
Antioxidant and ACE Inhibitory Activity of Enzymatic Hydrolysates from Ruditapes philippinarum


Abstract
:1. Introduction
2. Results and Discussions
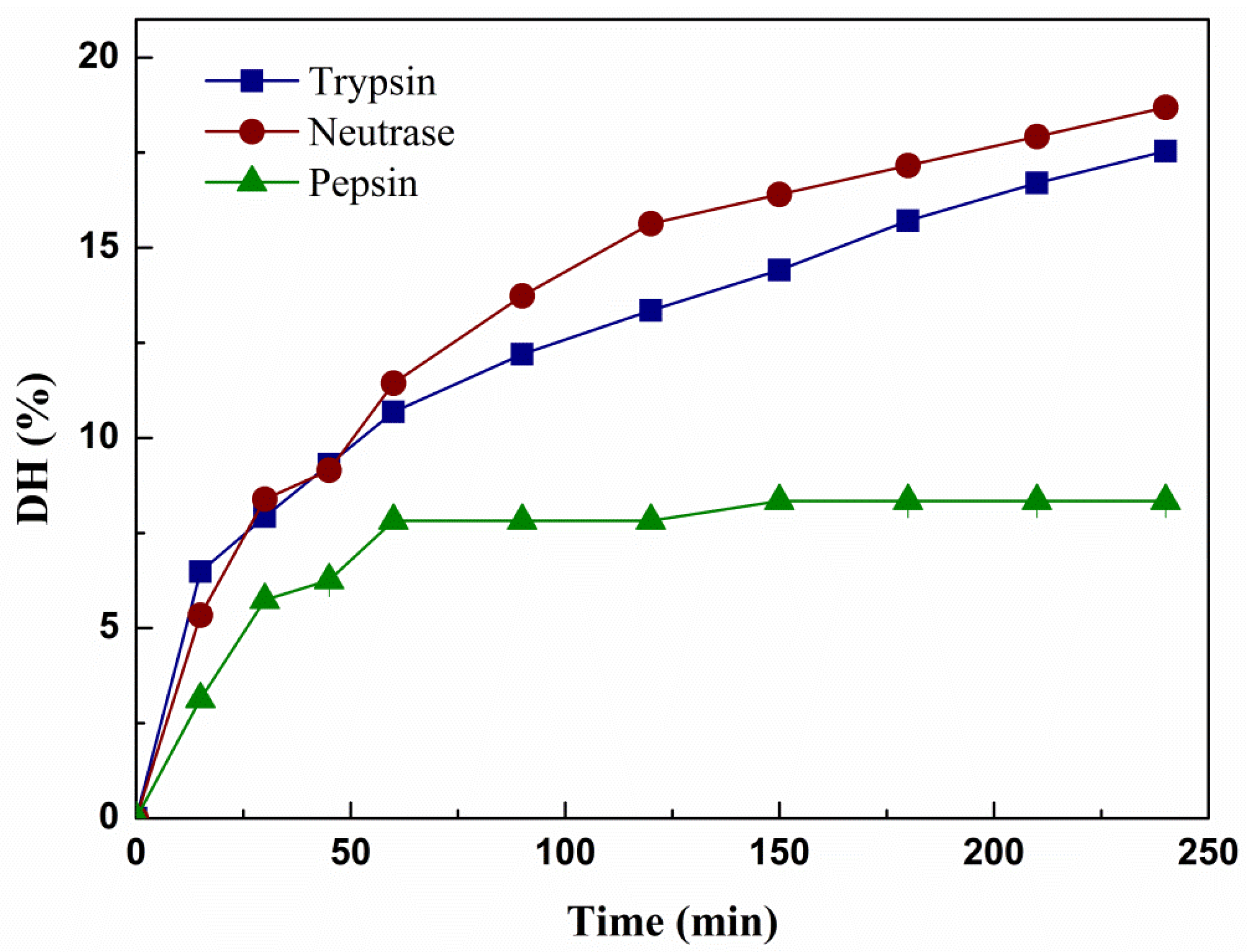
2.1. Typical Characteristics of Hydrolysates
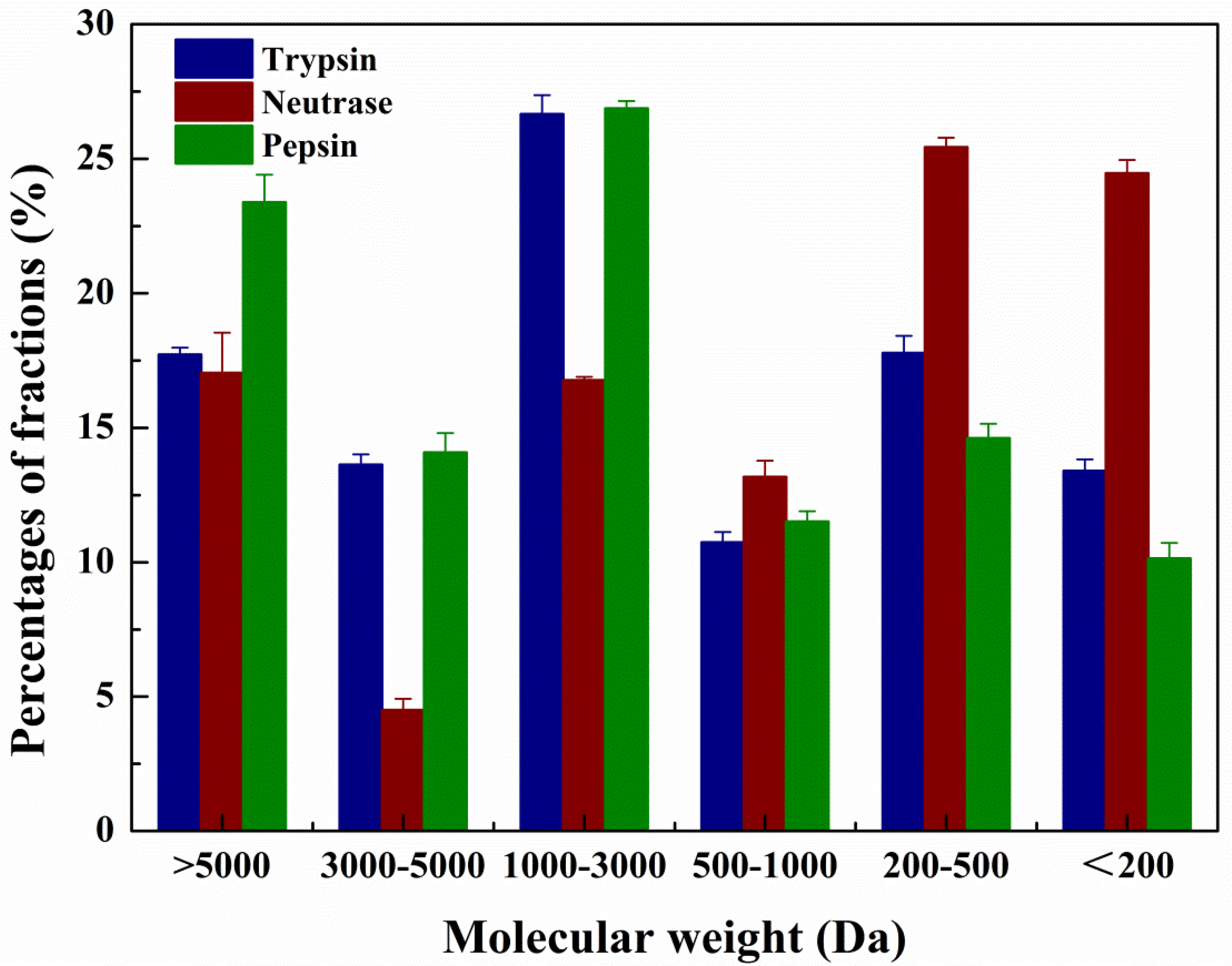
2.2. MW Distribution of the Hydrolysates
2.3. Amino Acid Composition Analysis
2.4. Inhibitory Activity on Linoleic Acid Peroxidation
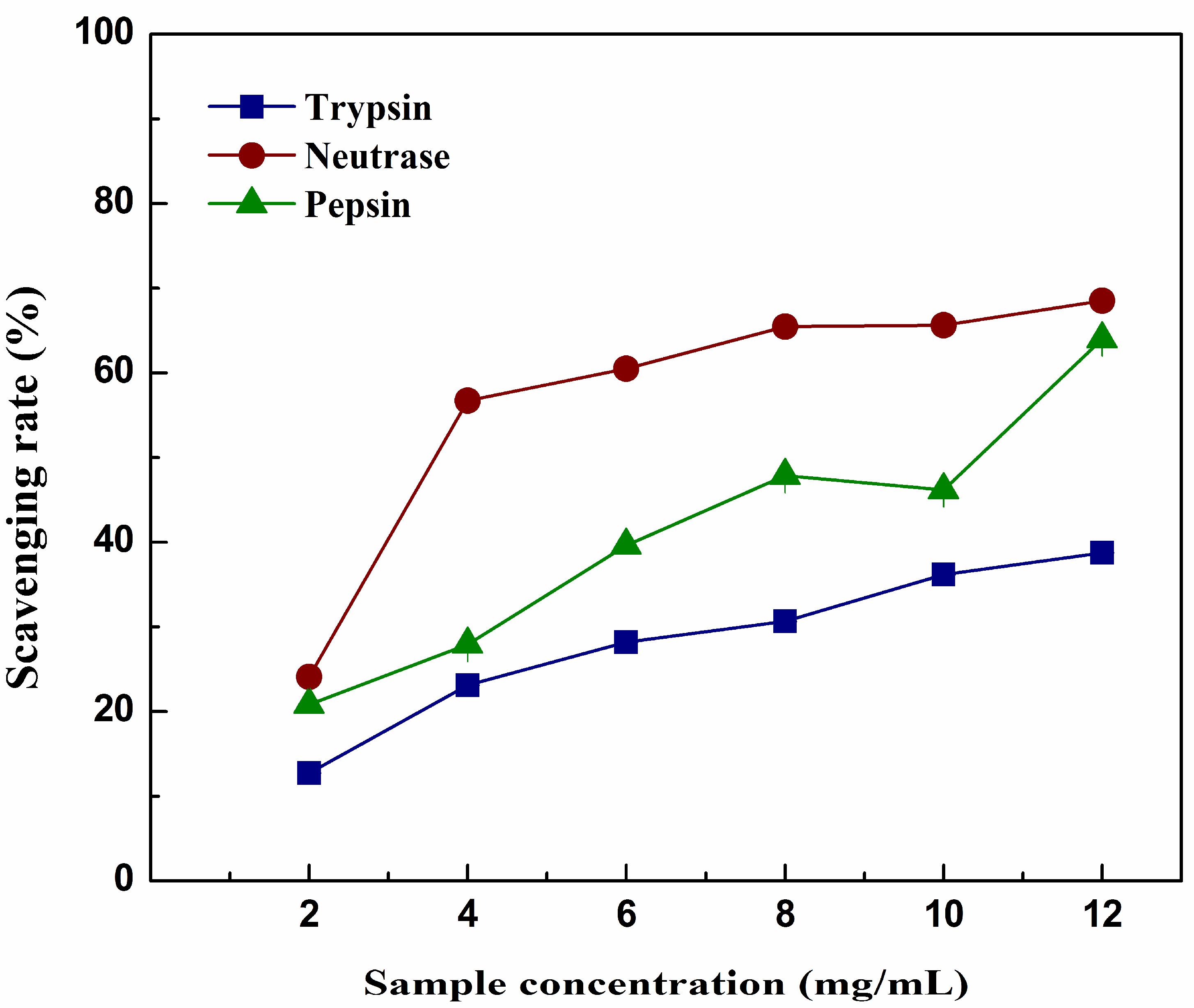
2.5. DPPH Scavenging Activity
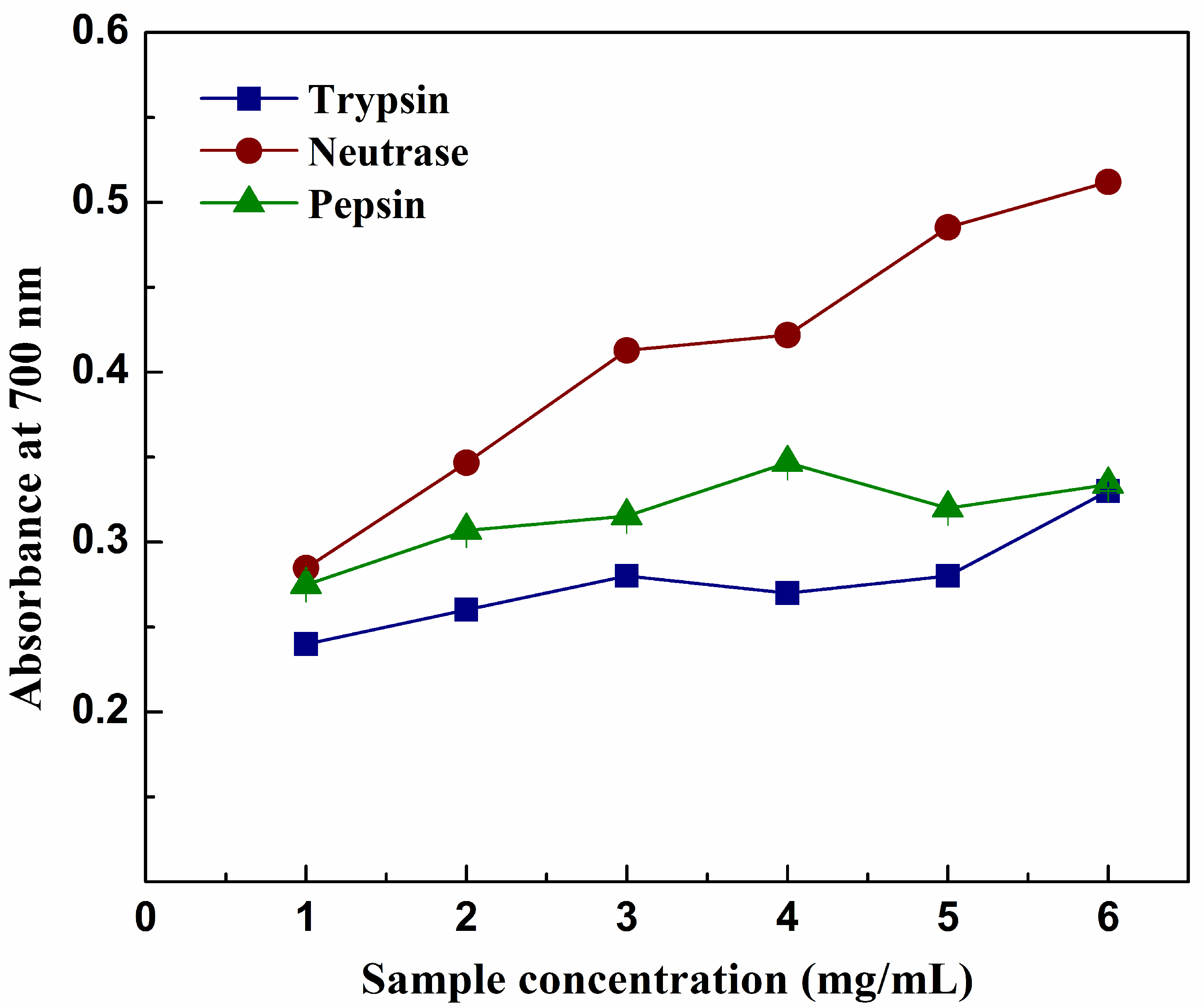
2.6. Reducing Power
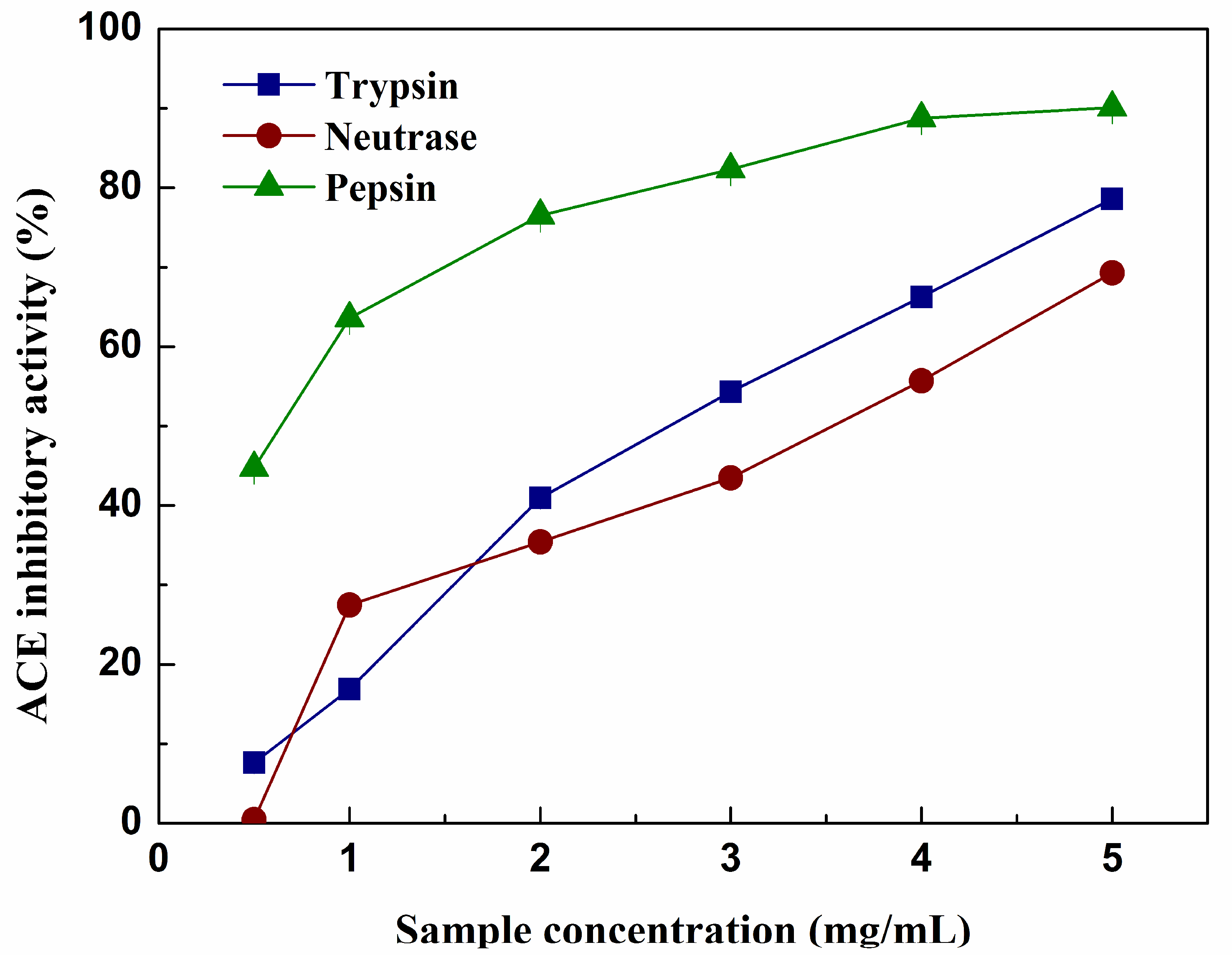
2.7. ACE inhibitory Activity
3. Materials and Methods
3.1. Materials
3.2. Preparation of Enzymatic Hydrolysates from R. philippinarum
3.3. Proximate Compositions Analysis
3.4. Molecular Weight Distribution
3.5. Determination of Free Amino Acid Composition of RPHs
3.6. Inhibitory Activity on Linoleic Acid Peroxidation
3.7. DPPH Scavenging Activity
3.8. Reducing Power
3.9. Angiotensin I-Converting Enzyme (ACE) Inhibition Assay
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Kim, S.K.; Wijesekara, I. Development and biological activities of marine-derived bioactive peptides: A review. J. Funct. Foods 2010, 2, 1–9. [Google Scholar] [CrossRef]
- Harnedy, P.A.; Fitzgerald, R.J. Bioactive peptides from marine processing waste and shellfish: A review. J. Funct. Foods 2012, 4, 6–24. [Google Scholar] [CrossRef]
- Murray, B.; Fitzgerald, R. Angiotensin converting enzyme inhibitory peptides derived from food proteins: Biochemistry, bioactivity and production. Curr. Pharm. Des. 2007, 13, 773–791. [Google Scholar] [CrossRef] [PubMed]
- Meisel, H. Multifunctional peptides encrypted in milk proteins. Biofactors 2004, 21, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Chung, E.Y.; Ryou, D.K.; Lee, J.H. Gonadal Development, Age and Growth of the Shortnecked Clam, Ruditapes philippinarum (Pelecypoda: Veneridae), on the Coast of Kimje, Korea. Korean J. Malacol. 1994, 10, 38–54. [Google Scholar]
- Cordero, D.; Delgado, M.; Liu, B.; Ruesink, J.; Saavedra, C. Population genetics of the Manila clam (Ruditapes philippinarum) introduced in North America and Europe. Sci. Rep. 2017, 7, 39745. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Kim, E.K.; Kim, Y.S.; Hwang, J.W.; Lee, K.H.; Choi, D.K.; Park, P.J. Purification and characterization of a nitric oxide inhibitory peptide from Ruditapes philippinarum. Food Chem. Toxicol. 2012, 50, 1660–1666. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Hwang, J.W.; Kim, Y.S.; Ahn, C.B.; Jeon, Y.J. A novel bioactive peptide derived from enzymatic hydrolysis of Ruditapes philippinarum: Purification and investigation of its free-radical quenching potential. Process Biochem. 2013, 48, 325–330. [Google Scholar] [CrossRef]
- Ames, B.N. Micronutrients prevent cancer and delay aging. Toxicol. Lett. 1998, 102–103, 5–18. [Google Scholar] [CrossRef]
- Ames, B.N.; Shigenaga, M.K.; Hagen, T.M. Oxidants, antioxidants, and the degenerative diseases of aging. Proc. Natl. Acad. Sci. USA 1993, 90, 7915–7922. [Google Scholar] [CrossRef] [PubMed]
- Hernándezledesma, B.; Dávalos, A.; Begoña Bartolomé, A.; Amigo, L. Preparation of Antioxidant Enzymatic Hydrolysates from α-Lactalbumin and β-Lactoglobulin. Identification of Active Peptides by HPLC-MS/MS. J. Agric. Food Chem. 2005, 53, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Ito, N.; Fukushima, S.; Tsuda, H. Carcinogenicity and Modification of the Carcinogenic Response by BHA, BHT, and Other Antioxidants. CRC Crit. Rev. Toxicol. 1985, 15, 109–150. [Google Scholar] [CrossRef] [PubMed]
- Moure, A.; Cruz, J.M.; Franco, D.; Domínguez, J.M.; Sineiro, J.; Domínguez, H.; Núñez, M.J.; Parajó, J.C. Natural antioxidants from residual sources. Food Chem. 2001, 72, 145–171. [Google Scholar] [CrossRef]
- Li, X.X.; Han, L.J.; Chen, L.J. In vitro, antioxidant activity of protein hydrolysates prepared from corn gluten meal. J. Sci. Food Agric. 2008, 88, 1660–1666. [Google Scholar] [CrossRef]
- Cheng, Y.H.; Wang, Z.; Xu, S.Y. Antioxidant properties of wheat germ protein hydrolysates evaluated in vitro. J. Cent. South Univ. Technol. 2006, 13, 160–165. [Google Scholar] [CrossRef]
- Li, X.U.; Liu, L.; Li, X.L.; Qiao, Y.M.; Wu, X.X.; Zhang, X.Z. Study on the preparation of corn functional oligopeptides and its SOD-like activity. Chin. J. Biochem. Pharm. 2002, 23, 78–80. [Google Scholar]
- Chen, G.T.; Zhao, L.; Zhao, L.Y.; Cong, T.; Bao, S.F. In vitro study on antioxidant activities of peanut protein hydrolysate. J. Sci. Food Agric. 2007, 87, 357–362. [Google Scholar] [CrossRef]
- PeñtaRamos, E.A.; Xiong, Y.L. Antioxidant Activity of Soy Protein Hydrolysates in a Liposomal System. J. Food Sci. 2002, 67, 2952–2956. [Google Scholar] [CrossRef]
- Wu, H.C.; Chen, H.M.; Shiau, C.Y. Free amino acids and peptides as related to antioxidant properties in protein hydrolysates of mackerel (Scomber austriasicus). Food Res. Int. 2003, 36, 949–957. [Google Scholar] [CrossRef]
- Brunner, H.R.; Laragh, J.H.; Baer, L.; Newton, M.A.; Goodwin, F.T.; Krakoff, L.R.; Bard, R.H.; Bühler, F.R. Essential hypertension: Renin and aldosterone, heart attack and stroke. N. Engl. J. Med. 1972, 286, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.K.; Mendis, E.; Je, J.Y.; Park, P.J.; Son, B.W.; Kim, H.C.; Choi, Y.K.; Kim, S.K. Angiotensin I-converting enzyme inhibitory peptide from yellowfin sole (Limanda aspera) frame protein and its antihypertensive effect in spontaneously hypertensive rats. Food Chem. 2006, 94, 26–32. [Google Scholar] [CrossRef]
- Katayama, K.; Tomatsu, M.; Kawahara, S.; Yamauchi, K.; Fuchu, H.; Kodama, Y.; Kawamura, Y.; Muguruma, M. Inhibitory Profile of Nonapeptide Derived from Porcine Troponin C against Angiotensin I-Converting Enzyme. J. Agric. Food Chem. 2004, 52, 771–775. [Google Scholar] [CrossRef] [PubMed]
- Yust, M.M.; Pedroche, J.; Girón-Calle, J.; Alaiz, M.; Millan, F.; Vioque, J. Production of ace inhibitory peptides by digestion of chickpea legumin with alcalase. Food Chem. 2003, 81, 363–369. [Google Scholar] [CrossRef]
- Kamau, S.M.; Lu, R.R. The Effect of Enzymes and Hydrolysis Conditions on Degree of Hydrolysis and DPPH Radical Scavenging Activity of Whey Protein Hydrolysates. Curr. Res. Dairy Sci. 2011, 3, 25–35. [Google Scholar] [CrossRef]
- Wu, H.T.; Jin, W.G.; Sun, S.G.; Li, X.S.; Duan, X.H.; Li, Y.; Yang, Y.T.; Han, J.R.; Zhu, B.W. Identification of antioxidant peptides from protein hydrolysates of scallop (Patinopecten yessoensis) female gonads. Eur. Food Res. Technol. 2015, 242, 713–722. [Google Scholar] [CrossRef]
- Qin, L.; Zhu, B.W.; Zhou, D.Y.; Wu, H.T.; Tan, H.; Yang, J.F.; Li, D.M.; Dong, X.P.; Murata, Y. Preparation and antioxidant activity of enzymatic hydrolysates from purple sea urchin (Strongylocentrotus nudus) gonad. LWT-Food Sci. Technol. 2011, 44, 1113–1118. [Google Scholar] [CrossRef]
- Feder, J. Studies on the Specificity of Bacillus subtilis Neutral Protease with Synthetic Substrates. Biochem. Biophys. Res. Commun. 1967, 28, 318–323. [Google Scholar] [CrossRef]
- Olsen, J.V.; Ong, S.E.; Mann, M. Trypsin cleaves exclusively C-terminal to arginine and lysine residues. Mol. Cell. Proteom. 2004, 3, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.; Gupta, N.; Smith, R.D.; Pevzneret, P.A. Does Trypsin Cut Before Proline? J. Proteome Res. 2008, 7, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Fruton, J.S. The Specificity and Mechanism of Pepsin Action; Acid Proteases: Structure, Function, and Biology; Springer: New York, NY, USA, 1977; pp. 401–443. [Google Scholar]
- Lee, M.Y.; Choi, Y.C.; Chun, J.Y.; Min, S.G.; Hong, G.P. Effects of High Pressure/High Temperature Processing on the Recovery and Characteristics of Porcine Placenta Hydrolysates. Korean J. Food Sci. Anim. Resour. 2013, 33, 474–480. [Google Scholar] [CrossRef]
- Saiga, A.; Tanabe, S.; Nishimura, T. Antioxidant Activity of Peptides Obtained from Porcine Myofibrillar Proteins by Protease Treatment. J. Agric. Food Chem. 2003, 51, 3661–3667. [Google Scholar] [CrossRef] [PubMed]
- Zainol, M.K.; Abdhamid, A.; Yusof, S.; Muse, R. Antioxidative activity and total phenolic compounds of leaf, root and petiole of four accessions of Centella asiatica (L.) Urban. Food Chem. 2003, 81, 575–581. [Google Scholar] [CrossRef]
- Suetsuna, K.; Ukeda, H.; Ochi, H. Isolation and characterization of free radical scavenging activities peptides derived from casein. J. Nutr. Biochem. 2000, 11, 128–131. [Google Scholar] [CrossRef]
- Chen, H.M.; Muramoto, K.; Yamauchi, F.; Nokihara, K. Antioxidant Activity of Designed Peptides Based on the Antioxidative Peptide Isolated from Digests of a Soybean Protein. J. Agric. Food Chem. 2014, 44, 2619–2623. [Google Scholar] [CrossRef]
- Guo, H.; Yoshiaki, K.; Masami, Y. Structures and properties of antioxidative peptides derived from royal jelly protein. Food Chem. 2009, 113, 238–245. [Google Scholar] [CrossRef]
- Cheng, Z.; Ren, J.; Li, Y.; Chang, W.; Chen, Z. Establishment of a quantitative structure-activity relationship model for evaluating and predicting the protective potentials of phenolic antioxidants on lipid peroxidation. J. Pharm. Sci. 2003, 92, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Pihlanto, A. Antioxidative peptides derived from milk proteins. Int. Dairy J. 2006, 16, 1306–1314. [Google Scholar] [CrossRef]
- Je, J.Y.; Qian, Z.J.; Byun, H.G.; Kim, S.J. Purification and characterization of an antioxidant peptide obtained from tuna backbone protein by enzymatic hydrolysis. Process Biochem. 2007, 42, 840–846. [Google Scholar] [CrossRef]
- Rajapakse, N.; Mendis, E.; Byun, H.G.; Kim, S.K. Purification and in vitro antioxidative effects of giant squid muscle peptides on free radical-mediated oxidative systems. J. Nutr. Biochem. 2005, 16, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Gao, X.D.; Zhou, G.C.; Cai, L.; Yao, W.B. In vitro and in vivo antioxidant activity of aqueous extract from Choerospondias axillaris fruit. Food Chem. 2008, 106, 888–895. [Google Scholar] [CrossRef]
- Jaeyoung, J.; Kahwa, L.; Mihyun, L.; Bum, A.C. Antioxidant and antihypertensive protein hydrolysates produced from tuna liver by enzymatic hydrolysis. Food Res. Int. 2009, 42, 1266–1272. [Google Scholar] [CrossRef]
- Hartmann, R.; Meisel, H. Food-derived peptides with biological activity: From research to food applications. Curr. Opin. Biotechnol. 2007, 18, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Mullally, M.M.; Meisel, H.; Fitzgerald, R.J. Angiotensin-I-coverting enzyme inhibitory activities of gastric and pancreatic proteinase digests of whey proteins. Int. Dairy J. 1997, 7, 299–303. [Google Scholar] [CrossRef]
- Lv, S.W.; Zhou, W.H.; Jiang, Z.M. The research about the hydrolysis process of casein. Sci. Technol. Food Ind. 2003, 20, 24–28. [Google Scholar]
- Lassoued, I.; Mora, L.; Nasri, R.; Jridi, M.; Toldra, F.; Aristoy, M.C.; Barkia, A.; Nasri, M. Characterization and comparative assessment of antioxidant and ACE inhibitory activities of thornback ray gelatin hydrolysates. J. Funct. Foods 2015, 13, 225–238. [Google Scholar] [CrossRef]
- Theodore, A.E.; Kristinsson, H.G. Angiotensin converting enzyme inhibition of fish protein hydrolysates prepared from alkaline-aided channel catfish protein isolate. J. Sci. Food Agric. 2007, 87, 2353–2357. [Google Scholar] [CrossRef]
- Wang, Q.; Wen, Y.; Xia, T.; Xiong, X.; Gao, T.; You, C.; Tao, X.; Zhang, F.; Chen, W. Quantification of 18 amino acids in human plasma: Application in renal transplant patient plasma by targeted UHPLC-MS/MS. Bioanalysis 2016, 8, 1337–1351. [Google Scholar] [CrossRef] [PubMed]
- Kapila, S.; Sinha, P.R. Antioxidative and hypocholesterolemic effect of Lactobacillus casei ssp casei (biodefensive properties of lactobacilli). Indian J. Med. Sci. 2006, 60, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Suda, I.; Masuda, M.; Furuta, S.; Nishiba, Y.; Oki, T.; Kobayashi, M. DPPH radical-scavenging activity of representative antioxidant. Kyushu Agric. Res. 2003, 6, 148–154. [Google Scholar]
- Cushman, D.W.; Cheung, H.S. Spectrophotometric assay and properties of the angiotensin-converting enzyme of rabbit lung. Biochem. Pharmacol. 1971, 20, 1637–1648. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition | Trypsin | Neutrase | Pepsin |
|---|---|---|---|
| Moisture (%) | 11.95 | 14.64 | 14.64 |
| Protein (%) | 55.79 | 53.60 | 50.96 |
| Fat (%) | 2.01 | 2.10 | 3.50 |
| Ash (%) | 18.13 | 14.59 | 19.00 |
| Amino Acid | Trypsin | Neutrase | Pepsin | |||
|---|---|---|---|---|---|---|
| mg/Kg | % | mg/Kg | % | mg/Kg | % | |
| Glycine | 139.62 | 15.56 | 186.96 | 11.52 | 106.73 | 14.43 |
| Alanine | 94.00 | 10.48 | 195.67 | 12.06 | 80.25 | 10.85 |
| Serine | 22.49 | 2.51 | 29.05 | 1.79 | 8.98 | 1.21 |
| Threonine | 27.28 | 3.04 | 67.05 | 4.13 | 16.17 | 2.19 |
| Aspartic acid | 13.43 | 1.50 | 26.19 | 1.61 | 18.69 | 2.53 |
| Glutamic acid | 76.81 | 8.56 | 138.31 | 8.53 | 90.57 | 12.24 |
| Cysteine | 1.80 | 0.20 | 2.32 | 0.14 | 1.84 | 0.25 |
| Proline | 14.68 | 1.64 | 39.07 | 2.41 | 29.25 | 3.95 |
| Lysine | 88.87 | 9.91 | 113.60 | 7.00 | 33.73 | 4.56 |
| Methionine | 9.61 | 1.07 | 26.79 | 1.65 | 10.78 | 1.46 |
| Arginine | 165.16 | 18.41 | 168.36 | 10.38 | 89.31 | 12.07 |
| Valine | 29.30 | 3.27 | 99.22 | 6.12 | 32.51 | 4.39 |
| Glutamine | 22.15 | 2.47 | 72.62 | 4.48 | 19.13 | 2.59 |
| Asparagine | 1.65 | 0.18 | 5.02 | 0.31 | 0.73 | 0.10 |
| Histidine | 4.79 | 0.53 | 24.29 | 1.50 | 4.10 | 0.55 |
| Phenylalanine | 35.51 | 3.96 | 64.04 | 3.95 | 38.65 | 5.22 |
| Tryptophan | 13.32 | 1.48 | 27.10 | 1.67 | 13.02 | 1.76 |
| Tyrosine | 37.66 | 4.20 | 59.57 | 3.67 | 31.62 | 4.27 |
| Leucine | 62.41 | 6.96 | 182.86 | 11.27 | 82.07 | 11.09 |
| Isoleucine | 36.71 | 4.09 | 94.29 | 5.81 | 31.68 | 4.28 |
| Total hydrophobic amino acids | 295.54 | 32.94 | 729.03 | 44.94 | 318.22 | 43.01 |
| Total essential amino acids | 331.07 | 36.90 | 707.71 | 43.62 | 279.47 | 37.77 |
| Total | 897.26 | 100.00 | 1622.38 | 100.00 | 739.83 | 100.00 |
| Enzyme | Optimum Conditions | Time (h) | E:S | DH (%) | |
|---|---|---|---|---|---|
| pH | Temperature (°C) | ||||
| Trypsin | 9.0 | 45 | 4 | 3/100 | 17.54% |
| Neutrase | 6.5 | 45 | 4 | 3/100 | 18.30% |
| Pepsin | 4.0 | 45 | 4 | 3/100 | 8.34% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Y.; Fan, F.; Wu, D.; Yu, C.; Wang, Z.; Du, M. Antioxidant and ACE Inhibitory Activity of Enzymatic Hydrolysates from Ruditapes philippinarum. Molecules 2018, 23, 1189. https://doi.org/10.3390/molecules23051189
Yu Y, Fan F, Wu D, Yu C, Wang Z, Du M. Antioxidant and ACE Inhibitory Activity of Enzymatic Hydrolysates from Ruditapes philippinarum. Molecules. 2018; 23(5):1189. https://doi.org/10.3390/molecules23051189
Chicago/Turabian StyleYu, Yue, Fengjiao Fan, Di Wu, Cuiping Yu, Zhenyu Wang, and Ming Du. 2018. "Antioxidant and ACE Inhibitory Activity of Enzymatic Hydrolysates from Ruditapes philippinarum" Molecules 23, no. 5: 1189. https://doi.org/10.3390/molecules23051189
APA StyleYu, Y., Fan, F., Wu, D., Yu, C., Wang, Z., & Du, M. (2018). Antioxidant and ACE Inhibitory Activity of Enzymatic Hydrolysates from Ruditapes philippinarum. Molecules, 23(5), 1189. https://doi.org/10.3390/molecules23051189