Antifeedant Effects of Essential Oil, Extracts, and Isolated Sesquiterpenes from Pilgerodendron uviferum (D. Don) Florin Heartwood on Red Clover Borer Hylastinus obscurus (Coleoptera: Curculionidae)

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
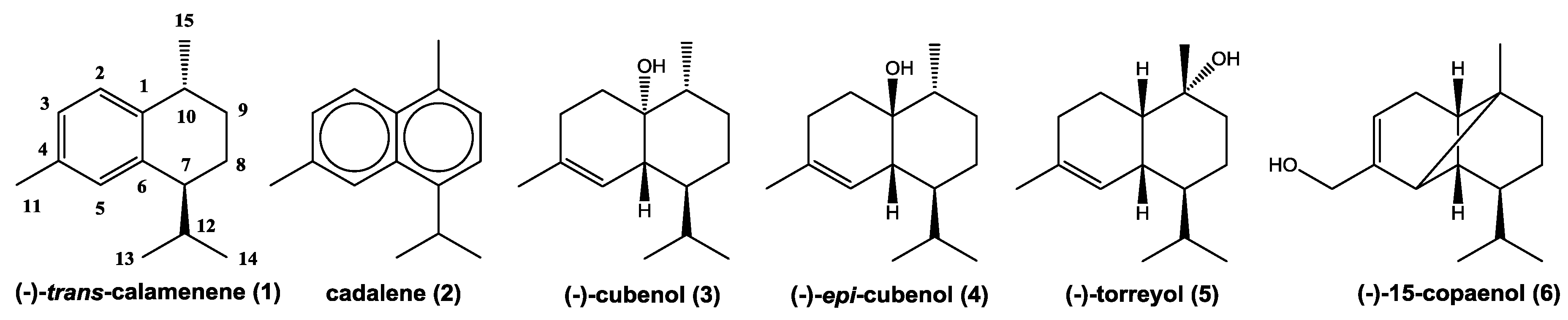
2.1. Chemical Analyses
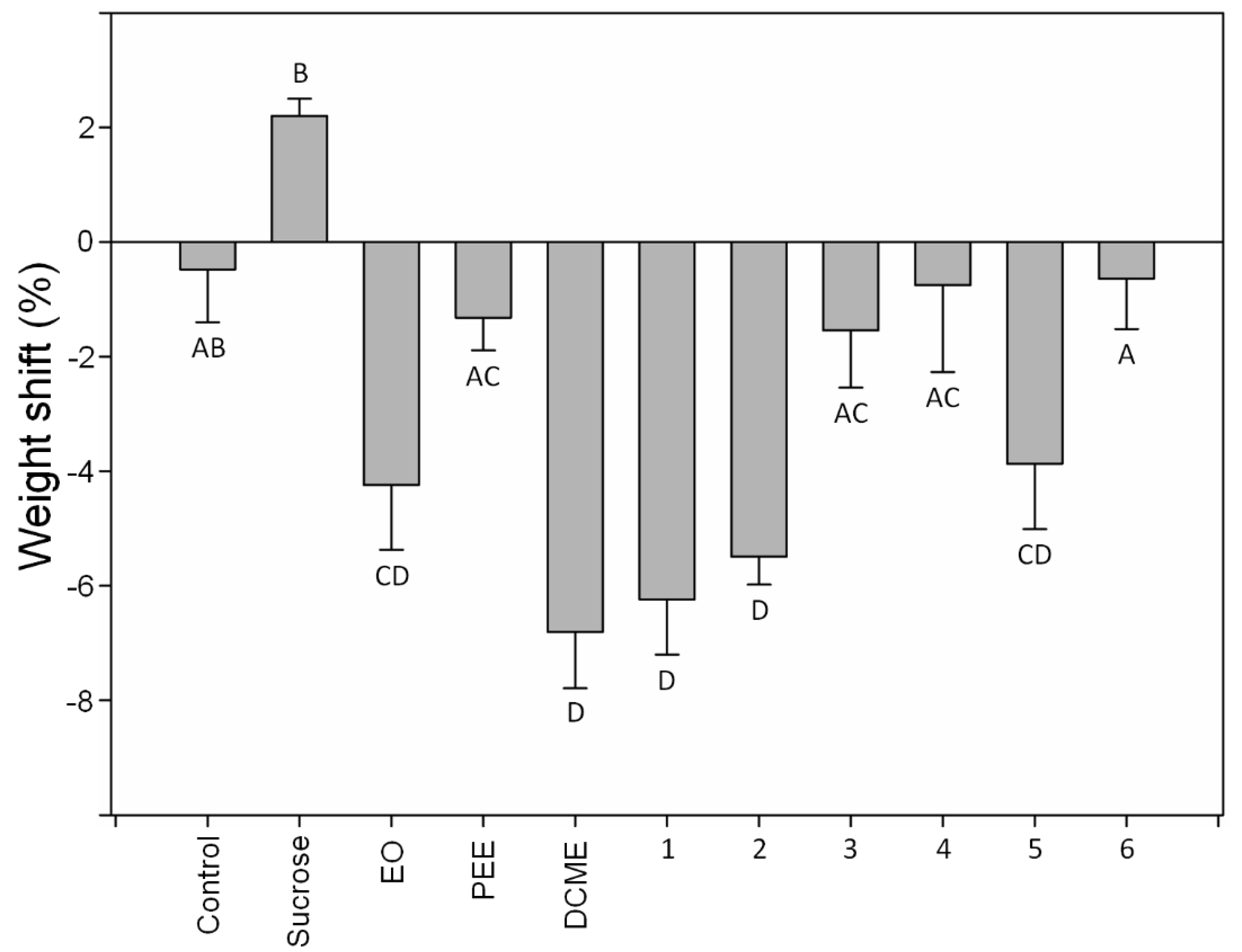
2.2. Antifeedant Bioassays
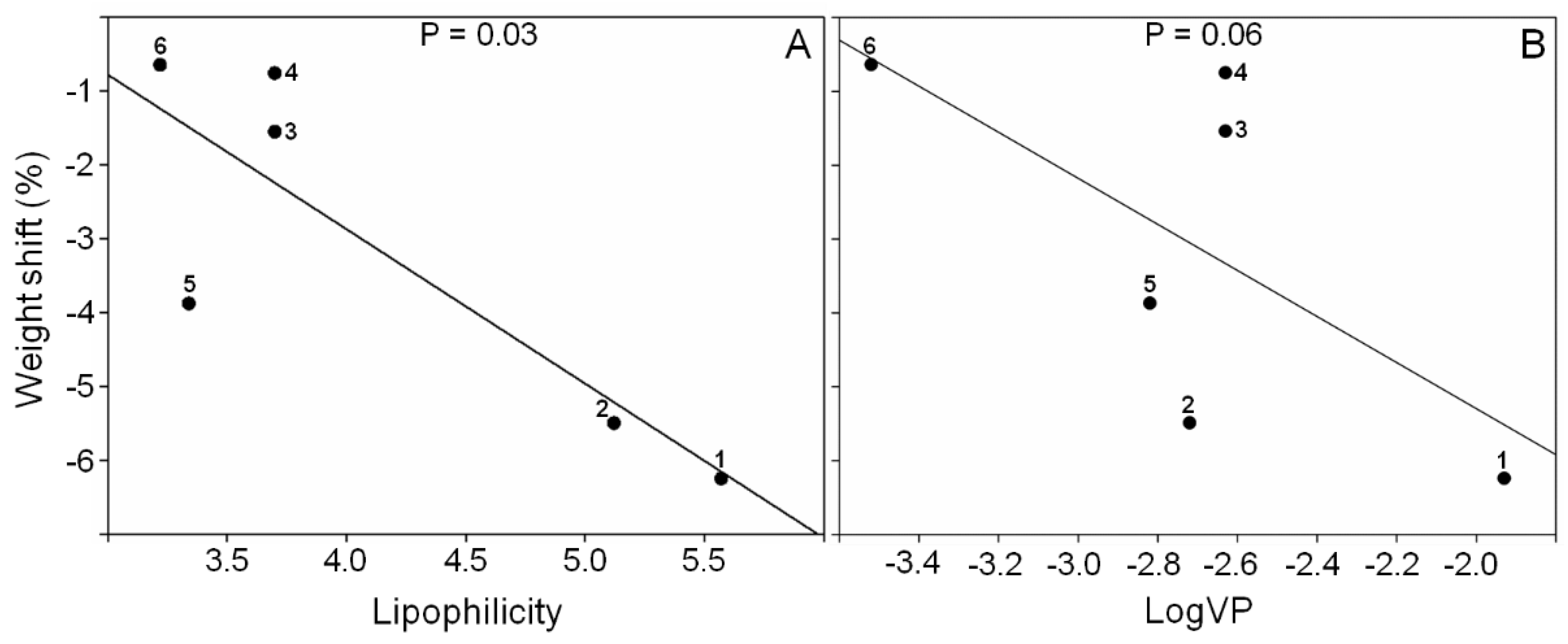
3. Discussion
4. Material and Methods
4.1. Plant and Insect Material
4.2. Heartwood EO, PEE, and DCME Extraction
4.3. Sesquiterpene Isolation
4.4. Analyses of Heartwood EO, Extracts, and Sesquiterpenes
4.5. Theoretical Estimation of Lipophilicity and Vapor Pressure
4.6. Feeding Bioassays
4.7. Statistical Data Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Taylor, N.L.; Quesenberry, K.H. Red Clover Science; Springer Science & Business Media: Dordrecht, The Netherlands, 1996. [Google Scholar]
- Fergus, E.N.; Hollowell, E.A. Red clover. Adv. Agron. 1960, 12, 365–436. [Google Scholar]
- Steiner, J.J.; Alderman, S.C. Red Clover Seed Production. VI. Effect and economic of soil pH adjusted by lime application. Crop Sci. 2003, 43, 624–630. [Google Scholar] [CrossRef]
- Ortega, F. Variation in Mortality, Yield and Persistence of Red Clover (Trifolium pratense L.). Ph.D. Thesis, University of Wales, Aberystwyth, UK, 1996. [Google Scholar]
- Manosalva, L.; Pardo, F.; Perich, F.; Mutis, A.; Parra, L.; Ortega, F.; Isaacs, R.; Quiroz, A. Behavioral responses of clover root borer to long-chain fatty acids from young red clover (Trifolium pratense) roots. Environ. Entomol. 2011, 40, 399–404. [Google Scholar] [CrossRef]
- Ferrero, A.A.; Chopa, C.S.; González, J.O.W.; Alzogaray, R.A. Repellence and toxicity of Schinus molle extracts on Blattella germanica. Fitoterapia 2007, 78, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Palacios, S.M.; Bertoni, A.; Rossi, Y.; Santander, R.; Urzúa, A. Efficacy of essential oils from edible plants as insecticides against the house fly, Musca domestica L. Molecules 2009, 14, 1938–1947. [Google Scholar] [CrossRef] [PubMed]
- Cantrell, C.L.; Dayan, F.E.; Duke, S.O. Natural products as sources for new pesticides. J. Nat. Prod. 2012, 75, 1231–1242. [Google Scholar] [CrossRef] [PubMed]
- Karamaouna, F.; Kimbaris, A.; Michaelakis, A.; Papachristos, D.; Polissiou, M.; Papatsakona, P.; Tsora, E.; Miller, T. Insecticidal activity of plant essential oils against the vine mealybug, Planococcus ficus. J. Insect. Sci. 2013, 13, 142. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-H.; Luo, S.-H.; Li, S.-H.; Gao, J.-M. New Antifeedant grayanane diterpenoids from the flowers of Pieris formosa. Molecules 2017, 22, 1431. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, E.; Balocchi, O. Producción de Forraje; Producción Animal; Universidad Austral de Chile: Valdivia, Chile, 1983. [Google Scholar]
- Parra, L.; Mutis, A.; Ortega, F.; Quiroz, A. Field response of Hylastinus obscurus Marsham (Coleoptera: Curculionidae) to E-2-hexenal and limonene, two host-derived semiochemicals. Cienc. Investig. Agrar. 2013, 40, 637–642. [Google Scholar] [CrossRef]
- Aguilera, A.; Cisternas, E.; Gerding, M.; Norambuena, H.; Ruiz, I. Plagas de las praderas. In Praderas Para Chile; Ruiz, I., Ed.; Instituto de Investigaciones Agropecuarias (INIA): Santiago, Chile, 1996; Volume 2, pp. 309–340. [Google Scholar]
- Quiroz, A.; Ortega, F.; Ramírez, C.C.; Wadhams, L.J.; Pinilla, K. Response of the beetle Hylastinus obscurus Marsham (Coleoptera: Scolytidae) to red clover (Trifolium pratense L.) volatiles in a laboratory olfactometer. Environ. Entomol. 2005, 34, 690–695. [Google Scholar] [CrossRef]
- Quiroz, A.; Mendez, L.; Mutis, A.; Hormazabal, E.; Ortega, F.; Birkett, M.A.; Parra, L. Antifeedant activity of red clover root isoflavonoids on Hylastinus obscurus. J. Soil Sci. Plant Nutr. 2017, 17, 231–239. [Google Scholar] [CrossRef]
- Solís, C.; Becerra, J.; Flores, C.; Robledo, J.; Silva, M. Antibacterial and antifungal terpenes from Pilgerodendron uviferum (D. Don) Florin. J. Chil. Chem. Soc. 2004, 49, 157–161. [Google Scholar] [CrossRef]
- Donoso, C.A.; Becerra, J.; Bittner, M.; Elissetche, J.P.; Freer, J.; Mendoza, R.; Sterner, O.; Silva, M. Allelochemicals and natural durability in Chilean Cupressaceae heartwoods. Allelopath. J. 2008, 21, 119–132. [Google Scholar]
- Espinoza, J.; Urzúa, A.; Tampe, J.; Parra, L.; Quiroz, A. Repellent activity of the essential oil from the heartwood of Pilgerodendron uviferum (D. Don) Florin against Aegorhinus superciliosus (Coleoptera: Curculionidae). Molecules 2016, 21, 533. [Google Scholar] [CrossRef] [PubMed]
- Oyarzún, M.L.; Garbarino, J.A. Sesquiterpenoids from Pilgerodendron uvífera. Phytochemistry 1988, 27, 1121–1123. [Google Scholar] [CrossRef]
- Croft, K.D.; Ghisalberti, E.L.; Hocart, C.H.; Jefferies, P.R.; Raston, C.L.; White, A.H. Absolute configuration of the (+)-calamenenes: Crystal structure of 7-hydroxycalamenene. J. Chem. Soc. Perkin Trans. 1 1978, 10, 1267–1270. [Google Scholar] [CrossRef]
- Bunko, J.D.; Ghisalberti, E.L.; Jefferies, P.R. (1R,4S)-Calamenene. Chemical correlation with the diterpene analogues of Eremophila. Aust. J. Chem. 1981, 34, 2237–2242. [Google Scholar] [CrossRef]
- Kuo, Y.-H.; Yu, M.-T. Four new sesquiterpenes from the heartwood of Juniperus formosana var. concolor. Chem. Pharm. Bull. 1999, 47, 1017–1019. [Google Scholar] [CrossRef]
- Nagasampagi, B.A.; Yankov, L.; Dev, S. Sesquiterpenoids from the wood of Cedrela toona Roxb; partial synthesis of t-muurolol, t-cadinol and cubenol; structures of δ-cadinene and δ-cadinol. Tetrahedron Lett. 1968, 9, 1913–1918. [Google Scholar] [CrossRef]
- Connolly, J.D.; Phillips, W.R.; Huneck, S. (+)-Ent-epicubenol from the liverwort Scapania undulata. Phytochemistry 1982, 21, 233–234. [Google Scholar] [CrossRef]
- Westfelt, L. (−)-Torreyol (“δ-cadinol”). Acta Chem. Scand. 1970, 24, 1618–1622. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy, 4th ed.; Stream, C., Ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Conti, B.; Canale, A.; Cioni, P.L.; Flamini, G. Repellence of essential oils from tropical and Mediterranean Lamiaceae against Sitophilus zeamais. Bull. Insectol. 2010, 63, 197–202. [Google Scholar]
- Mossi, A.J.; Astolfi, V.; Kubiak, G.; Lerin, L.; Zanella, C.; Toniazzo, G.; Oliveira, D.; deTreichel, H.; Devilla, I.A.; Cansian, R. Insecticidal and repellency activity of essential oil of Eucalyptus sp. against Sitophilus zeamais Motschulsky (Coleoptera, Curculionidae). J. Sci. Food Agric. 2011, 91, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Kordali, S.; Aslan, I.; Çalmaşur, O.; Cakir, A. Toxicity of essential oils isolated from three Artemisia species and some of their major components to granary weevil, Sitophilus granarius (L.) (Coleoptera: Curculionidae). Ind. Crops Prod. 2006, 23, 162–170. [Google Scholar] [CrossRef]
- Shukla, P.; Vidyasagar, P.; Aldosari, S.A.; Abdel-Azim, M. Antifeedant activity of three essential oils against the red palm weevil, Rhynchophorus ferrugineus. Bull. Insectology 2012, 65, 71–76. [Google Scholar]
- Ortiz de Elguea-Culebras, G.; Sánchez-Vioque, R.; Berruga, M.I.; Herraiz-Peñalver, D.; Santana-Méridas, O. Antifeedant effects of common terpenes from Mediterranean aromatic plants on Leptinotarsa decemlineata. J. Soil Sci. Plant Nutr. 2017, 17, 475–485. [Google Scholar] [CrossRef]
- Toledo, D.; Parra, L.; Mutis, A.; Ortega, F.; Hormazábal, E.; Quiroz, A. Influence of long-chain fatty acids on weight gain of Hylastinus obscurus (Coleoptera: Curculionidae). Cienc. Investig. Agrar. 2014, 41, 357–364. [Google Scholar] [CrossRef]
- Cheng, T.; Zhao, Y.; Li, X.; Lin, F.; Xu, Y.; Zhang, X.; Li, Y.; Wang, R.; Lai, L. Computation of octanol-water partition coefficients by guiding an additive model with knowledge. J. Chem. Inf. Model. 2007, 47, 2140–2148. [Google Scholar] [CrossRef] [PubMed]
- Epa, U. Estimation Programs Interface SuiteTM for Microsoft® Windows, v 4.11; United States Environmental Protection Agency: Washington, DC, USA, 2012.
- Faccoli, M.; Schlyter, F. Conifer phenolic resistance markers are bark beetle antifeedant semiochemicals. Agric. For. Entomol. 2007, 9, 237–245. [Google Scholar] [CrossRef]
Sample Availability: Samples of compounds 1–6 are available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| RT | RIC | RIL | Compounds | PEE % | DCME % | EO * % | Identification |
|---|---|---|---|---|---|---|---|
| 20.16 | 1327 | 1343 | α-cubebene | 0.09 | - | 0.05 | RI, MS |
| 20.82 | 1353 | 1359 | copaene | 0.75 | 0.55 | 0.71 | RI, MS |
| 21.13 | 1364 | 1387 | β-cubebene | 0.07 | - | - | RI, MS |
| 22.66 | 1424 | 1428 | α-caryophyllene | 0.08 | 0.15 | 1.27 | RI, MS |
| 23.13 | 1443 | 1445 | bicyclosesquiphellandrene | - | 0.46 | - | RI, MS |
| 23.2 | 1446 | 1462 | γ-muurolene | 0.33 | - | - | RI, MS |
| 23.77 | 1468 | 1469 | α-muurolene | 0.85 | 0.63 | - | RI, MS |
| 24.17 | 1484 | 1509 | (−)-trans-calamenene | 7.06 | 5.26 | - | RI, MS,NMR |
| 24.28 | 1488 | 1515 | δ-cadinene | 2.75 | 6.43 | 10.8 | RI, MS |
| 24.5 | 1497 | 1524 | 4,5,9,10-dehydroisolongifolene | 0.27 | 2.14 | - | RI, MS |
| 24.6 | 1501 | 1522 | α-calacorene | 0.72 | 5.12 | 0.95 | RI, MS |
| 25.5 | 1540 | 1549 | caryophyllene oxide | 3.12 | 0.48 | 1.02 | RI, MS |
| 25.67 | 1547 | 1553 | germacrene B | 3.00 | 2.6 | - | RI, MS |
| 26.08 | 1565 | 1588 | 1,2-epoxyhumulene | 2.06 | 0.34 | 0.51 | RI, MS |
| 26.65 | 1588 | 1620 | (−)-torreyol | 18.1 | 18.53 | 24.16 | RI, MS,NMR |
| 26.98 | 1602 | 1627 | (−)-epi-cubenol | 10.14 | 8.01 | 12.44 | RI, MS,NMR |
| 26.98 | 1602 | 1628 | (−)-cubenol | 7.0 | 8.1 | 10.5 | RI, MS,NMR |
| 27.17 | 1611 | - | (−)-15-copaenol | 14.25 | 12.5 | 15.46 | MS,NMR |
| 27.33 | 1618 | - | cadala-1(10),3,8-triene | 2.03 | 1.57 | - | MS |
| 27.52 | 1627 | 1652 | cadalene | 0.86 | 3.66 | - | RI, MS,NMR |
| 35.04 | 1991 | - | 7-isopropyl-1,1,4a-trimethyl-1,2,3,4,4a,9,10,10a-octahydrophenanthrene | 0.19 | - | - | MS |
| 39.55 | 2243 | - | 6,7-dehydroferruginol | 3.76 | 1.06 | - | MS |
| 39.63 | 2248 | - | ferruginol | 4.28 | 1.13 | - | MS |
| 42.92 | 2450 | - | hinokione | 0.26 | 0.25 | - | MS |
| 43.53 | 2489 | - | hinokiol | 0.29 | 0.75 | - | MS |
| 44.14 | 2529 | - | sugiol | 0.12 | 0.12 | - | MS |
| Carbon Number a | 13C Chemical Shift (ppm) | |||||
|---|---|---|---|---|---|---|
| trans-Calamenene (1) | Cadalene (2) | epi-Cubenol (3) | Cubenol (4) | 15-Copaenol (5) | Torreyol (6) | |
| 1 | 140.5 | 131.5 | 71.1 | 73.1 | 37.6 | 45.9 |
| 2 | 127.2 | 125.2 | 32.5 | 22.4 | 30.3 | 35.7 |
| 3 | 126.6 | 127.6 | 27.2 | 27.1 | 118.3 | 31.5 |
| 4 | 134.9 | 135.2 | 135.6 | 134.3 | 147.6 | 134.8 |
| 5 | 129.2 | 123.4 | 120.2 | 122.5 | 50.7 | 125.0 |
| 6 | 140.3 | 131.9 | 46.4 | 48.5 | 44.8 | 37.2 |
| 7 | 44.2 | 142.5 | 40.2 | 49.6 | 44.9 | 44.2 |
| 8 | 21.9 | 121.8 | 24.5 | 24.4 | 22.1 | 21.9 |
| 9 | 31.2 | 126.0 | 30.6 | 31.6 | 36.4 | 18.9 |
| 10 | 32.9 | 132.3 | 40.2 | 42.3 | 39.7 | 73.0 |
| 11 | 21.7 | 22.5 | 24.1 | 24.0 | 66.4 | 24.0 |
| 12 | 32.3 | 28.6 | 26.3 | 27.3 | 32.5 | 26.8 |
| 13 | 17.8 | 24.1 | 15.3 | 15.6 | 20.0 | 15.7 |
| 14 | 21.6 | 24.1 | 21.8 | 22.1 | 20.0 | 22.1 |
| 15 | 22.7 | 19.9 | 15.5 | 15.6 | 20.4 | 28.4 |
| Hydrogen Number a | 1H Chemical Shift (ppm) | |||||
|---|---|---|---|---|---|---|
| trans-Calamenene (1) | Cadalene (2) | epi-Cubenol (3) | Cubenol (4) | 15-Copaenol (5) | Torreyol (6) | |
| 1 | - | - | - | - | 2.12 m (1) | 1.62 bs (1) |
| 2 | 7.14 d, J = 8 (1) | 7.95 d, J = 8 (1) | 2.02/1.33 m (1) | 1.66/1.60 m (1) | 2.26 bs (2) | 1.52 m (2) |
| 3 | 6.97 d, J = 8 (1) | 7.37 d, J = 8 (1) | 2.13/1.98 m (1) | 2.08 m (2) | 5.48 m (1) | 1.99 m (2) |
| 4 | - | - | - | - | - | - |
| 5 | 7.04 s (1) | 7.94 s (1) | 5.41 s (1) | 5.44 d, J = 4 (1) | 1.72 d, J = 1 (1) | 5.52 dd (1) |
| 6 | - | - | 1.85 m (1) | 1.67 m (1) | 1.59 b m (1) | 2.02 m (1) |
| 7 | 2.71 m (1) | - | 1.39 m (1) | 1.14 m (1) | 1.69 b m (1) | 1.32 m (1) |
| 8 | 1.84/1.61 m (1) | 7.30 d, J = 7 (1) | 1.63/1.04 m (1) | 1.55/1.04 m (1) | 1.53/1.59 m (1) | 1.49/1.10 m (1) |
| 9 | 1.98/1.34 m (1) | 7.24 d, J = 7 (1) | 1.49 m (2) | 1.60/1.08 m (1) | 1.73/1.63 m (1) | 1.88/1.58 bs (1) |
| 10 | 2.79 m (1) | - | 1.35 m (1) | 1.60 m (1) | - | - |
| 11 | 2.31 s (3) | 2.58 s (3) | 1.71 s (3) | 1.70 s (3) | 3.98 d, J = 1 (2) | 1.65 s (3) |
| 12 | 2.26 m (1) | 3.74 m (1) | 2.07 m (1) | 1.96 m (1) | 1.51 m (1) | 1.97 m (1) |
| 13 b | 0.73 d, J = 7 (3) | 1.41 d, J = 7 (3) | 0.73 d, J = 7 (3) | 0.80 d, J = 7 (3) | 0.82 d, J = 7 (3) | 0.81 d, J = 7 (3) |
| 14 b | 1.02 d, J = 7 (3) | 0.92 d, J = 7 (3) | 0.87 d, J = 7 (3) | 0.84 d, J = 7 (3) | 0.89 d, J = 7 (3) | |
| 15 | 1.28 d, J = 7 (3) | 2.67 s (3) | 0.93 d, J = 7 (3) | 0.96 d, J = 7 (3) | 0.78 s (3) | 1.29 s (3) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Espinoza, J.; Urzúa, A.; Bardehle, L.; Quiroz, A.; Echeverría, J.; González-Teuber, M. Antifeedant Effects of Essential Oil, Extracts, and Isolated Sesquiterpenes from Pilgerodendron uviferum (D. Don) Florin Heartwood on Red Clover Borer Hylastinus obscurus (Coleoptera: Curculionidae). Molecules 2018, 23, 1282. https://doi.org/10.3390/molecules23061282
Espinoza J, Urzúa A, Bardehle L, Quiroz A, Echeverría J, González-Teuber M. Antifeedant Effects of Essential Oil, Extracts, and Isolated Sesquiterpenes from Pilgerodendron uviferum (D. Don) Florin Heartwood on Red Clover Borer Hylastinus obscurus (Coleoptera: Curculionidae). Molecules. 2018; 23(6):1282. https://doi.org/10.3390/molecules23061282
Chicago/Turabian StyleEspinoza, Javier, Alejandro Urzúa, Leonardo Bardehle, Andrés Quiroz, Javier Echeverría, and Marcia González-Teuber. 2018. "Antifeedant Effects of Essential Oil, Extracts, and Isolated Sesquiterpenes from Pilgerodendron uviferum (D. Don) Florin Heartwood on Red Clover Borer Hylastinus obscurus (Coleoptera: Curculionidae)" Molecules 23, no. 6: 1282. https://doi.org/10.3390/molecules23061282