Xylobiose Prevents High-Fat Diet Induced Mice Obesity by Suppressing Mesenteric Fat Deposition and Metabolic Dysregulation

 , ,
, , 

Abstract
:
1. Introduction
2. Results
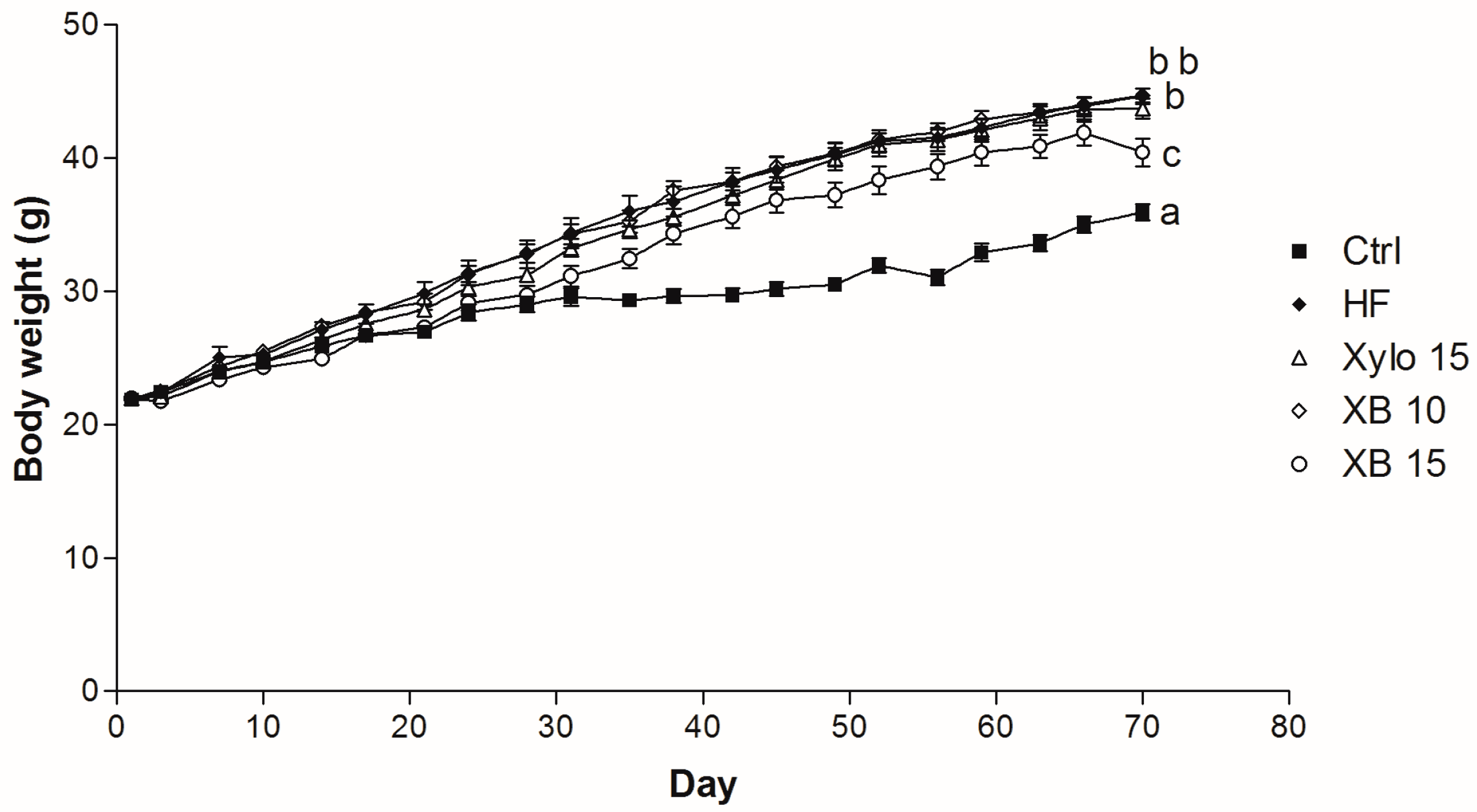
2.1. Effects of XB on Body Weight and Various Adipose Tissue Weights
2.2. Effects of XB on Blood Profiles
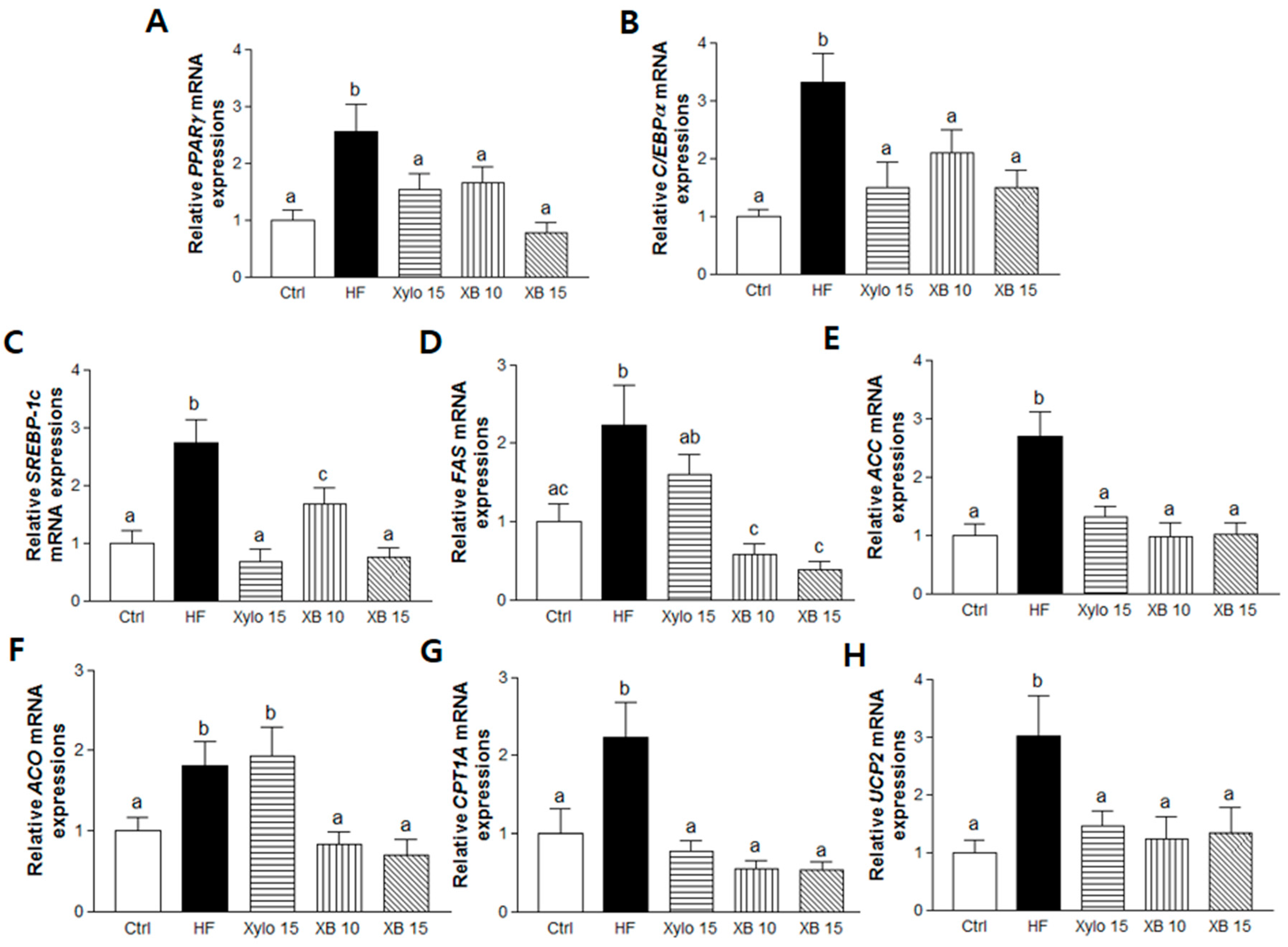
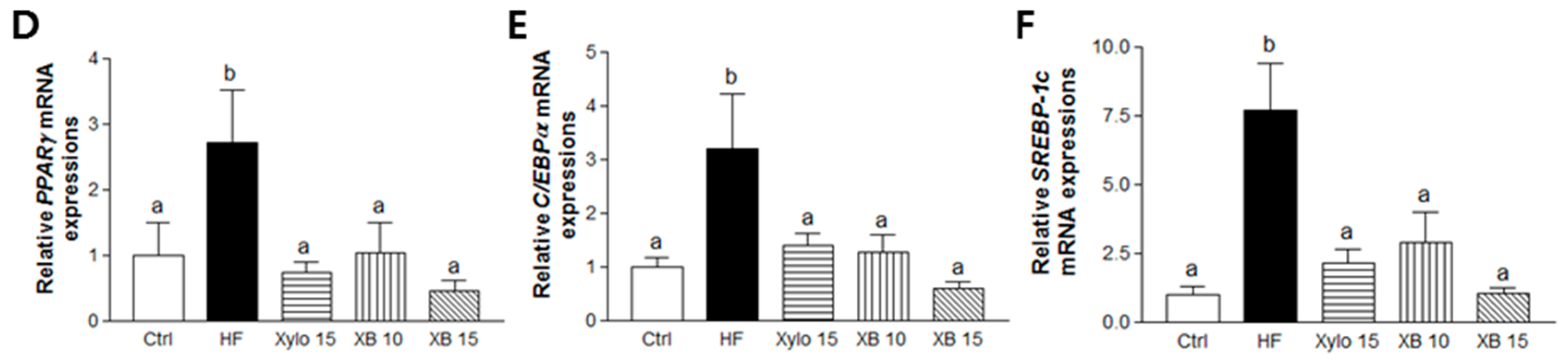
2.3. Effects of XB on Gene Expression Levels Related to Adipogenesis, Lipogenesis, and β-Oxidation in Mesenteric Adipose Tissues
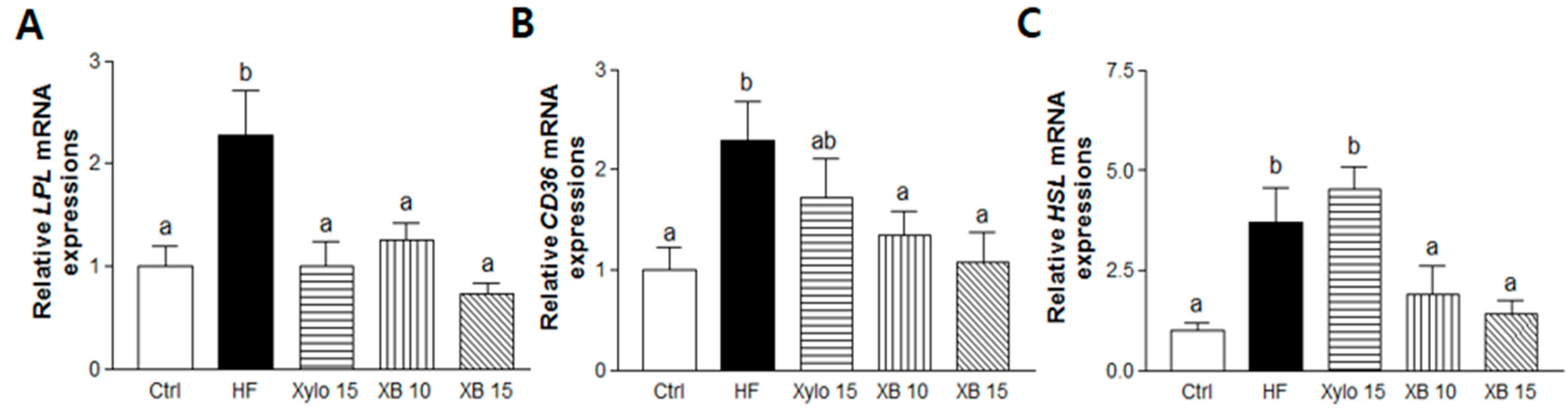
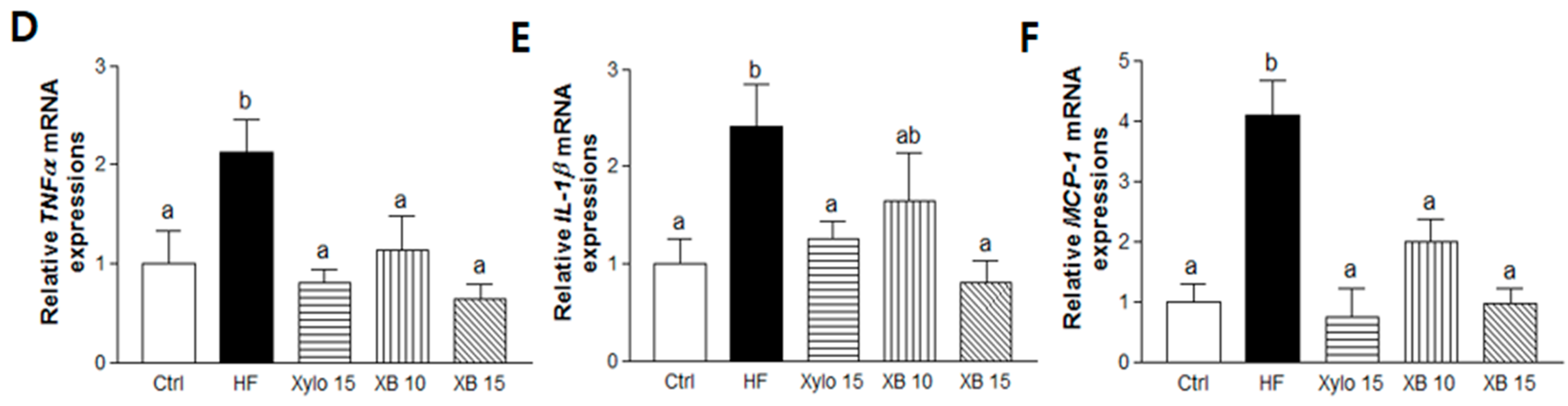
2.4. Effects of XB on Fatty Acid Uptake, Lipolysis, and Related Pro-Inflammatory Cytokines in Mesenteric Adipose Tissues
2.5. Effects of XB on Lipid Accumulation and Lipogenesis in the Liver
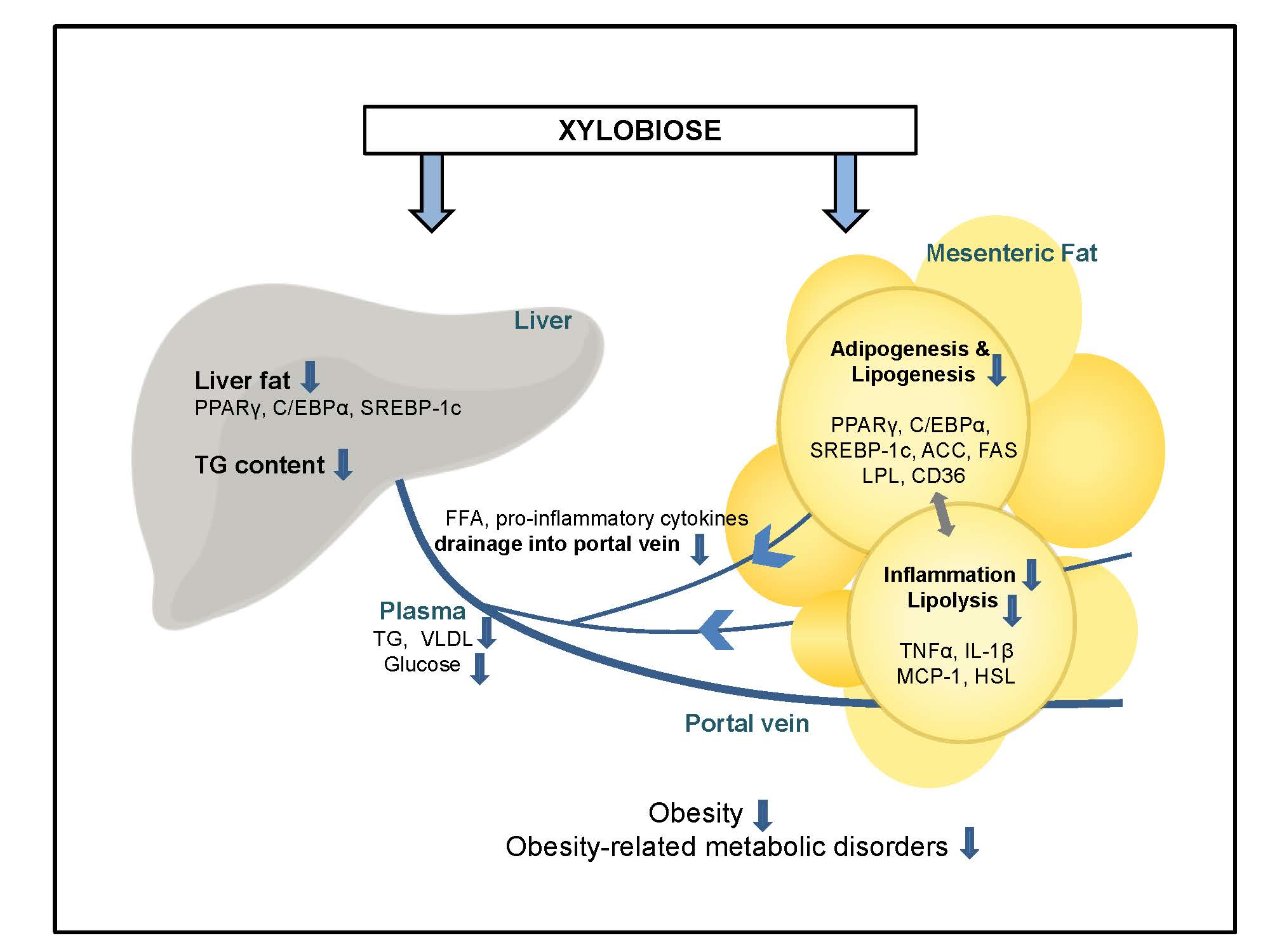
3. Discussion
4. Materials and Methods
4.1. XB Prepration
4.2. Animals and Diet
4.3. Blood and Tissue Sample Preparation
4.4. Biochemical Analysis of Blood Samples
4.5. RNA Isolation and Quantitative Real-Time-PCR (RT-PCR)
4.6. Histopathological Analysis and Liver TG Content Analysis
4.7. Statistical Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- World Health Organization (WHO). Global Health Observatory, Overweight and Obesity. Available online: http://www.who.int/gho/ncd/risk_factors/overweight/en/ (accessed on 18 May 2017).
- Faust, I.M.; Johnson, P.R.; Stern, J.S.; Hirsch, J. Diet-induced adipocyte number increase in adult rats: A new model of obesity. Am. J. Physiol. 1978, 235, E279–E286. [Google Scholar] [CrossRef] [PubMed]
- Jo, J.; Gavrilova, O.; Pack, S.; Jou, W.; Mullen, S.; Sumner, A.E.; Cushman, S.W.; Periwal, V. Hypertrophy and/or hyperplasia: Dynamics of adipose tissue growth. PLoS Comput. Biol. 2009, 5, e1000324. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Kokoeva, M.V.; Inouye, K.; Tzameli, I.; Yin, H.; Flier, J.S. Tlr4 links innate immunity and fatty acid-induced insulin resistance. J. Clin. Investig. 2006, 116, 3015–3025. [Google Scholar] [CrossRef] [PubMed]
- De Heredia, F.P.; Gomez-Martinez, S.; Marcos, A. Obesity, inflammation and the immune system. Proc. Nutr. Soc. 2012, 71, 332–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, M.G., Jr.; Leibel, R.L.; Seeley, R.J.; Schwartz, M.W. Obesity and leptin resistance: Distinguishing cause from effect. Trends Endocrinol. Metab. 2010, 21, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Poirier, P.; Giles, T.D.; Bray, G.A.; Hong, Y.; Stern, J.S.; Pi-Sunyer, F.X.; Eckel, R.H. Obesity and cardiovascular disease: Pathophysiology, evaluation, and effect of weight loss. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 968–976. [Google Scholar] [CrossRef] [PubMed]
- Klop, B.; Elte, J.W.; Cabezas, M.C. Dyslipidemia in obesity: Mechanisms and potential targets. Nutrients 2013, 5, 1218–1240. [Google Scholar] [CrossRef] [PubMed]
- Kersten, S. Mechanisms of nutritional and hormonal regulation of lipogenesis. EMBO Rep. 2001, 2, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Evans, K.; Burdge, G.C.; Wootton, S.A.; Clark, M.L.; Frayn, K.N. Regulation of dietary fatty acid entrapment in subcutaneous adipose tissue and skeletal muscle. Diabetes 2002, 51, 2684–2690. [Google Scholar] [CrossRef] [PubMed]
- Aalto-Setala, K.; Fisher, E.A.; Chen, X.; Chajek-Shaul, T.; Hayek, T.; Zechner, R.; Walsh, A.; Ramakrishnan, R.; Ginsberg, H.N.; Breslow, J.L. Mechanism of hypertriglyceridemia in human apolipoprotein (apo) CIII transgenic mice. Diminished very low density lipoprotein fractional catabolic rate associated with increased apo ciii and reduced apo e on the particles. J. Clin. Investig. 1992, 90, 1889–1900. [Google Scholar] [CrossRef] [PubMed]
- Poirier, P.; Giles, T.D.; Bray, G.A.; Hong, Y.; Stern, J.S.; Pi-Sunyer, F.X.; Eckel, R.H. Obesity and cardiovascular disease: Pathophysiology, evaluation, and effect of weight loss: An update of the 1997 american heart association scientific statement on obesity and heart disease from the obesity committee of the council on nutrition, physical activity, and metabolism. Circulation 2006, 113, 898–918. [Google Scholar] [PubMed]
- Steppan, C.M.; Bailey, S.T.; Bhat, S.; Brown, E.J.; Banerjee, R.R.; Wright, C.M.; Patel, H.R.; Ahima, R.S.; Lazar, M.A. The hormone resistin links obesity to diabetes. Nature 2001, 409, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Wellen, K.E.; Hotamisligil, G.S. Obesity-induced inflammatory changes in adipose tissue. J. Clin. Investig. 2003, 112, 1785–1788. [Google Scholar] [CrossRef] [PubMed]
- Konrad, D.; Wueest, S. The gut-adipose-liver axis in the metabolic syndrome. Physiology 2014, 29, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Dinh, C.H.; Szabo, A.; Yu, Y.; Camer, D.; Wang, H.; Huang, X.F. Bardoxolone methyl prevents mesenteric fat deposition and inflammation in high-fat diet mice. Sci. World J. 2015, 2015, 549352. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.H.; Chan, Y.L.; Chan, J.C.; Chan, W.B.; Kong, W.L. Mesenteric fat thickness as an independent determinant of fatty liver. Int. J. Obes. 2006, 30, 787–793. [Google Scholar] [CrossRef] [PubMed]
- De Wilde, J.; Smit, E.; Mohren, R.; Boekschoten, M.V.; de Groot, P.; van den Berg, S.A.; Bijland, S.; Voshol, P.J.; van Dijk, K.W.; de Wit, N.W.; et al. An 8-week high-fat diet induces obesity and insulin resistance with small changes in the muscle transcriptome of C57bL/6j mice. J. Nutrigenet. Nutrigenom. 2009, 2, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Mitsutomi, K.; Masaki, T.; Shimasaki, T.; Gotoh, K.; Chiba, S.; Kakuma, T.; Shibata, H. Effects of a nonnutritive sweetener on body adiposity and energy metabolism in mice with diet-induced obesity. Metabolism 2014, 63, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Maersk, M.; Belza, A.; Stødkilde-Jørgensen, H.; Ringgaard, S.; Chabanova, E.; Thomsen, H.; Pedersen, S.B.; Astrup, A.; Richelsen, B. Sucrose-sweetened beverages increase fat storage in the liver, muscle, and visceral fat depot: A 6-mo randomized intervention study. Am. J. Clin. Nutr. 2012, 95, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Gardner, C.; Wylie-Rosett, J.; Gidding, S.S.; Steffen, L.M.; Johnson, R.K.; Reader, D.; Lichtenstein, A.H. Nonnutritive sweeteners: Current use and health perspectives: A scientific statement from the American heart association and the American diabetes association. Diabetes Care 2012, 35, 1798–1808. [Google Scholar] [CrossRef] [PubMed]
- Seri, K.; Sanai, K.; Matsuo, N.; Kawakubo, K.; Xue, C.; Inoue, S. l-Arabinose selectively inhibits intestinal sucrase in an uncompetitive manner and suppresses glycemic response after sucrose ingestion in animals. Metabolism 1996, 45, 1368–1374. [Google Scholar] [CrossRef]
- Lim, E.; Lim, J.Y.; Shin, J.H.; Seok, P.R.; Jung, S.; Yoo, S.H.; Kim, Y. d-Xylose suppresses adipogenesis and regulates lipid metabolism genes in high-fat diet-induced obese mice. Nutr. Res. 2015, 35, 626–636. [Google Scholar] [CrossRef] [PubMed]
- Samanta, A.K.; Senani, S.; Kolte, A.P.; Sridhar, M.; Sampath, K.T.; Jayapal, N.; Devi, A. Production and in vitro evaluation of xylooligosaccharides generated from corn cobs. Food Bioprod. Process. 2012, 90, 466–474. [Google Scholar] [CrossRef]
- Okazaki, M.; Koda, M.; Izumi, R.; Fujikawa, S.; Matsumoto, N. In vitro digestibility and in vivo utilization of xylobiose. J. Jpn. Soc. Nutr. Food Sci. 1991, 9, 77–86. [Google Scholar] [CrossRef]
- Sheu, W.H.; Lee, I.T.; Chen, W.; Chan, Y.C. Effects of xylooligosaccharides in type 2 diabetes mellitus. J. Nutr. Sci. Vitaminol. 2008, 54, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Imaizumi, K.; Nakatsu, Y.; Sato, M.; Sedarnawati, Y.; Sugano, M. Effects of xylooligosaccharides on blood glucose, serum and liver lipids and cecum short-chain fatty acids in diabetic rats. Agric. Biol. Chem. 1991, 55, 199–205. [Google Scholar]
- Lin, S.H.; Chou, L.M.; Chien, Y.W.; Chang, J.S.; Lin, C.I. Prebiotic effects of xylooligosaccharides on the improvement of microbiota balance in human subjects. Gastroenterol. Res. Pract. 2016, 2016, 5789232. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Summanen, P.H.; Komoriya, T.; Finegold, S.M. In vitro study of the prebiotic xylooligosaccharide (XOS) on the growth of Bifidobacterium spp. and Lactobacillus spp. Int. J. Food Sci. Nutr. 2015, 66, 919–922. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.M.; Fahey, G.C., Jr.; Wolf, B.W. Selected indigestible oligosaccharides affect large bowel mass, cecal and fecal short-chain fatty acids, pH and microflora in rats. J. Nutr. 1997, 127, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.; Hsu, C.; Ko, C.; Chan, Y. Dietary intake of xylooligosaccharides improves the intestinal microbiota, fecal moisture, and ph value in the elderly. Nutr. Res. 2007, 27, 756–761. [Google Scholar] [CrossRef]
- Lim, E.; Lim, J.Y.; Kim, E.; Kim, Y.S.; Shin, J.H.; Seok, P.R.; Jung, S.; Yoo, S.H.; Kim, Y. Xylobiose, an alternative sweetener, ameliorates diabetes-related metabolic changes by regulating hepatic lipogenesis and miR-122a/33a in db/db mice. Nutrients 2016, 8, 791. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Maki, K.C.; Nieman, K.M.; Schild, A.L.; Kaden, V.N.; Lawless, A.L.; Kelley, K.M.; Rains, T.M. Sugar-sweetened product consumption alters glucose homeostasis compared with dairy product consumption in men and women at risk of type 2 diabetes mellitus. J. Nutr. 2015, 145, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Spiegelman, B.M. Adipocytes as regulators of energy balance and glucose homeostasis. Nature 2006, 444, 847–853. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, A.; Tumer, N.; Gao, Y.; Cheng, K.Y.; Scarpace, P.J. Prevention and reversal of diet-induced leptin resistance with a sugar-free diet despite high fat content. Br. J. Nutr. 2011, 106, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, M.J.; Alonso, J.L.; Domınguez, H.; Parajo, J.C. Xylooligosaccharides: Manufacture and applications. Trends Food Sci. Technol. 2000, 11, 387–393. [Google Scholar] [CrossRef]
- Manisseri, C.; Gudipati, M. Prebiotic activity of purified xylobiose obtained from ragi (Eleusine coracana, indaf-15) bran. Indian J. Microbiol. 2012, 52, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Dobiasova, M.; Frohlich, J. The plasma parameter log (TG/HDL-C) as an atherogenic index: Correlation with lipoprotein particle size and esterification rate in apob-lipoprotein-depleted plasma (FER(HDL)). Clin. Biochem. 2001, 34, 583–588. [Google Scholar] [CrossRef]
- Tan, M.H.; Johns, D.; Glazer, N.B. Pioglitazone reduces atherogenic index of plasma in patients with type 2 diabetes. Clin. Chem. 2004, 50, 1184–1188. [Google Scholar] [CrossRef] [PubMed]
- Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z. The metabolic syndrome. Lancet 2005, 365, 1415–1428. [Google Scholar] [CrossRef]
- Farooqi, I.S.; Matarese, G.; Lord, G.M.; Keogh, J.M.; Lawrence, E.; Agwu, C.; Sanna, V.; Jebb, S.A.; Perna, F.; Fontana, S.; et al. Beneficial effects of leptin on obesity, T cell hyporesponsiveness, and neuroendocrine/metabolic dysfunction of human congenital leptin deficiency. J. Clin. Investig. 2002, 110, 1093–1103. [Google Scholar] [CrossRef] [PubMed]
- Leon-Cabrera, S.; Solis-Lozano, L.; Suarez-Alvarez, K.; Gonzalez-Chavez, A.; Bejar, Y.L.; Robles-Diaz, G.; Escobedo, G. Hyperleptinemia is associated with parameters of low-grade systemic inflammation and metabolic dysfunction in obese human beings. Front. Integr. Neurosci. 2013, 7, 62. [Google Scholar] [CrossRef] [PubMed]
- Van Heek, M.; Compton, D.S.; France, C.F.; Tedesco, R.P.; Fawzi, A.B.; Graziano, M.P.; Sybertz, E.J.; Strader, C.D.; Davis, H.R., Jr. Diet-induced obese mice develop peripheral, but not central, resistance to leptin. J. Clin. Investig. 1997, 99, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Zimmet, P.Z.; Collins, V.R.; de Courten, M.P.; Hodge, A.M.; Collier, G.R.; Dowse, G.K.; Alberti, K.G.; Tuomilehto, J.; Hemraj, F.; Gareeboo, H.; et al. Is there a relationship between leptin and insulin sensitivity independent of obesity? A population-based study in the indian ocean nation of mauritius. Mauritius ncd study group. Int. J. Obes. Relat. Metab. Disord. 1998, 22, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Fanjul, C.; Barrenetxe, J.; Inigo, C.; Sakar, Y.; Ducroc, R.; Barber, A.; Lostao, M.P. Leptin regulates sugar and amino acids transport in the human intestinal cell line caco-2. Acta Physiol. 2012, 205, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Soukas, A.; Cohen, P.; Socci, N.D.; Friedman, J.M. Leptin-specific patterns of gene expression in white adipose tissue. Genes Dev. 2000, 14, 963–980. [Google Scholar] [PubMed]
- Chevillotte, E.; Giralt, M.; Miroux, B.; Ricquier, D.; Villarroya, F. Uncoupling protein-2 controls adiponectin gene expression in adipose tissue through the modulation of reactive oxygen species production. Diabetes 2007, 56, 1042–1050. [Google Scholar] [CrossRef] [PubMed]
- Kubota, N.; Terauchi, Y.; Yamauchi, T.; Kubota, T.; Moroi, M.; Matsui, J.; Eto, K.; Yamashita, T.; Kamon, J.; Satoh, H.; et al. Disruption of adiponectin causes insulin resistance and neointimal formation. J. Biol. Chem. 2002, 277, 25863–25866. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Hsu, C.H.; Wang, X.; Sakai, S.; Freeman, M.W.; Gonzalez, F.J.; Spiegelman, B.M. C/ebpalpha induces adipogenesis through ppargamma: A unified pathway. Genes Dev. 2002, 16, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Yang, J.; Horton, J.D.; Hammer, R.E.; Goldstein, J.L.; Brown, M.S. Diminished hepatic response to fasting/refeeding and liver X receptor agonists in mice with selective deficiency of sterol regulatory element-binding protein-1c. J. Biol. Chem. 2002, 277, 9520–9528. [Google Scholar] [CrossRef] [PubMed]
- Schutz, Y.; Tremblay, A.; Weinsier, R.L.; Nelson, K.M. Role of fat oxidation in the long-term stabilization of body weight in obese women. Am. J. Clin. Nutr. 1992, 55, 670–674. [Google Scholar] [CrossRef] [PubMed]
- Warfel, J.D.; Vandanmagsar, B.; Dubuisson, O.S.; Hodgeson, S.M.; Elks, C.M.; Ravussin, E.; Mynatt, R.L. Examination of carnitine palmitoyltransferase 1 abundance in white adipose tissue: Implications in obesity research. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 312, R816–R820. [Google Scholar] [CrossRef] [PubMed]
- Varanasi, U.; Chu, R.; Chu, S.; Espinosa, R.; LeBeau, M.M.; Reddy, J.K. Isolation of the human peroxisomal acyl-coa oxidase gene: Organization, promoter analysis, and chromosomal localization. Proc. Natl. Acad. Sci. USA 1994, 91, 3107–3111. [Google Scholar] [CrossRef] [PubMed]
- Fleury, C.; Neverova, M.; Collins, S.; Raimbault, S.; Champigny, O.; Levi-Meyrueis, C.; Bouillaud, F.; Seldin, M.F.; Surwit, R.S.; Ricquier, D.; et al. Uncoupling protein-2: A novel gene linked to obesity and hyperinsulinemia. Nat. Genet. 1997, 15, 269–272. [Google Scholar] [CrossRef] [PubMed]
- Reilly, J.M.; Thompson, M.P. Dietary fatty acids up-regulate the expression of ucp2 in 3t3-l1 preadipocytes. Biochem. Biophys. Res. Commun. 2000, 277, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Youm, Y.H.; Vandanmagsar, B.; Ravussin, A.; Gimble, J.M.; Greenway, F.; Stephens, J.M.; Mynatt, R.L.; Dixit, V.D. Obesity increases the production of proinflammatory mediators from adipose tissue t cells and compromises tcr repertoire diversity: Implications for systemic inflammation and insulin resistance. J. Immunol. 2010, 185, 1836–1845. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, S.; Manabe, I.; Nagasaki, M.; Eto, K.; Yamashita, H.; Ohsugi, M.; Otsu, M.; Hara, K.; Ueki, K.; Sugiura, S.; et al. Cd8+ effector T cells contribute to macrophage recruitment and adipose tissue inflammation in obesity. Nat. Med. 2009, 15, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Turrubiarte, G.; Gonzalez-Chavez, A.; Perez-Tamayo, R.; Salazar-Vazquez, B.Y.; Hernandez, V.S.; Garibay-Nieto, N.; Fragoso, J.M.; Escobedo, G. Severity of non-alcoholic fatty liver disease is associated with high systemic levels of tumor necrosis factor alpha and low serum interleukin 10 in morbidly obese patients. Clin. Exp. Med. 2016, 16, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Lapice, E.; Maione, S.; Patti, L.; Cipriano, P.; Rivellese, A.A.; Riccardi, G.; Vaccaro, O. Abdominal adiposity is associated with elevated C-reactive protein independent of BMI in healthy nonobese people. Diabetes Care 2009, 32, 1734–1736. [Google Scholar] [CrossRef] [PubMed]
- Day, C.P. Pathogenesis of steatohepatitis. Best Pract. Res. Clin. Gastroenterol. 2002, 16, 663–678. [Google Scholar] [CrossRef] [PubMed]
- Vroegrijk, I.O.; van Klinken, J.B.; van Diepen, J.A.; van den Berg, S.A.; Febbraio, M.; Steinbusch, L.K.; Glatz, J.F.; Havekes, L.M.; Voshol, P.J.; Rensen, P.C.; et al. Cd36 is important for adipocyte recruitment and affects lipolysis. Obesity (Silver Spring) 2013, 21, 2037–2045. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, A.M.; Orlando, R.A. Role of adipocyte-derived lipoprotein lipase in adipocyte hypertrophy. Nutr. Metab. 2007, 4, 22. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, I.J.; Eckel, R.H.; Abumrad, N.A. Regulation of fatty acid uptake into tissues: Lipoprotein lipase- and cd36-mediated pathways. J. Lipid Res. 2009, 50, S86–S90. [Google Scholar] [CrossRef] [PubMed]
- Klannemark, M.; Orho, M.; Langin, D.; Laurell, H.; Holm, C.; Reynisdottir, S.; Arner, P.; Groop, L. The putative role of the hormone-sensitive lipase gene in the pathogenesis of Type II diabetes mellitus and abdominal obesity. Diabetologia 1998, 41, 1516–1522. [Google Scholar] [CrossRef] [PubMed]
- Frayn, K.N.; Coppack, S.W.; Fielding, B.A.; Humphreys, S.M. Coordinated regulation of hormone-sensitive lipase and lipoprotein lipase in human adipose tissue in vivo: Implications for the control of fat storage and fat mobilization. Adv. Enzym. Regul. 1995, 35, 163–178. [Google Scholar] [CrossRef]
- Feldstein, A.E.; Canbay, A.; Guicciardi, M.E.; Higuchi, H.; Bronk, S.F.; Gores, G.J. Diet associated hepatic steatosis sensitizes to fas mediated liver injury in mice. J. Hepatol. 2003, 39, 978–983. [Google Scholar] [CrossRef]
- Thorne, A.; Lofgren, P.; Hoffstedt, J. Increased visceral adipocyte lipolysis—A pathogenic role in nonalcoholic fatty liver disease? J. Clin. Endocrinol. Metab. 2010, 95, E209–E213. [Google Scholar] [CrossRef] [PubMed]
- Bae, Y.J.; Bak, Y.K.; Kim, B.; Kim, M.S.; Lee, J.H.; Sung, M.K. Coconut-derived d-xylose affects postprandial glucose and insulin responses in healthy individuals. Nutr. Res. Pract. 2011, 5, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef] [PubMed]
- An, H.M.; Park, S.Y.; Lee, D.K.; Kim, J.R.; Cha, M.K.; Lee, S.W.; Lim, H.T.; Kim, K.J.; Ha, N.J. Antiobesity and lipid-lowering effects of Bifidobacterium spp. In high fat diet-induced obese rats. Lipids Health Dis. 2011, 10, 116. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Choi, J.; Kyung, M.; Seo, S.; Jo, S.-E.; Lee, K.; Kim, P.; Wang, N.-H.L.; Jung, S.; Mun, S. Application of bacillus pumilus β-xylosidase reaction and simulated moving bed purification to efficient production of high-purity xylobiose from xylose. J. Ind. Eng. Chem. 2017, 47, 431–438. [Google Scholar] [CrossRef]
- Davidson, M.; Liu, S.X.; Barter, P.; Brinton, E.A.; Cannon, C.P.; Gotto, A.M., Jr.; Leary, E.T.; Shah, S.; Stepanavage, M.; Mitchel, Y.; et al. Measurement of ldl-c after treatment with the cetp inhibitor anacetrapib. J. Lipid Res. 2013, 54, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Onat, A.; Can, G.; Kaya, H.; Hergenc, G. “Atherogenic index of plasma” (log10 triglyceride/high-density lipoprotein-cholesterol) predicts high blood pressure, diabetes, and vascular events. J. Clin. Lipidol. 2010, 4, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Jouihan, H. Measurement of liver triglyceride content. Bio-Protocol 2012, 2, e223. [Google Scholar] [CrossRef]
Sample Availability: Not available. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ctrl | HF | Xylo 15 | XB 10 | XB 15 | |
|---|---|---|---|---|---|
| Final body weight (g) | 35.93 ± 0.60 a | 44.64 ± 0.58 b | 43.72 ± 0.75 b | 44.69 ± 0.51 b | 41.23 ± 0.76 c |
| Food intake (g/day)Mesenteric adipose tissue (g) | 3.32 ± 0.04 | 2.76 ± 0.03 | 2.87 ± 0.32 | 2.75 ± 0.03 | 2.89 ± 0.09 |
| 0.56 ± 0.03 a | 1.4 ± 0.05 b | 1.3 ± 0.08 b | 1.38 ± 0.06 b | 0.99 ± 0.09 c | |
| Subcutaneous adipose tissue (g) | 0.81 ± 0.11 a | 2.33 ± 0.11 b | 2.07 ± 0.12 bc | 1.91 ± 0.12 c | 1.81 ± 0.17 c |
| Perirenal adipose tissue (g) | 0.59 ± 0.04 a | 0.95 ± 0.02 b | 0.94 ± 0.04 b | 0.94 ± 0.03 b | 0.80 ± 0.04 c |
| Epididymal adipose tissue (g) | 1.31 ± 0.07 a | 1.85 ± 0.07 b | 2.04 ± 0.09 b | 1.93 ± 0.05 b | 1.91 ± 0.06 b |
| Total adipose tissue (g) | 3.27 ± 0.22 a | 6.53 ± 0.14 b | 6.35 ± 0.24 b | 6.15 ± 0.17 b | 5.52 ± 0.30 c |
| Ctrl | HF | Xylo 15 | XB 10 | XB 15 | |
|---|---|---|---|---|---|
| Fasting blood glucose (mg/dL) | 75.00 ± 5.92 a | 188.46 ± 8.77 b | 146.77 ± 5.88 c | 171.62 ± 6.44 b | 130.75 ± 5.97 c |
| Plasma insulin (μg/L) | 0.44 ± 0.04 a | 3.24 ± 0.16 b | 1.97 ± 0.12 c | 1.83 ± 0.10 c | 1.56 ± 0.10 c |
| HOMA-IR | 1.50 ± 0.17 a | 27.00 ± 1.55 b | 12.79 ± 0.92 c | 14.00 ± 0.90 c | 8.97 ± 0.74 d |
| Triglyceride (mg/dL) | 150.61 ± 11.98 a | 204.6 ± 3.42 b | 123.55 ± 5.76 c | 119.7 ± 3.84 c | 119.67 ± 3.41 c |
| Total cholesterol (mg/dL) | 110.69 ± 3.97 a | 151.67 ± 6.54 b | 132.07 ± 4.94 c | 126.03 ± 3.34 ac | 112.56 ± 4.91 a |
| LDL-cholesterol (mg/dL) | 4.37 ± 4.01 a | 43.62 ± 7.46 b | 27.55 ± 4.49 c | 16.61 ± 2.56 a,c | 10.94 ± 3.1 a |
| VLDL-cholesterol (mg/dL) | 30.12 ± 2.4 a | 40.92 ± 0.69 b | 24.71 ± 1.15 c | 23.94 ± 0.77 c | 23.93 ± 0.68 c |
| HDL-cholesterol (mg/dL) | 76.2 ± 3.34 a,b | 67.13 ± 3.41 a | 79.81 ± 4.13 a,b | 85.48 ± 2.51 b | 77.69 ± 4.1 a,b |
| Atherogenic index | 0.29 ± 0.04 a | 0.49 ± 0.02 b | 0.19 ± 0.03 c | 0.15 ± 0.02 c | 0.19 ± 0.02 c |
| Leptin (ng/mL) | 14.44 ± 1.49 a | 30.85 ± 0.84 b | 29.35 ± 1.44 b,c | 29.42 ± 0.54 b,c | 25.54 ± 1.42 c |
| Adiponectin (ng/mL) | 1.11 ± 0.03 a | 0.84 ± 0.02 b | 0.98 ± 0.03 c | 0.88 ± 0.02 b | 0.96 ± 0.04 c |
| Ingredient | AIN-93G 1 | High Fat 2 | Xylo 15 3 | XB 10 4 | XB 15 5 |
|---|---|---|---|---|---|
| (g) | (g) | (g) | (g) | (g) | |
| Casein, lactic | 200 | 200 | 200 | 200 | 200 |
| l-Cystine | 3 | 3 | 3 | 3 | 3 |
| Corn starch | 397.5 | 0 | 0 | 0 | 0 |
| Maltodextrin | 132 | 116.4 | 116.4 | 116.4 | 116.4 |
| Sucrose | 100 | 77.4 | 65.8 | 69.6 | 65.8 |
| d-Xylose | 0 | 0 | 11.6 | 0 | 0 |
| Xylobiose | 0 | 0 | 0 | 7.7 | 11.6 |
| Cellulose | 50 | 50 | 50 | 50 | 50 |
| Soybean oil | 70 | 25 | 25 | 25 | 25 |
| Lard | - | 245 | 245 | 245 | 245 |
| Mineral mix | 35 | 10 | 10 | 10 | 10 |
| Dicalcium phosphate | - | 13 | 13 | 13 | 13 |
| Calcium carbonate | - | 5.5 | 5.5 | 5.5 | 5.5 |
| Potassium citrate·H2O | - | 16.5 | 16.5 | 16.5 | 16.5 |
| Vitamin mix | 10 | 10 | 10 | 10 | 10 |
| Choline bitartrate | 2.5 | 2 | 2 | 2 | 2 |
| Total amount | 1000 | 773.85 | 773.85 | 773.75 | 773.85 |
| Energy (kcal/g) | 4 | 5.24 | 5.24 | 5.24 | 5.24 |
| Genes | Gene Symbol | GenBank ID | Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) |
|---|---|---|---|---|
| ACC | Acaca | 107476 | AGGATTTGCTGTTTCTCAGAGCTT | CAGGATCTACCCAGGCCACAT |
| ACO | Acox1 | 11430 | TTGGAAACCACTGCCACATA | AGGCATGTAACCCGTAGCAC |
| CD36 | Cd36 | 12491 | GTGCTCTCCCTTGATTCTGC | TGAGAATGCCTCCAAACACA |
| C/EBPα | Cebpa | 12606 | CCAAGAAGTCGGTGGACAAGA | CGGTCATTGTCACTGGTCAACT |
| CPT1A | Cpt1a | 12894 | AACCCAGTGCCTTAACGATG | GAACTGGTGGCCAATGAGAT |
| FAS | Fasn | 14104 | TGTGAGTGGTTCAGAGGCAT | TTCTGTAGTGCCAGCAAGCT |
| HSL | Lipe | 16890 | GGGCTGTCAAGCACTGT | GTAACTGGGTAGGCTGCCAT |
| IL-1β | Il1b | 16176 | ATGGCAACTGTTCCTGAACTCAACT | CAGGACAGGTATAGATTCTTTCCTTT |
| MCP-1 | Mcpt1 | 17224 | CCCACTCACCTGCTGCTACT | TCTGGACCCATTCCTTCTTG |
| LPL | Lpl | 16956 | TCCCGTTACCGTCCATCC | GAGTTTGACCGCCTTCCG |
| PPARα | Ppara | 19013 | CTCTGGGCAAGAGAATCCAC | ACTGGCAGCAGTGGAAGAAT |
| PPARγ | Pparg | 19016 | GAGCACTTCACAAGAAATTACC | GAACTCCATAGTGGAAGCCT |
| SREBP-1c | Srebf1 | 20787 | TAGAGCATATCCCCCAGGTG | GGTACGGGCCACAAGAAGTA |
| TNFα | Tnfaip1 | 21926 | TACGCCACAGAGAAGAAGCA | TGGCCTCCAGTAAGGAATTG |
| UCP2 | Ucp2 | 22228 | CACTTTCCCTCTGGATACCG | GTGCTCTGGTATCTCCGACC |
| GAPDH | Gapdh | 14433 | AACTTTGGCATTGTGGAAGG | TGTGAGGGAGATGCTCAGTG |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, S.-m.; Kim, E.; Shin, J.-H.; Seok, P.R.; Jung, S.; Yoo, S.-H.; Kim, Y. Xylobiose Prevents High-Fat Diet Induced Mice Obesity by Suppressing Mesenteric Fat Deposition and Metabolic Dysregulation. Molecules 2018, 23, 705. https://doi.org/10.3390/molecules23030705
Lim S-m, Kim E, Shin J-H, Seok PR, Jung S, Yoo S-H, Kim Y. Xylobiose Prevents High-Fat Diet Induced Mice Obesity by Suppressing Mesenteric Fat Deposition and Metabolic Dysregulation. Molecules. 2018; 23(3):705. https://doi.org/10.3390/molecules23030705
Chicago/Turabian StyleLim, Soo-min, Eunju Kim, Jae-Ho Shin, Pu Reum Seok, Sangwon Jung, Sang-Ho Yoo, and Yuri Kim. 2018. "Xylobiose Prevents High-Fat Diet Induced Mice Obesity by Suppressing Mesenteric Fat Deposition and Metabolic Dysregulation" Molecules 23, no. 3: 705. https://doi.org/10.3390/molecules23030705
APA StyleLim, S.-m., Kim, E., Shin, J.-H., Seok, P. R., Jung, S., Yoo, S.-H., & Kim, Y. (2018). Xylobiose Prevents High-Fat Diet Induced Mice Obesity by Suppressing Mesenteric Fat Deposition and Metabolic Dysregulation. Molecules, 23(3), 705. https://doi.org/10.3390/molecules23030705