Enzymatic Synthesis of a Novel Pterostilbene α-Glucoside by the Combination of Cyclodextrin Glucanotransferase and Amyloglucosidase

 ,
,  , , ,
, , ,  , ,
, , 
Abstract
:
1. Introduction
2. Results and Discussion
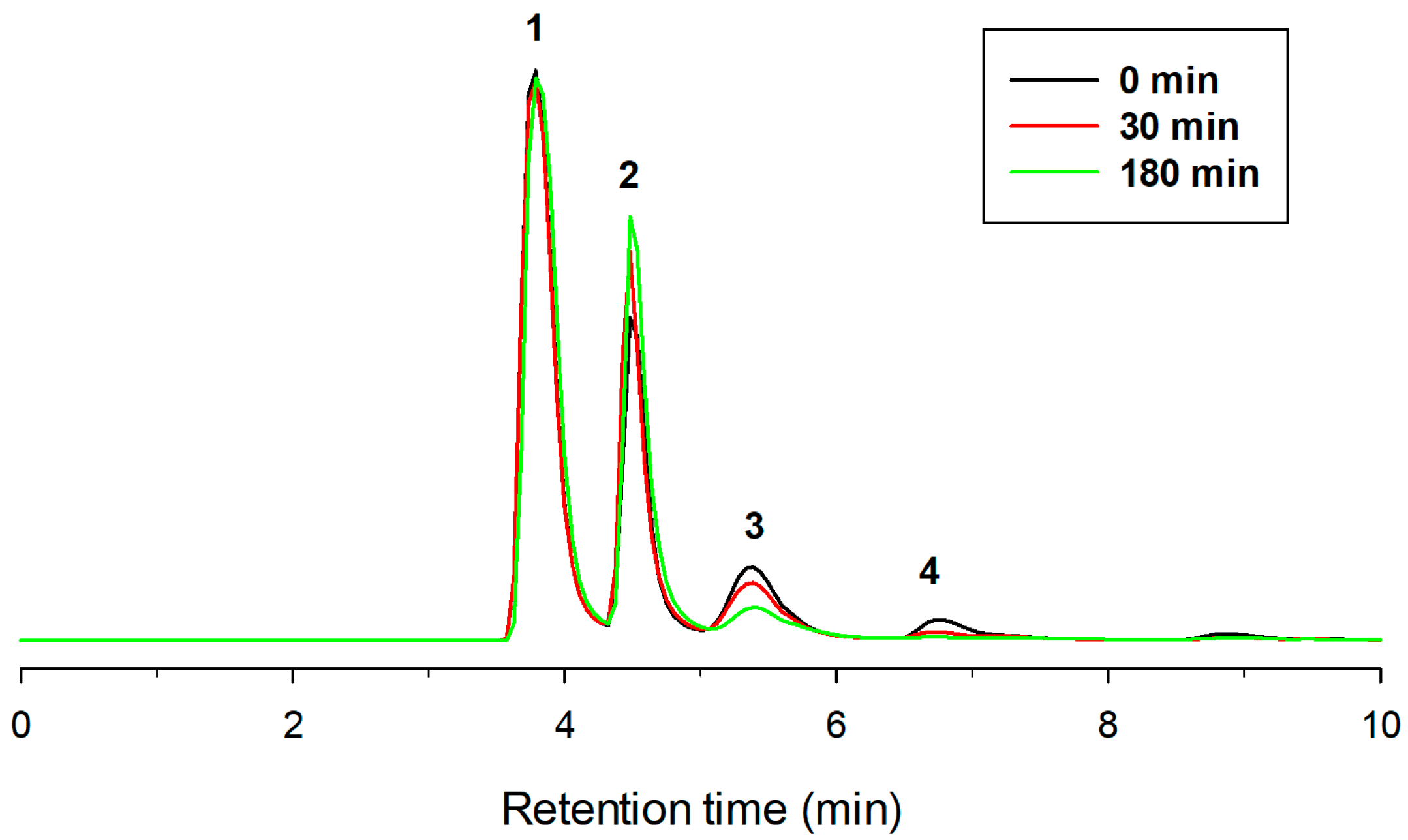
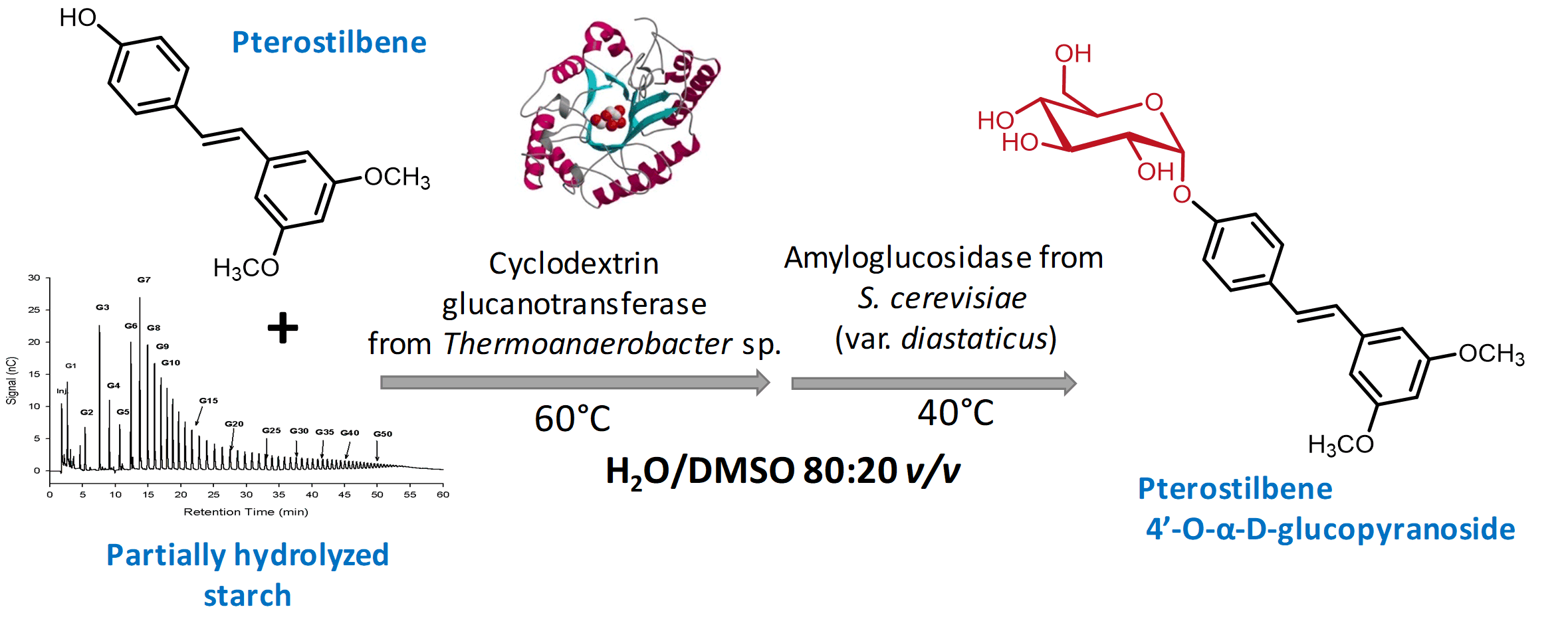
2.1. Enzymatic Glucosylation of Pterostilbene
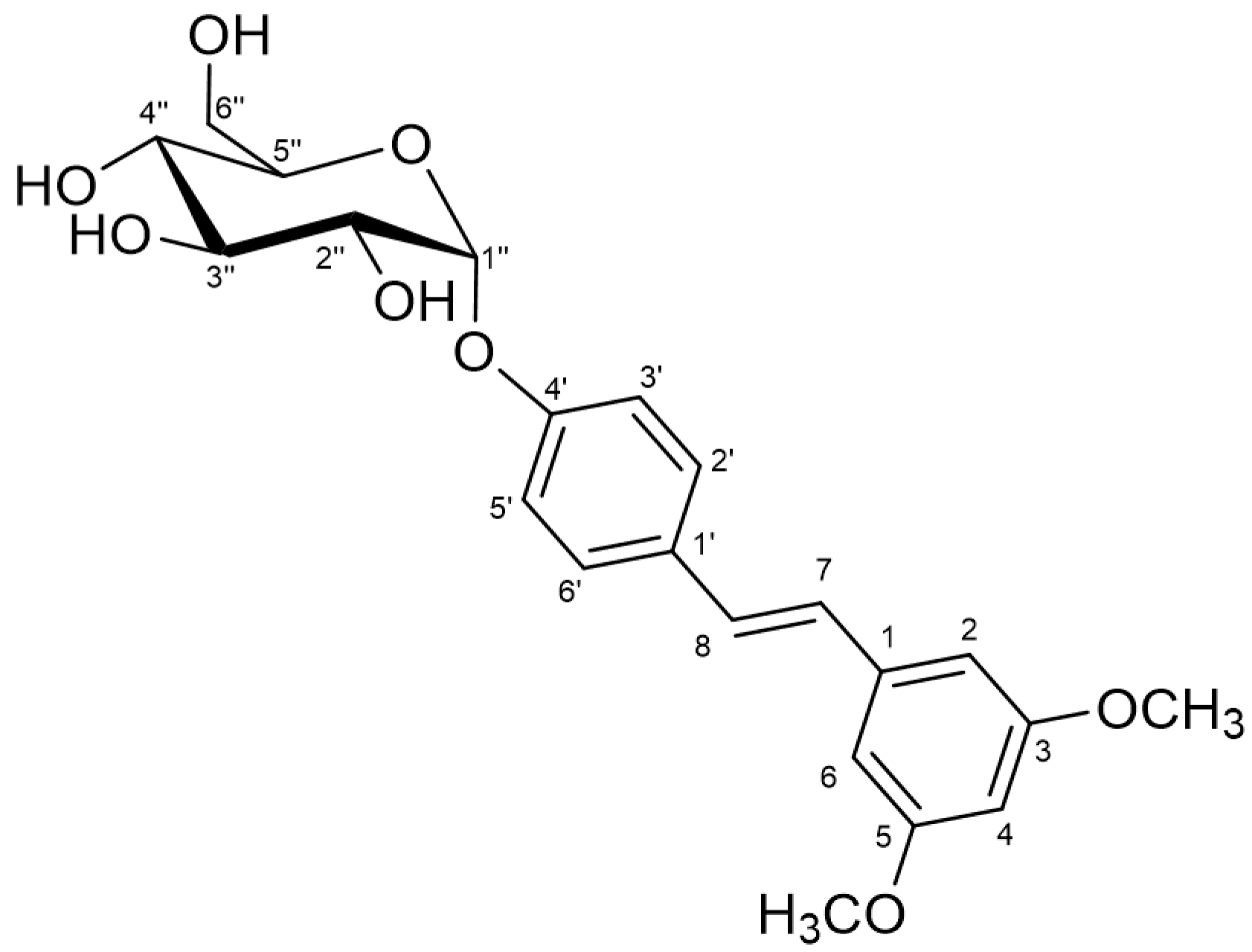
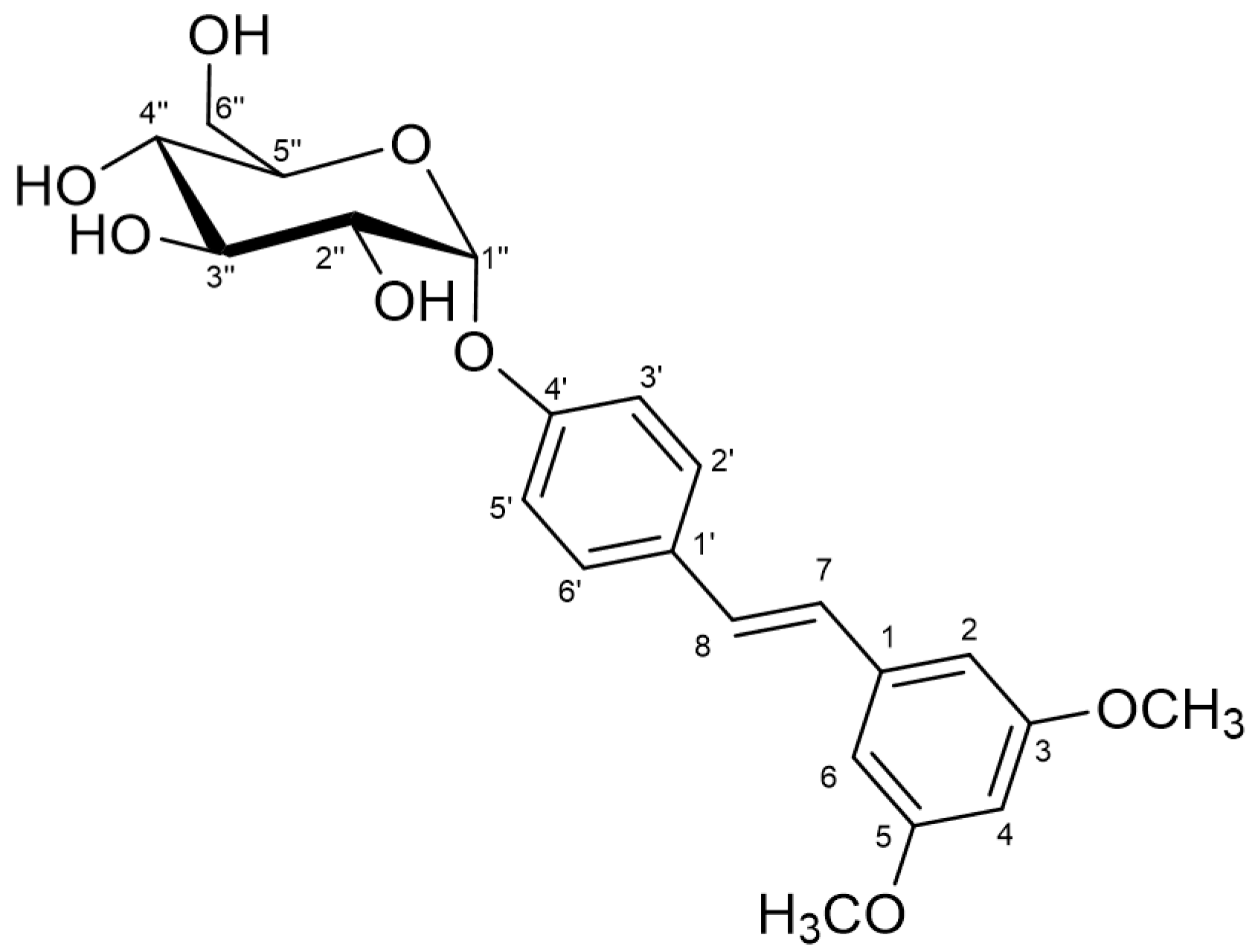
2.2. Characterization of the Monoglucosylated Derivative
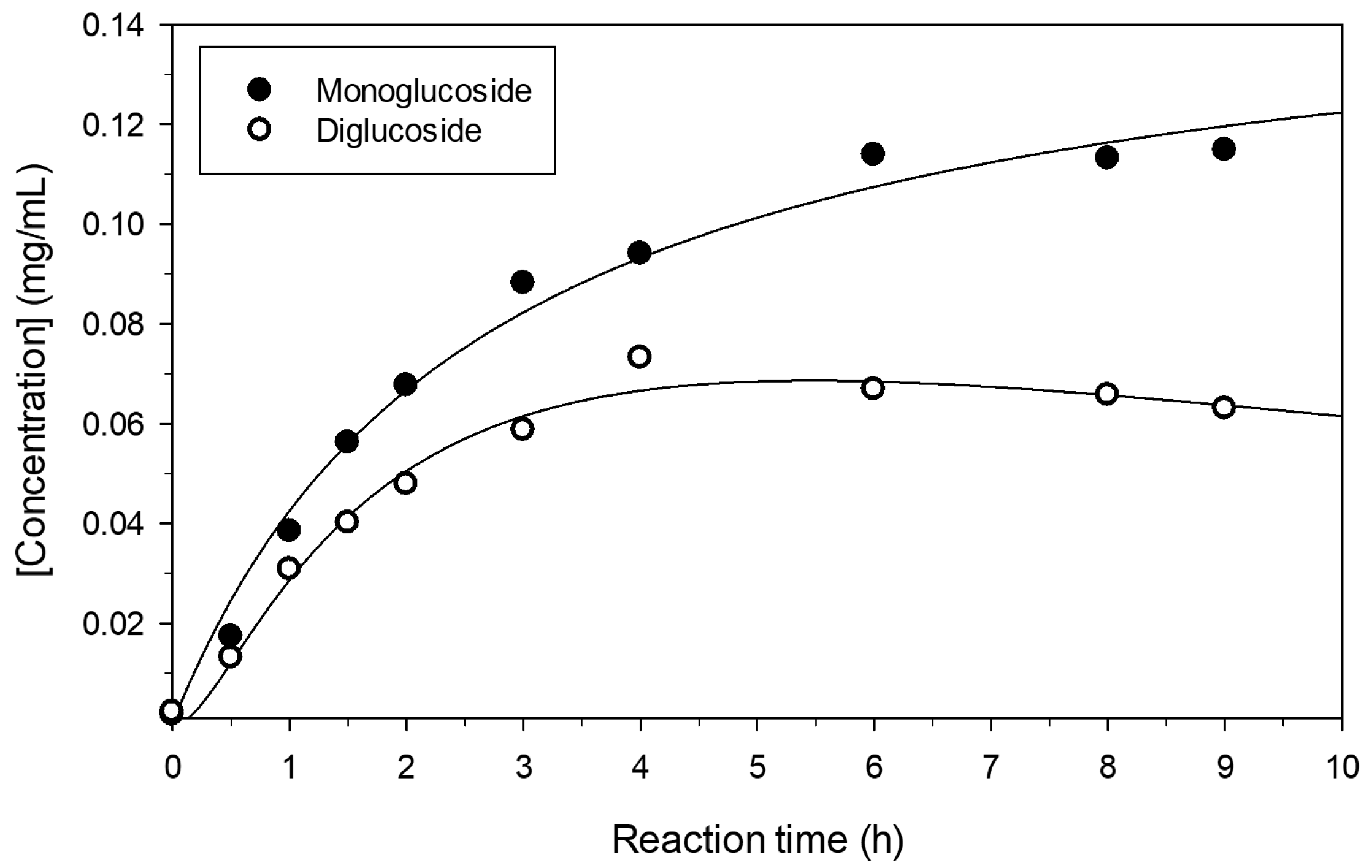
2.3. Increase of Monoglucoside Yield by Treatment with Amyloglucosidase
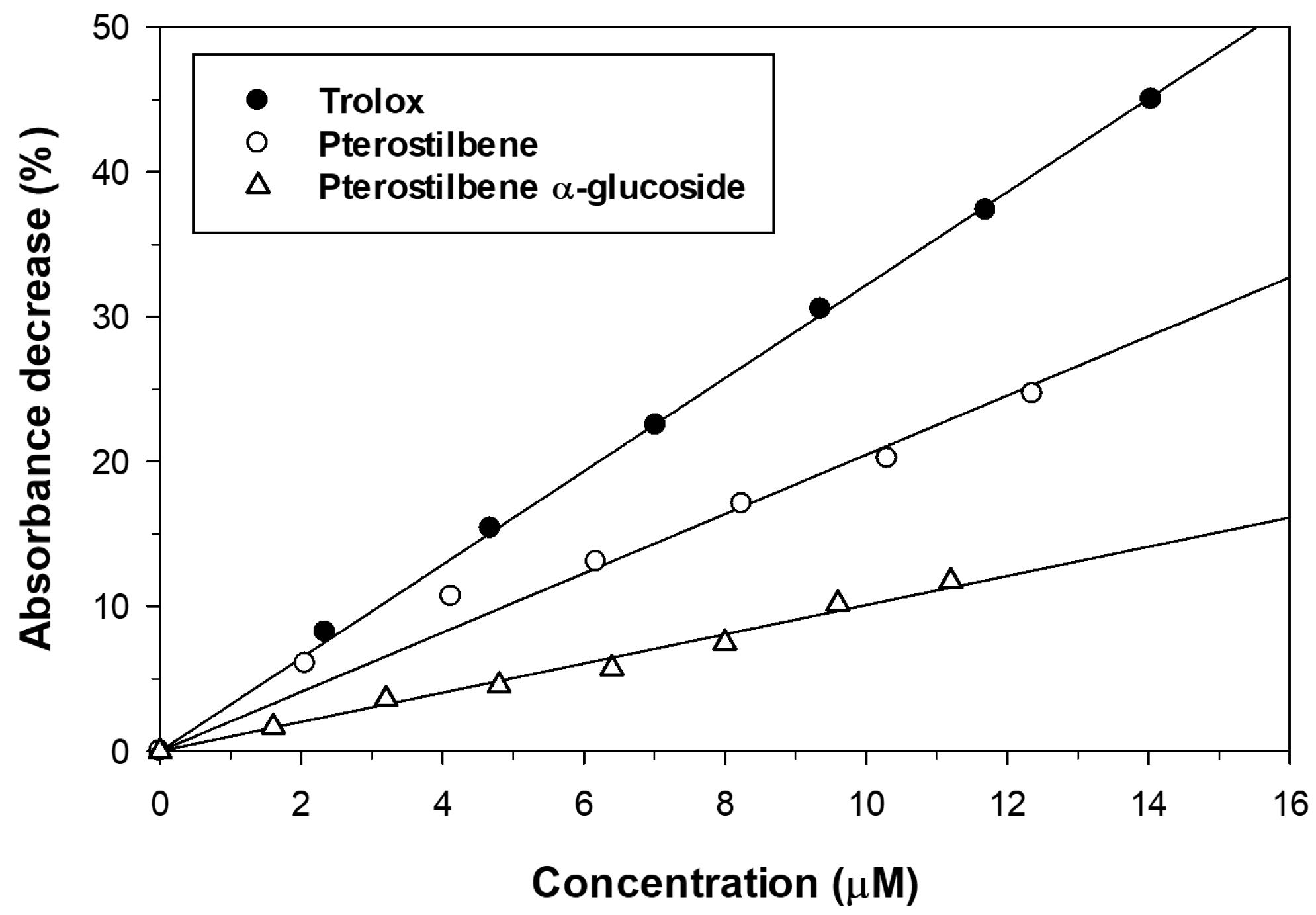
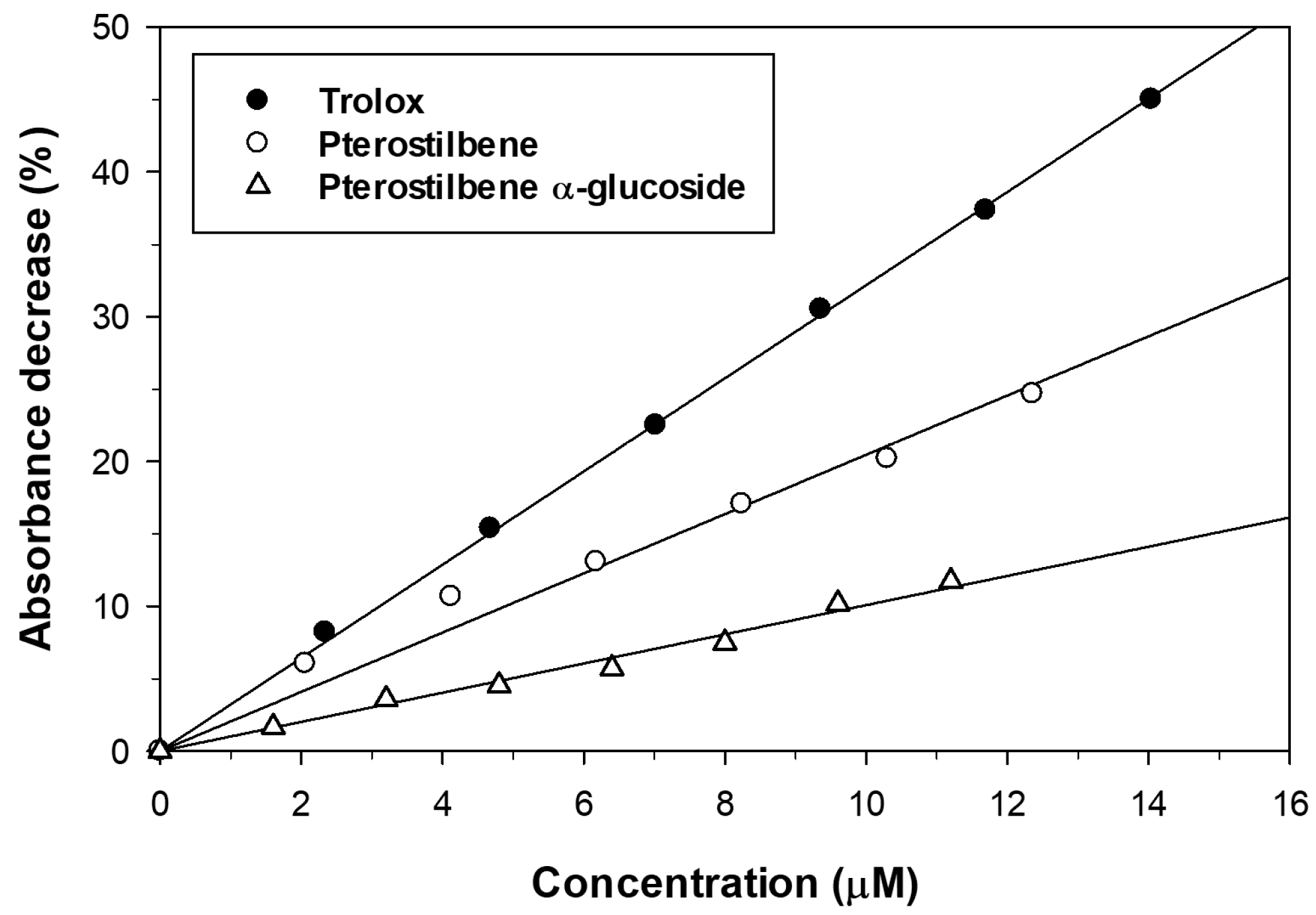
2.4. Effect of Glucosylation on Solubility and Antioxidant Activity of Pterostilbene
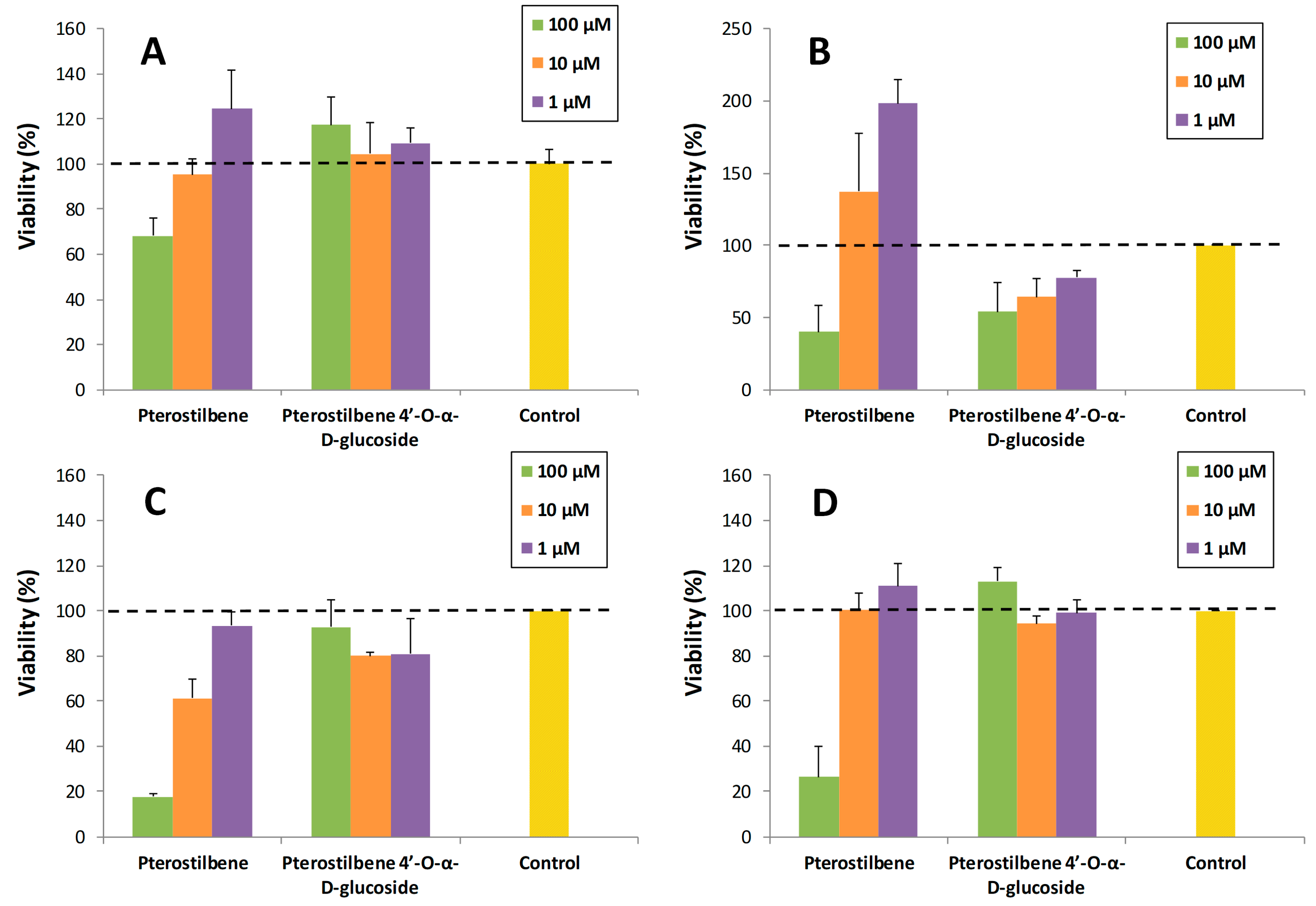
2.5. Effect of Glucosylation on Toxicity of Pterostilbene
3. Materials and Methods
3.1. Chemicals
3.2. Enzymes
3.3. Enzymatic Glucosylation of Pterostilbene
3.4. High-Performance Liquid Chromatography (HPLC)
3.5. Purification of Pterostilbene Glucosides
3.6. Mass Spectrometry (MS)
3.7. Nuclear Magnetic Resonance (NMR)
3.8. Trolox Equivalent Antioxidant Capacity (TEAC) Assay
3.9. Solubility Assay
3.10. Toxicity Tests
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gonzalez-Sarrias, A.; Nuñez-Sanchez, M.A.; Tomas-Barberan, F.A.; Espin, J.C. Neuroprotective effects of bioavailable polyphenol-derived metabolites against oxidative stress-induced cytotoxicity in human neuroblastoma SH-SY5Y cells. J. Agric. Food Chem. 2017, 65, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Peng, R.M.; Lin, G.R.; Ting, Y.; Hu, J.Y. Oral delivery system enhanced the bioavailability of stilbenes: Resveratrol and pterostilbene. BioFactors 2018, 44, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Sang, S. Metabolism and pharmacokinetics of resveratrol and pterostilbene. BioFactors 2018, 44, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.J.; Kuo, H.C.; Cheng, L.H.; Lee, Y.H.; Chang, W.T.; Wang, B.J.; Wang, Y.J.; Cheng, H.C. Apoptotic and nonapoptotic activities of pterostilbene against cancer. Int. J. Mol. Sci. 2018, 19, 287. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, B.; Ammazzalorso, A.; Fantacuzzi, M.; Giampietro, L.; Maccallini, C.; Amoroso, R. Anticancer activity of stilbene-based derivatives. ChemMedChem 2017, 12, 558–570. [Google Scholar] [CrossRef] [PubMed]
- Seo, E.J.; Fischer, N.; Efferth, T. Phytochemicals as inhibitors of NF-κB for treatment of Alzheimer’s disease. Pharm. Res. 2018, 129, 262–273. [Google Scholar] [CrossRef] [PubMed]
- Lange, K.W.; Li, S. Resveratrol, pterostilbene, and dementia. BioFactors 2018, 44, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.H.; Wu, J.C.; Ho, C.T.; Lai, C.S. Antiobesity molecular mechanisms of action: Resveratrol and pterostilbene. BioFactors 2018, 44, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Kosuru, R.; Singh, S. Pterostilbene ameliorates insulin sensitivity, glycemic control and oxidative stress in fructose-fed diabetic rats. Life Sci. 2017, 182, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.R.; Li, S.; Lin, C.C. Effect of resveratrol and pterostilbene on aging and longevity. BioFactors 2018, 44, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Vivekanandhan, D.K.; Verma, P.R.P.; Singh, S.K. Emerging technologies for improving bioavailability of polyphenols. Curr. Nutr. Food Sci. 2016, 12, 12–22. [Google Scholar] [CrossRef]
- Vrba, J.; Kren, V.; Vacek, J.; Papouskova, B.; Ulrichova, J. Quercetin, quercetin glycosides and taxifolin differ in their ability to induce AhR activation and CYP1A1 expression in HepG2 cells. Phytother. Res. 2012, 26, 1746–1752. [Google Scholar] [CrossRef] [PubMed]
- Makino, T.; Shimizu, R.; Kanemaru, M.; Suzuki, Y.; Moriwaki, M.; Mizukami, H. Enzymatically modified isoquercitrin, α-oligoglucosyl quercetin 3-O-glucoside, is absorbed more easily than other quercetin glycosides or aglycone after oral administration in rats. Biol. Pharm. Bull. 2009, 32, 2034–2040. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Domínguez, M.; de Eugenio, L.I.; Peñalver, P.; Belmonte-Reche, E.; Morales, J.C.; Poveda, A.; Jiménez-Barbero, J.; Prieto, A.; Plou, F.J.; Martínez, M.J. Enzymatic synthesis of a novel neuroprotective hydroxytyrosyl glycoside. J. Agric. Food Chem. 2017, 65, 10526–10533. [Google Scholar] [CrossRef] [PubMed]
- Lepak, A.; Gutmann, A.; Kulmer, S.T.; Nidetzky, B. Creating a water-soluble resveratrol-based antioxidant by site-selective enzymatic glucosylation. ChemBioChem 2015, 16, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Medina, I.; Alcantara, D.; Gonzalez, M.J.; Torres, P.; Lucas, R.; Roque, J.; Plou, F.J.; Morales, J.C. Antioxidant activity of resveratrol in several fish lipid matrices: Effect of acylation and glucosylation. J. Agric. Food Chem. 2010, 58, 9778–9786. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Qi, T.; Xu, L.; Lu, L.; Xiao, M. Recent progress in the enzymatic glycosylation of phenolic compounds. J. Carbohydr. Chem. 2016, 35, 1–23. [Google Scholar] [CrossRef]
- Nadim, M.; Auriol, D.; Lamerant-FayeL, N.; Lefe, F.; Dubanet, L.; Redziniak, G.; Kieda, C.; Grillon, C. Improvement of polyphenol properties upon glucosylation in a UV-induced skin cell ageing model. Int. J. Cosmet. Sci. 2014, 36, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Desmet, T.; Soetaert, W.; Bojarova, P.; Kren, V.; Dijkhuizen, L.; Eastwick-Field, V.; Schiller, A. Enzymatic glycosylation of small molecules: Challenging substrates require tailored catalysts. Chem. Eur. J. 2012, 18, 10786–10801. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Soto, M.E.; Seibel, J. Biotechnological synthesis and transformation of valuable sugars in the food and pharmaceutical industry. Curr. Org. Chem. 2014, 18, 964–986. [Google Scholar] [CrossRef]
- Plou, F.J.; Gómez de Segura, A.; Ballesteros, A. Application of glycosidases and transglycosidases for the synthesis of oligosaccharides. In Industrial Enzymes: Structure, Function and Application; Polaina, J., MacCabe, A.P., Eds.; Springer: New York, NY, USA, 2007; pp. 141–157. [Google Scholar]
- Planas, A.; Faijes, M. Glycosidases and glycosynthases in enzymatic synthesis of oligosaccharides. An overview. Afinidad 2002, 59, 295–313. [Google Scholar]
- Fernandez-Arrojo, L.; Marin, D.; de Segura, A.G.; Linde, D.; Alcalde, M.; Gutierrez-Alonso, P.; Ghazi, I.; Plou, F.J.; Fernandez-Lobato, M.; Ballesteros, A. Transformation of maltose into prebiotic isomaltooligosaccharides by a novel alpha-glucosidase from Xantophyllomyces dendrorhous. Process Biochem. 2007, 42, 1530–1536. [Google Scholar] [CrossRef]
- Bojarova, P.; Kren, V. Glycosidases: A key to tailored carbohydrates. Trends Biotechnol. 2009, 27, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Alcalde, M.; Plou, F.J.; Andersen, C.; Martin, M.T.; Pedersen, S.; Ballesteros, A. Chemical modification of lysine side chains of cyclodextrin glycosyltransferase from Thermoanaerobacter causes a shift from cyclodextrin glycosyltransferase to alpha-amylase specificity. FEBS Lett. 1999, 445, 333–337. [Google Scholar] [CrossRef]
- Torres, P.; Poveda, A.; Jimenez-Barbero, J.; Parra, J.L.; Comelles, F.; Ballesteros, A.O.; Plou, F.J. Enzymatic synthesis of α-glucosides of resveratrol with surfactant activity. Adv. Synth. Catal. 2011, 353, 1077–1086. [Google Scholar] [CrossRef]
- Marié, T.; Willig, G.; Teixeira, A.R.S.; Gazaneo Barboza, E.; Kotland, A.; Gratia, A.; Courot, E.; Hubert, J.; Renault, J.H.; Allais, F. Enzymatic synthesis of resveratrol α-glycosides from β-cyclodextrin-resveratrol complex in water. ACS Sustain. Chem. Eng. 2018, 6, 5370–5380. [Google Scholar] [CrossRef]
- Funayama, M.; Nishino, T.; Hirota, A.; Murao, S.; Takenishi, S.; Nakano, H. Enzymatic synthesis of (+)catechin-α-glucoside and its effect on tyrosinase activity. Biosci. Biotech. Biochem. 1993, 57, 1666–1669. [Google Scholar] [CrossRef]
- Mathew, S.; Adlercreutz, P. Regioselective glycosylation of hydroquinone to α-arbutin by cyclodextrin glucanotransferase from Thermoanaerobacter sp. Biochem. Eng. J. 2013, 79, 187–193. [Google Scholar] [CrossRef]
- Choung, W.J.; Hwang, S.H.; Ko, D.S.; Kim, S.B.; Kim, S.H.; Jeon, S.H.; Choi, H.D.; Lim, S.S.; Shim, J.H. Enzymatic synthesis of a novel kaempferol-3-O-β-d-glucopyranosyl- (1→4)-O-α-d-glucopyranoside using cyclodextrin glucanotransferase and its inhibitory effects on aldose reductase, inflammation, and oxidative stress. J. Agric. Food Chem. 2017, 65, 2760–2767. [Google Scholar] [CrossRef] [PubMed]
- Han, R.; Ge, B.; Jiang, M.; Xu, G.; Dong, J.; Ni, Y. High production of genistein diglucoside derivative using cyclodextrin glycosyltransferase from Paenibacillus macerans. J. Ind. Microbiol. Biot. 2017, 44, 1343–1354. [Google Scholar] [CrossRef] [PubMed]
- Ioffe, I.N.; Granovsky, A.A. Photoisomerization of stilbene: The detailed XMCQDPT2 treatment. J. Chem. Theory Comput. 2013, 9, 4973–4990. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.T.; Cruces, M.A.; Alcalde, M.; Plou, F.J.; Bernabe, M.; Ballesteros, A. Synthesis of maltooligosyl fructofuranosides catalyzed by immobilized cyclodextrin glucosyltransferase using starch as donor. Tetrahedron 2004, 60, 529–534. [Google Scholar] [CrossRef]
- Plou, F.J.; Martin, M.T.; Gomez de Segura, A.; Alcalde, M.; Ballesteros, A. Glucosyltransferases acting on starch or sucrose for the synthesis of oligosaccharides. Can. J. Chem. 2002, 80, 743–752. [Google Scholar] [CrossRef]
- Mizuno, C.S.; Schrader, K.K.; Rimando, A.M. Algicidal activity of stilbene analogues. J. Agric. Food Chem. 2008, 56, 9140–9145. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, K.; Kubota, N.; Uesugi, D.; Hamada, H.; Tanigawa, M.; Hamada, H. Synthesis and pharmacological evaluation of glycosides of resveratrol, pterostilbene, and piceatannol. Ann. N.Y. Acad. Sci. 2015, 1348, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Uesugi, D.; Hamada, H.; Shimoda, K.; Kubota, N.; Ozaki, S.I.; Nagatani, N. Synthesis, oxygen radical absorbance capacity, and tyrosinase inhibitory activity of glycosides of resveratrol, pterostilbene, and pinostilbene. Biosci. Biotech. Biochem. 2017, 81, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Chen, Y.; Han, M.; Sun, L.; Wang, Q. Synthesis and biological activity of pterostilbene and 3′-methoxy pterostilbene and their glucosides derivatives. Chin. J. Org. Chem. 2011, 31, 2114–2120. [Google Scholar]
- Monthieu, C.; Guibert, A.; Taravel, F.R.; Nardin, R.; Combes, D. Purification and characterisation of polyglucosyl-fructosides produced by means of cyclodextrin glucosyl transferase. Biocatal. Biotransfor. 2003, 21, 7–15. [Google Scholar] [CrossRef]
- Kloosterman, W.M.J.; Dijk, G.S.V.; Loos, K. Biocatalytic synthesis of maltodextrin-based acrylates from starch and a-cyclodextrina. Macromol. Biosci. 2014, 14, 1268–1279. [Google Scholar] [CrossRef] [PubMed]
- Tonkova, A. Bacterial cyclodextrin glucanotransferase. Enzyme Microb. Tech. 1998, 22, 678–686. [Google Scholar] [CrossRef]
- Miranda-Molina, A.; Marquina-Bahena, S.; Lopez-Munguía, A.; Alvarez, L.; Castillo, E. Regioselective glucosylation of inositols catalyzed by Thermoanaerobacter sp. CGTase. Carbohydr. Res. 2012, 360, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Gudiminchi, R.K.; Towns, A.; Varalwar, S.; Nidetzky, B. Enhanced synthesis of 2-O-α-d-glucopyranosyl-L-ascorbic acid from α-cyclodextrin by a highly disproportionating CGTase. ACS Catal. 2016, 6, 1606–1615. [Google Scholar] [CrossRef]
- Latorre-García, L.; Adam, A.C.; Polaina, J. Overexpression of the glucoamylase-encoding STA1 gene of Saccharomyces cerevisiae var. diastaticus in laboratory and industrial strains of Saccharomyces. World J. Microb. Biot. 2008, 24, 2957–2963. [Google Scholar]
- Lee, Y.S.; Woo, J.B.; Ryu, S.I.; Moon, S.K.; Han, N.S.; Lee, S.B. Glucosylation of flavonol and flavanones by Bacillus cyclodextrin glucosyltransferase to enhance their solubility and stability. Food Chem. 2017, 229, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Yang, Y.; Di, S.; Feng, X.; Liu, D.; Jiang, S.; Hu, W.; Qin, Z.; Li, Y.; Lv, J.; et al. Pterostilbene exerts anticancer activity on non-small-cell lung cancer via activating endoplasmic reticulum stress. Sci. Rep. 2017, 7, 8091. [Google Scholar] [CrossRef] [PubMed]
- He, J.L.; Dong, X.H.; Li, Z.H.; Wang, X.Y.; Fu, Z.A.; Shen, N. Pterostilbene inhibits reactive oxygen species production and apoptosis in primary spinal cord neurons by activating autophagy via the mechanistic target of rapamycin signaling pathway. Mol. Med. Rep. 2018, 17, 4406–4414. [Google Scholar] [CrossRef] [PubMed]
- Sato, D.; Shimizu, N.; Shimizu, Y.; Akagi, M.; Eshita, Y.; Ozaki, S.I.; Nakajima, N.; Ishihara, K.; Masuoka, N.; Hamada, H.; et al. Synthesis of glycosides of resveratrol, pterostilbene, and piceatannol, and their anti-oxidant, anti-allergic, and neuroprotective activities. Biosci. Biotech. Biochem. 2015, 78, 1123–1128. [Google Scholar] [CrossRef] [PubMed]
- Hamada, H.; Shimoda, K.; Horio, Y.; Ono, T.; Hosoda, R.; Nakayama, N.; Urano, K. Pterostilbene and its glucoside induce type XVII collagen expression. Nat. Prod. Commun. 2017, 12, 85–86. [Google Scholar]
- Falomir, E.; Lucas, R.; Peñalver, P.; Marti-Centelles, R.; Dupont, A.; Zafra-Gómez, A.; Carda, M.; Morales, J.C. Cytotoxic, antiangiogenic and antitelomerase activity of glucosyl- and acyl- resveratrol prodrugs and resveratrol sulfate metabolites. ChemBioChem 2016, 17, 1343–1348. [Google Scholar] [CrossRef] [PubMed]
- Correia-da-Silva, M.; Rocha, V.; Marques, C.; Deus, C.; Marques-Carvalho, A.; Oliveira, P.; Palmeira, A.; Pinto, M.; Sousa, M.E.; Sousa Lobo, J.; et al. Potential benefits of a sulfated resveratrol derivative for topical application. J. Mol. Endocrinol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Kuete, V.; Sandjo, L.P.; Mbaveng, A.T.; Seukep, J.A.; Ngadjui, B.T.; Efferth, T. Cytotoxicity of selected Cameroonian medicinal plants and Nauclea pobeguinii towards multi-factorial drug-resistant cancer cells. BMC Complem. Altern. Med. 2015, 15, 309. [Google Scholar] [CrossRef] [PubMed]
- Su, D.; Cheng, Y.; Liu, M.; Liu, D.; Cui, H.; Zhang, B.; Zhou, S.; Yang, T.; Mei, Q. Comparision of piceid and resveratrol in antioxidation and antiproliferation activities in vitro. PLoS ONE 2013, 8, e54505. [Google Scholar] [CrossRef] [PubMed]
- Gimeno-Perez, M.; Linde, D.; Fernandez-Arrojo, L.; Plou, F.J.; Fernandez-Lobato, M. Heterologous overproduction of beta-fructofuranosidase from yeast Xanthophyllomyces dendrorhous, an enzyme producing prebiotic sugars. Appl. Microbiol. Biotechnol. 2015, 99, 3459–3467. [Google Scholar] [CrossRef] [PubMed]
- Adam, A.C.; Latorre-García, L.; Polaina, J. Structural analysis of glucoamylase encoded by the STA1 gene of Saccharomyces cerevisiae (var. diastaticus). Yeast 2004, 21, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Letchmanan, K.; Shen, S.C.; Ng, W.K.; Tan, R.B.H. Application of transglycosylated stevia and hesperidin as drug carriers to enhance biopharmaceutical properties of poorly-soluble artemisinin. Colloid. Surf. B 2018, 161, 83–93. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Source | Glycosyl Donor | [Monoglucoside] (mg/mL) a |
|---|---|---|---|
| β-galactosidase | Bacillus circulans | Lactose | 0.0015 |
| α-glucosidase | Aspergillus niger | Maltose | 0.0002 |
| β-fructofuranosidase | Saccharomyces cerevisiae | Sucrose | − |
| Xanthophyllomyces dendrorhous | Sucrose | − | |
| CGTase | Thermoanaerobacter sp. | Starch | 0.1148 |
| Bacillus macerans | Starch | 0.0129 |
| Position | δC, Type a | δH (J in Hz) |
|---|---|---|
| 2/6 | 103.98, CH | 6.75 (d, J = 2.2 Hz, 2H) |
| 4 | 99.28, CH | 6.40 (t, J = 2.2 Hz, 1H) |
| OMe (3/5) | 54.90, CH3 | 3.78 (s, 6H) |
| 7 | 128.15, CH | 7.22 (d, J = 16.3 Hz, 1H) |
| 8 | 126.40, CH | 7.05 (d, J = 16.3 Hz, 1H) |
| 2′/6′ | 127.39, CH | 7.53 (d, J = 8.7 Hz, 2H) |
| 3′/5′ | 116.85, CH | 7.10 (d, J = 8.7 Hz, 2H) |
| 1′′ | 97.62, CH | 5.41 (d, J = 3.6 Hz, 1H) |
| 2′′ | 71.29, CH | 3.38 (ddd, J = 9.6, 6.3, 3.6 Hz, 1H) |
| 3′′ | 72.84, CH | 3.63 (td, J = 9.6, 5.0 Hz, 1H) |
| 4′′ | 69.65, CH | 3.20 (td, J = 9.6, 5.8 Hz, 1H) |
| 5′′ | 73.49, CH | 3.44–3.51 (m, 1H)b |
| 6′′ | 60.45, CH2 | 3.44–3.51 (m, 1H)b/3.57 (dd, J = 10.0, 5.6 Hz, 1H) |
| OH2′′ | - | 5.05 (d, J = 6.3 Hz, 1H) |
| OH3′′ | - | 4.93 (d, J = 5.0 Hz, 1H) |
| OH4′′ | - | 4.97 (d, J = 5.8 Hz, 1H) |
| OH6′′ | - | 4.47 (t, J = 5.7 Hz, 1H) |
| Compound | Slope | R2 | TEAC |
|---|---|---|---|
| Trolox | 3.22 | 0.996 | 1.00 |
| Pterostilbene | 1.74 | 0.990 | 2.40 |
| Pterostilbene 4′-O-α-d-glucoside | 1.03 | 0.980 | 4.05 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Alfonso, J.L.; Rodrigo-Frutos, D.; Belmonte-Reche, E.; Peñalver, P.; Poveda, A.; Jiménez-Barbero, J.; Ballesteros, A.O.; Hirose, Y.; Polaina, J.; Morales, J.C.; et al. Enzymatic Synthesis of a Novel Pterostilbene α-Glucoside by the Combination of Cyclodextrin Glucanotransferase and Amyloglucosidase. Molecules 2018, 23, 1271. https://doi.org/10.3390/molecules23061271
González-Alfonso JL, Rodrigo-Frutos D, Belmonte-Reche E, Peñalver P, Poveda A, Jiménez-Barbero J, Ballesteros AO, Hirose Y, Polaina J, Morales JC, et al. Enzymatic Synthesis of a Novel Pterostilbene α-Glucoside by the Combination of Cyclodextrin Glucanotransferase and Amyloglucosidase. Molecules. 2018; 23(6):1271. https://doi.org/10.3390/molecules23061271
Chicago/Turabian StyleGonzález-Alfonso, José L., David Rodrigo-Frutos, Efres Belmonte-Reche, Pablo Peñalver, Ana Poveda, Jesús Jiménez-Barbero, Antonio O. Ballesteros, Yoshihiko Hirose, Julio Polaina, Juan C. Morales, and et al. 2018. "Enzymatic Synthesis of a Novel Pterostilbene α-Glucoside by the Combination of Cyclodextrin Glucanotransferase and Amyloglucosidase" Molecules 23, no. 6: 1271. https://doi.org/10.3390/molecules23061271