | [127] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Park, H.-A.; Broman, K.; Stumpf, A.; Kazyak, S.; Jonas, E.A. Nutritional Regulators of Bcl-xL in the Brain. Molecules 2018, 23, 3019. https://doi.org/10.3390/molecules23113019
AMA Style
Park H-A, Broman K, Stumpf A, Kazyak S, Jonas EA. Nutritional Regulators of Bcl-xL in the Brain. Molecules. 2018; 23(11):3019. https://doi.org/10.3390/molecules23113019
Chicago/Turabian StylePark, Han-A, Katheryn Broman, Allison Stumpf, Sara Kazyak, and Elizabeth A. Jonas. 2018. "Nutritional Regulators of Bcl-xL in the Brain" Molecules 23, no. 11: 3019. https://doi.org/10.3390/molecules23113019