
The Growth Proliferation, Apoptotic Prevention, and Differentiation Induction of the Gelatin Hydrolysates from Three Sources to Human Fetal Osteoblasts (hFOB 1.19 Cells)


Abstract
:1. Introduction
2. Results
2.1. Growth Proliferation of Gelatin Hydrolysates for the Osteoblasts
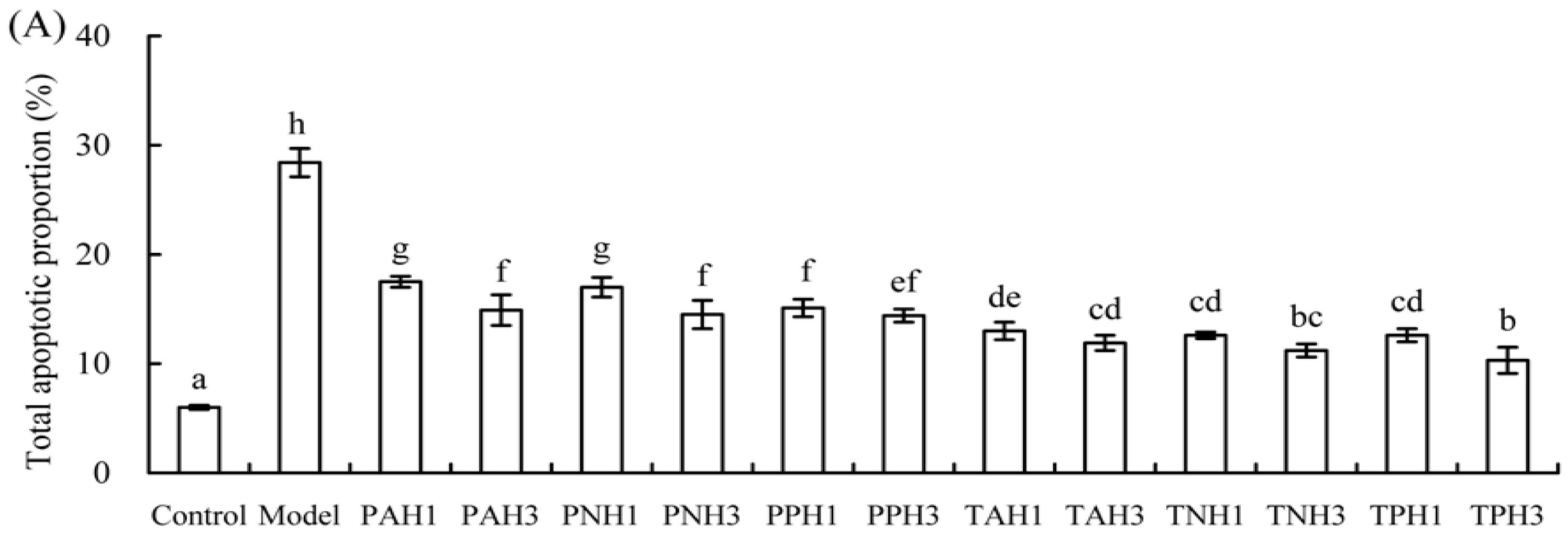
2.2. Apoptotic Prevention of Gelatin Hydrolysates on the Osteoblasts
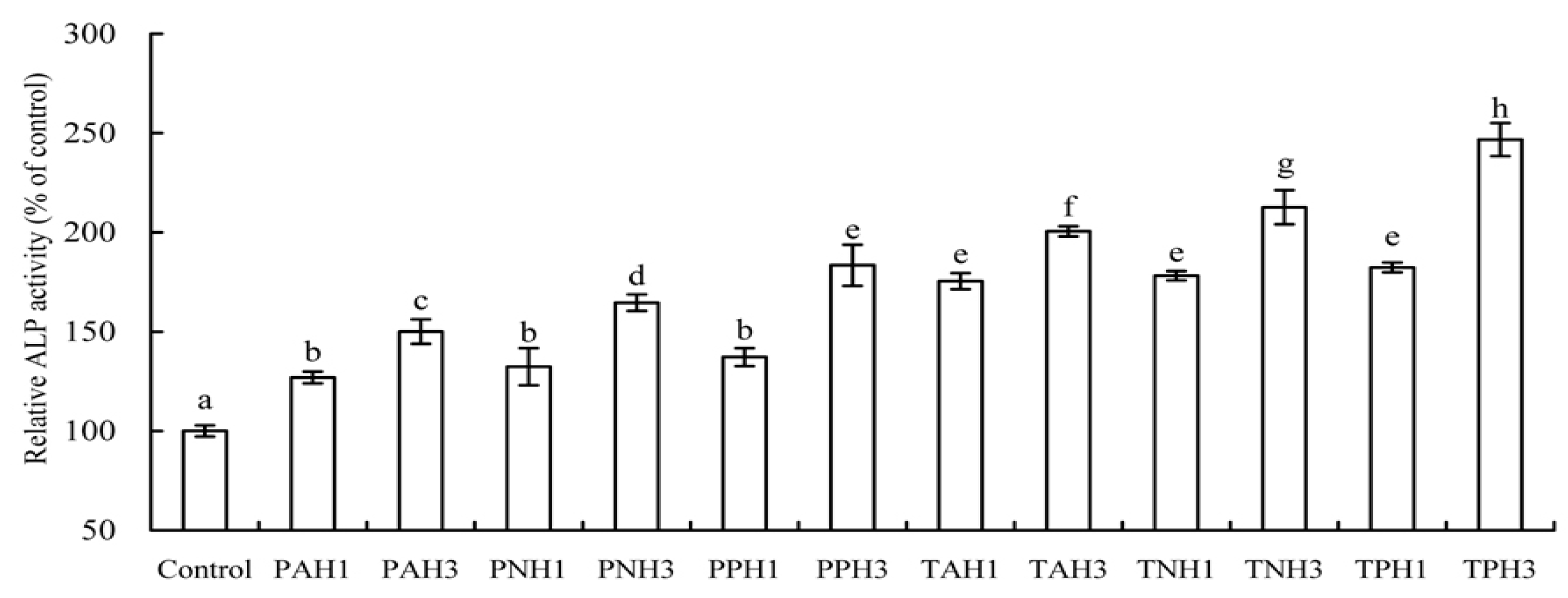
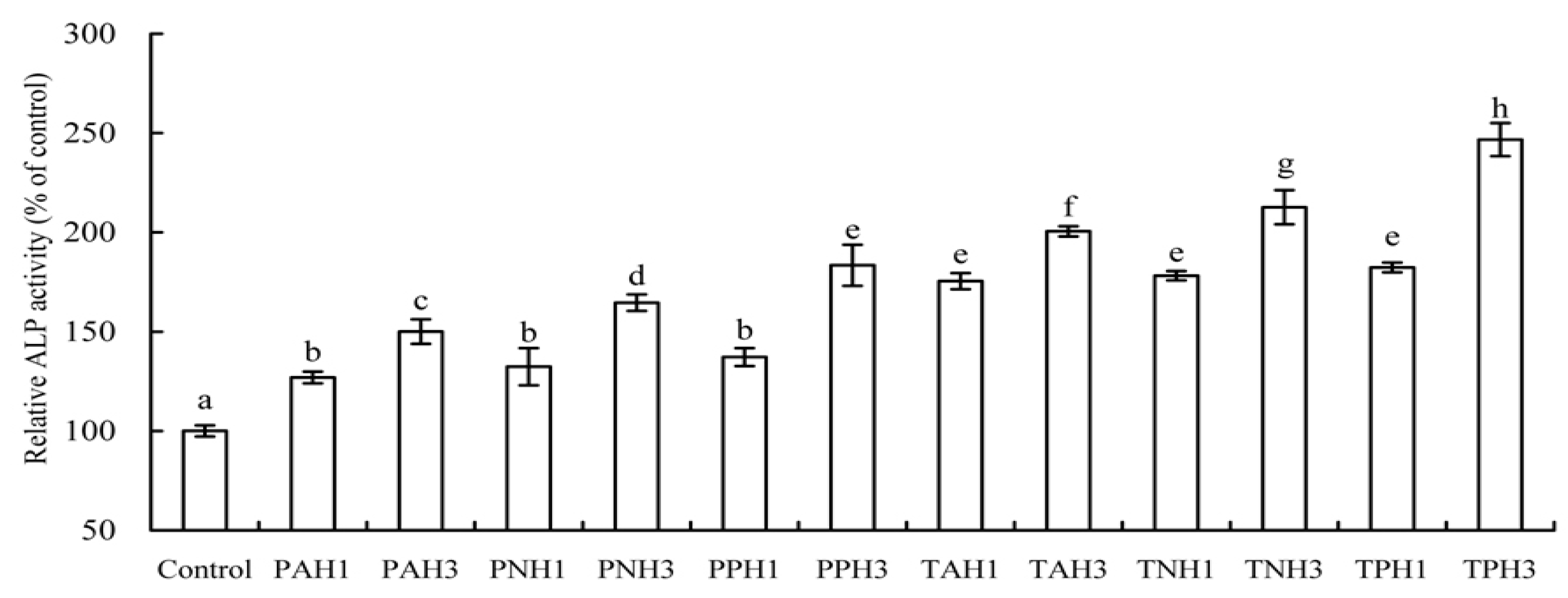
2.3. Differentiation Induction of Gelatin Hydrolysates to the Osteoblasts
2.4. Amino Acid Compositions of Gelatin Hydrolysates
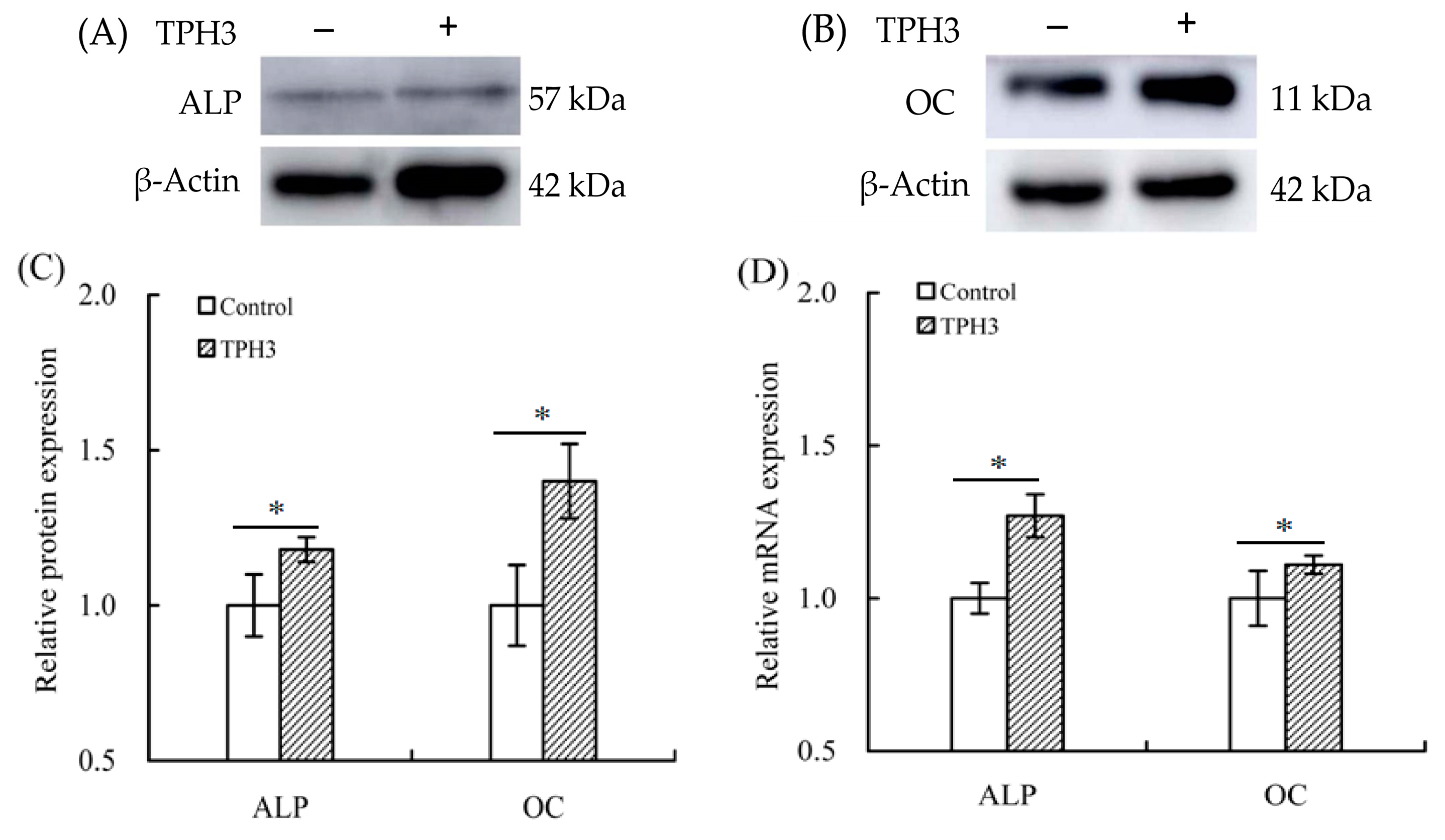
2.5. The Changes of Gene Expression in the Treated Osteoblasts
2.6. The Changes of Proteins Expression in the Treated Osteoblasts
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Preparation of the Hydrolysates
4.3. Chemical Analyses
4.4. Cell Line and Culture Conditions
4.5. Measurements of Growth Proliferation and Induced Apoptosis
4.6. Assays of Alkaline Phosphatase Activity and Osteocalcin Content
4.7. Real-Time RT-PCR Analysis
4.8. Western-Blot Assay
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| ALP | Alkaline phosphatase |
| BAH | Bovine skin gelatin hydrolysate by alcalase |
| BNH | Bovine skin gelatin hydrolysate by neutrase |
| BPH | Bovine skin gelatin hydrolysate by papain |
| CCK-8 | Cell counting kit-8 |
| DH | Degree of hydrolysis |
| DKK-1 | Dickkopf-1 |
| E2 | 17β-estradiol |
| EDTA | Ethylenediamine tetra-acetic acid |
| EP | Etoposide |
| ER-FBS | Estrogen-removed fetal bovine serum |
| FBS | Fetal bovine serum |
| hFOB 1.19 cells | Human fetal osteoblastic cells |
| PAH | Porcine skin gelatin hydrolysate by alcalcase |
| PBS | Phosphate-buffered saline |
| PNH | Porcine skin gelatin hydrolysate by neytrase |
| PPH | Provine skin gelatin hydrolysate by papain |
| TAH | Tilapia skin gelatin hydrolysate by alcalase |
| TNH | Tilapia skin gelatin hydrolysate by neutrase |
| TPH | Tilapia skin gelatin hydrolysate by papain |
References
- Raggatt, L.J.; Partridge, N.C. Cellular and molecular mechanisms of bone remodeling. J. Biol. Chem. 2010, 285, 25103–25108. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Lee, J.E.; Ryu, S.H.; Suh, P.G. Chlormadinone acetate promotes osteoblast differentiation of human mesenchymal stem cells through the ERK signaling pathway. Eur. J. Pharmacol. 2014, 726, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rodan, G.A.; Martin, T.J. Therapeutic approaches to bone diseases. Science 2000, 289, 1508–1514. [Google Scholar] [CrossRef] [PubMed]
- Jakab, L. Bone tissue: Rebuilding and inflammation. Orvosi Hetil. 2014, 155, 1575–1583. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.H.; Chen, C.; Zhang, Z.P.; Wu, Z.G.; Fan, X.L.; Zhang, Z.Y.; Di, W.; Shi, L. Ginkgo biloba extract promotes osteogenic differentiation of human bone marrow mesenchymal stem cells in a pathway involving Wnt/β-catenin signaling. Pharmacol. Res. 2015, 97, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Dennison, E.; Mohamed, M.A.; Cooper, C. Epidemiology of osteoporosis. Rheum. Dis. Clin. N. Am. 2006, 32, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Gambacciani, M.; Vacca, F. Postmenopausal osteoporosis and hormone replacement therapy. Minerva Med. 2004, 95, 507–520. [Google Scholar] [PubMed]
- Neuprez, A.; Reginster, J.Y. Bone-forming agents in the management of osteoporosis. Best Pract. Res. Clin. Endocrinol. Metab. 2008, 22, 869–883. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.F.; Yang, T.S.; Chiu, W.C.; Hsu, C.Y.; Chou, C.L.; Su, Y.J.; Lai, H.M.; Chen, Y.C.; Chen, C.J.; Cheng, T.T. Non-adherence to anti-osteoporotic medications in Taiwan: Physician specialty makes a difference. J. Bone Miner. Metab. 2013, 31, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Saag, K.G.; Shane, E.; Boonen, S.; Marín, F.; Donley, D.W.; Taylor, K.A.; Dalsky, G.P.; Marcus, R. Teriparatide or alendronate in glucocorticoid-induced osteoporosis. N. Engl. J. Med. 2007, 357, 2028–2039. [Google Scholar] [CrossRef] [PubMed]
- Lambert, M.N.T.; Thybo, C.B.; Lykkeboe, S.; Rasmussen, L.M.; Frette, X.; Christensen, L.P.; Jeppesen, P.B. Combined bioavailable isoflavones and probiotics improve bone status and estrogen metabolism in postmenopausal osteopenic women: A randomized controlled trial. Am. J. Clin. Nutr. 2017, 106, 909–920. [Google Scholar] [CrossRef] [PubMed]
- Takada, Y.; Kobayashi, N.; Kato, K.; Matsuyama, H.; Yahiro, M.; Aoe, S. Effects of whey protein on calcium and bone metabolism in ovariectomized rats. J. Nutr. Sci. Vitaminol. 1997, 43, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Moskowitz, R.W. Role of collagen hydrolysate in bone and joint disease. Semin. Arthritis Rheum. 2000, 30, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Picherit, C.; Bennetau-Pelissero, C.; Chanteranne, B.; Lebecque, P.; Davicco, M.J.; Barlet, J.P.; Coxam, V. Soybean isoflavones dose-dependently reduce bone turnover but do not reverse established osteopenia in adult ovariectomized rats. J. Nutr. 2001, 131, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Schadow, S.; Siebert, H.C.; Lochnit, G.; Kordelle, J.; Rickert, M.; Steinmeyer, J. Collagen metabolism of human osteoarthritic articular cartilage as modulated by bovine collagen hydrolysates. PLoS ONE 2013, 8, e53955. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Fujioka, M.; Sugimoto, K.; Mu, G.; Ishimi, Y. Assessment of effectiveness of oral administration of collagen peptide on bone metabolism in growing and mature rats. J. Bone Miner. Metab. 2004, 22, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S.; Park, J.H.; Kawashima, E.; Ezawa, I.; Omi, N. Hydrolyzed collagen intake increases bone mass of growing rats trained with running exercise. J. Int. Soc. Sports Nutr. 2013, 10, e35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, Y.; Oohashi, K.; Watanabe, M.; Kasugai, S. Increase in bone mineral density through oral administration of shark gelatin to ovariectomized rats. Nutrition 2005, 21, 1120–1126. [Google Scholar] [CrossRef] [PubMed]
- Tsuruoka, N.; Yamato, R.; Sakai, Y.; Yoshitake, Y.; Yonekura, H. Promotion by collagen tripeptide of type I collagen gene expression in human osteoblastic cells and fracture healing of rat femur. Biosci. Biotechnol. Biochem. 2007, 71, 2680–2687. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zhao, X.H. In vitro responses of hFOB 1.19 cells towards chum salmon (Oncorhynchus keta) skin gelatin hydrolysates in cell proliferation, cycle progression and apoptosis. J. Funct. Foods 2013, 5, 279–288. [Google Scholar] [CrossRef]
- Hyung, J.H.; Ahn, C.B.; Je, J.Y. Blue mussel (Mytilus edulis) protein hydrolysate promotes mouse mesenchymal stem cell differentiation into osteoblasts through up-regulation of bone morphogenetic protein. Food Chem. 2018, 242, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.L.; Zhang, B.; Song, S.J.; Ma, M.; Si, S.Y.; Wang, Y.H.; Xu, B.X.; Feng, K.; Wu, J.G.; Guo, Y.C. Bovine collagen peptides compounds promote the proliferation and differentiation of MC3T3-E1 pre-osteoblasts. PLoS ONE 2014, 9, e99920. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Hirota, A.; Mori, H.; Takahara, H.; Kuwaba, K.; Kusubata, M.; Matsubara, Y.; Kasugai, S.; Itoh, M.; Irie, S. Ingestion of gelatin has differential effect on bone mineral density and body weight in protein undernutrition. J. Nutr. Sci. Vitaminol. 2001, 47, 84–86. [Google Scholar] [CrossRef] [PubMed]
- Guillerminet, F.; Beaupied, H.; Fabien-Soulé, V.; Tomé, D.; Benhamou, C.L.; Roux, C.; Blais, A. Hydrolyzed collagen improves bone metabolism and biomechanical parameters in ovariectomized mice: An in vitro and in vivo study. Bone 2010, 46, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Kim, M.G.; Leem, K.H. Osteogenic activity of collagen peptide via ERK/MAPK pathway mediated boosting of collagen synthesis and its therapeutic efficacy in osteoporotic bone by back-scattered electron imaging and microarchitecture analysis. Molecules 2013, 18, 15474–15489. [Google Scholar] [CrossRef] [PubMed]
- Ohara, H.; Matsumoto, H.; Ito, K.; Iwai, K.; Sato, K. Comparison of quantity and structures of hydroxyproline-containing peptides in human blood after oral ingestion of gelatin hydrolysates from different sources. J. Agric. Food Chem. 2007, 55, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.P.; Boyce, B.F. Regulation of apoptosis in osteoclasts and osteoblastic cells. Biochem. Biophys. Res. Commun. 2005, 328, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Jilka, R.L.; Weinstein, R.S.; Bellido, T.; Parfitt, A.M.; Manolagas, S.C. Osteoblast programmed cell death (apoptosis): Modulation by growth factors and cytokines. J. Bone Miner. Res. 1998, 13, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.E.; Dai, A.; Tiffee, J.C.; Li, H.H.; Mundy, G.R.; Boyce, B.F. Estrogen promotes apoptosis of murine osteoclasts mediated by TGF-β. Nat. Med. 1996, 2, 1132–1136. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.R.; Plotkin, L.I.; Aguirre, J.I.; Han, L.; Jilka, R.L.; Kousteni, S.; Bellido, T.; Manolagas, S.C. Transient versus sustained phosphorylation and nuclear accumulation of ERKs underlie anti-versus pro-apoptotic effects of estrogens. J. Biol. Chem. 2005, 280, 4632–4638. [Google Scholar] [CrossRef] [PubMed]
- Kousteni, S.; Bellido, T.; Plotkin, L.I.; O’Brien, C.A.; Bodenner, D.L.; Han, L.; Han, K.; DiGregorio, G.B.; Katzenellenbogen, J.A.; Katzenellenbogen, B.S.; et al. Nongenotropic, sex-nonspecific signaling through the estrogen or androgen receptors: Dissociation from transcriptional activity. Cell 2001, 104, 719–730. [Google Scholar] [CrossRef]
- Wang, S.; Fu, Y.; Zhao, X.H. The cooperative effect of genistein and protein hydrolysates on the proliferation and survival of osteoblastic cell (hFOB 1.19). Molecules 2016, 21, 1489. [Google Scholar] [CrossRef] [PubMed]
- Bao, Q.W.; Chen, S.X.; Qin, H.; Feng, J.Q.; Liu, H.Y.; Liu, D.C.; Li, A.; Shen, Y.; Zhong, X.Z.; Li, J.F.; et al. Constitutive β-catenin activation in osteoblasts impairs terminal osteoblast differentiation and bone quality. Exp. Cell Res. 2017, 350, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Reddi, S.; Kumar, N.; Vij, R.; Mada, S.B.; Kapila, S.; Kapila, R. Akt drives buffalo casein-derived novel peptide-mediated osteoblast differentiation. J. Nutr. Biochem. 2016, 38, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Orimo, H. The mechanism of mineralization and the role of alkaline phosphatase in health and disease. J. Nippon Med. Sch. 2010, 77, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Patti, A.; Gennari, L.; Merlotti, D.; Dotta, F.; Nuti, R. Endocrine actions of osteocalcin. Int. J. Endocrinol. 2013, 2013, e846480. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, T.; Nanbu, P.N.; Kurokawa, M. Distribution of prolylhydroxyproline and its metabolites after oral administration in rats. Biol. Pharm. Bull. 2012, 35, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Nakatani, S.; Mano, H.; Sampei, C.; Shimizu, J.; Wada, M. Chondroprotective effect of the bioactive peptide prolyl-hydroxyproline in mouse articular cartilage in vitro and in vivo. Osteoarthr. Cartil. 2009, 17, 1620–1627. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.W.; Zhao, X.H. In vitro proliferation and anti-apoptosis of the papain-generated casein and soy protein hydrolysates towards osteoblastic cells (hFOB1.19). Int. J. Mol. Sci. 2015, 16, 13908–13920. [Google Scholar] [CrossRef] [PubMed]
- Price, M.A. CKI, there’s more than one: Casein kinase I family members in Wnt and Hedgehog signaling. Genes Dev. 2006, 20, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Glass, D.A.; Bialek, P.; Ahn, J.D.; Starbuck, M.; Patel, M.S.; Clevers, H.; Taketo, M.M.; Long, F.; McMahon, A.P.; Lang, R.A.; et al. Canonical Wnt signaling in differentiated osteoblasts controls osteoclast differentiation. Dev. Cell 2005, 8, 751–764. [Google Scholar] [CrossRef] [PubMed]
- Holmen, S.L.; Zylstra, C.R.; Mukherjee, A.; Sigler, R.E.; Faugere, M.C.; Bouxsein, M.L.; Deng, L.; Clemens, T.L.; Williams, B.O. Essential role of beta-catenin in postnatal bone acquisition. J. Biol. Chem. 2005, 280, 21162–21168. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, T. Wnt/β-catenin signaling. Cytokine Growth Factor Rev. 2000, 11, 273–282. [Google Scholar] [CrossRef]
- Chen, D.; Zhao, M.; Mundy, G.R. Bone morphogenetic proteins. Growth Factors 2004, 22, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Dang, C.V. c-Myc target genes involved in cell growth, apoptosis, and metabolism. Mol. Cell. Biol. 1999, 19, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mirando, A.J.; Maruyama, T.; Fu, J.; Yu, H.M.I.; Hsu, W. Beta-catenin/cyclin D1 mediated development of suture mesenchyme in calvarial morphogenesis. BMC Dev. Biol. 2010, 10, 116. [Google Scholar] [CrossRef] [PubMed]
- Piek, E.; Sleumer, L.S.; van Someren, E.P.; Heuver, L.; de Haan, J.R.; de Grijs, I.; Gilissen, C.; Hendriks, J.M.; van Ravestein-van Os, R.I.; Bauerschmidt, S.; et al. Osteo-transcriptomics of human mesenchymal stem cells: Accelerated gene expression and osteoblast differentiation induced by vitamin D reveals c-MYC as an enhancer of BMP2-induced osteogenesis. Bone 2010, 46, 613–627. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.O.; Insogna, K.L. Where Wnts went: The exploding field of Lrp 5 and Lrp 6 signaling in bone. J. Bone Miner. Res. 2009, 24, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Daoussis, D.; Andonopoulos, A.P. The emerging role of Dickkopf-1 in bone biology: Is it the main switch controlling bone and joint remodeling? Semin. Arthritis Rheum. 2011, 41, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Eckert, R.L.; Katzenallenbogen, B.S. Effect of estrogens and antiestrogen receptor dynamics and the induction of progesterone receptor in MCF-7 human breast cancer cells. Cancer Res. 1982, 42, 139–144. [Google Scholar] [PubMed]
- Association of Official Agricultural Chemists (AOAC). Official Methods of Analysis of Association of Official Analytical Chemists International, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Bergman, I.; Loxley, R. Two improved and simplified methods for the spectrophotometric determination of hydroxyproline. Anal. Chem. 1963, 35, 1961–1965. [Google Scholar] [CrossRef]
- Harris, S.A.; Enger, R.J.; Riggs, B.L.; Spelsberg, T.C. Development and characterization of a conditionally immoralized human fetal osteoblastic cell line. J. Bone Miner. Res. 1995, 10, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Miao, T.X.; Rao, K.S.; Spees, J.L.; Oldinski, R.A. Osteogenic differentiation of human mesenchymal stem cells through alginate-graft-poly (ethylene glycol) microsphere-mediated inracellular growth factor delivery. J. Control. Release 2014, 192, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Vermes, I.; Haanen, C.; Steffens-Nakken, H.; Reutellingsperger, C. A novel assay for apoptosis Flow cytometric detection of phosphatidylserine expression on early apoptotic cells using fluorescein labelled Annexin V. J. Immunol. Methods 1995, 184, 39–51. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆Ct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, Y.F.; Xiong, Y.J.; Wang, W.J.; Fei, Y.; Tan, C.H.; Liang, Z.Q. K-ras mutation promotes ionizing radiation-induced invasion and migration of lung cancer in part via the Cathepsin L/CUX1 pathway. Exp. Cell Res. 2018, 362, 424–435. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds gelatin hydrolysates are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hydrolysates | Hydrolysis Times (h) | Proteases and Used Levels | DH (%) |
|---|---|---|---|
| BAH1 | 0.5 | Alcalase, 5.5 kU/g protein | 7.1 |
| BAH2 | 3.5 | 11.2 | |
| BAH3 | 13 | 14.6 | |
| BNH1 | 0.5 | Neutrase, 6.0 kU/g protein | 6.7 |
| BNH2 | 2 | 10.7 | |
| BNH3 | 11 | 14.8 | |
| BPH1 | 1 | Papain, 1.5 kU/g protein | 7.3 |
| BPH2 | 15 | 10.8 | |
| PAH1 | 0.25 | Alcalase, 7.0 kU/g protein | 6.5 |
| PAH2 | 3 | 11.1 | |
| PAH3 | 12.5 | 15.2 | |
| PNH1 | 0.3 | Neutrase, 6.0 kU/g protein | 6.5 |
| PNH2 | 3 | 10.8 | |
| PNH3 | 12.5 | 15.2 | |
| PPH1 | 0.25 | Papain, 1.5 kU/g protein | 7.2 |
| PPH2 | 3 | 11.0 | |
| PPH3 | 12.5 | 15.0 | |
| TAH1 | 0.3 | Alcalase, 6.0 kU/g protein | 6.8 |
| TAH2 | 3 | 11.2 | |
| TAH3 | 13 | 14.6 | |
| TNH1 | 0.5 | Neutrase, 6.0 kU/g protein | 7.3 |
| TNH2 | 2 | 10.9 | |
| TNH3 | 12 | 14.8 | |
| TPH1 | 0.25 | Papain, 1.5 kU/g protein | 7.2 |
| TPH2 | 1.5 | 10.5 | |
| TPH3 | 7 | 15.4 |
| Samples | Hydrolysate Doses and Treating Times of the Cells | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 0.02 g/L | 0.05 g/L | 0.1 g/L | |||||||
| 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | |
| BAH1 | 107.2 ± 4.8abcd | 112.4 ± 4.8abcdefg | 107.9 ± 4.0abcdef | 115.8 ± 1.9abcde | 117.0 ± 4.6ab | 113.9 ± 2.7abcd | 104.0 ± 3.3a | 109.9 ± 5.7abc | 112.6 ± 3.3defghij |
| BAH2 | 108.6 ± 2.0abcd | 112.7 ± 5.3abcdefg | 108.1 ± 3.0abcdef | 119.8 ± 6.8abcde | 118.2 ± 3.2abc | 118.3 ± 4.0abcdefg | 104.5 ± 5.7ab | 107.6 ± 5.4a | 117.6 ± 4.6ghij |
| BAH3 | 116.8 ± 4.4ef | 119.5 ± 3.4efg | 113.5 ± 3.4fghij | 122.5 ± 3.6cde | 123.6 ± 4.3abcde | 119.3 ± 2.0cdefg | 114.7 ± 4.2defg | 114.2 ± 2.4abcdefg | 115.0 ± 1.6efghij |
| BNH1 | 107.7 ± 4.6abcd | 108.7 ± 4.4ab | 105.7 ± 5.7abcd | 114.9 ± 4.4abc | 116.9 ± 5.9ab | 111.3 ± 5.1a | 107.2 ± 4.9abcd | 108.5 ± 3.7ab | 107.8 ± 3.6abcde |
| BNH2 | 108.9 ± 2.6abcde | 112.1 ± 3.6abcdef | 108.3 ± 3.7abcdef | 115.5 ± 1.0abcd | 118.6 ± 1.7abc | 115.3 ± 5.1abcde | 104.1 ± 5.4a | 110.6 ± 4.0abc | 107.5 ± 5.1abcd |
| BNH3 | 111.0 ± 1.9bcdef | 119.3 ± 5.4defg | 109.4 ± 1.8bcdefg | 117.2 ± 2.2abcde | 125.6 ± 5.2cde | 117.2 ± 4.0abcdef | 108.3 ± 4.0abcde | 115.2 ± 2.6abcdefg | 108.7 ± 3.3abcde |
| BPH1 | 104.6 ± 4.3abc | 109.3 ± 6.2abc | 109.6 ± 2.1bcdefg | 114.4 ± 3.4abc | 116.5 ± 2.7a | 114.8 ± 3.0abcd | 108.6 ± 2.6abcde | 113.1 ± 2.6abcde | 110.7 ± 1.6bcdefg |
| BPH2 | 106.3 ± 2.6abcd | 115.0 ± 2.5abc | 111.7 ± 2.0defghi | 116.1 ± 1.7abcde | 122.8 ± 4.6abcde | 119.5 ± 2.3cdefg | 109.2 ± 4.6abcde | 110.7 ± 2.6abc | 112.4 ± 2.9defghi |
| PAH1 | 101.4 ± 2.9a | 108.8 ± 3.3abc | 102.4 ± 1.9a | 113.4 ± 5.8ab | 120.4 ± 3.5abcd | 111.7 ± 3.4ab | 104.6 ± 1.4ab | 110.6 ± 3.5abc | 104.9 ± 2.7abc |
| PAH2 | 103.5 ± 5.3ab | 109.7 ± 5.6abc | 103.7 ± 2.4ab | 119.9 ± 4.2abcde | 123.3 ± 4.7abcde | 112.9 ± 5.0abc | 106.3 ± 2.4abc | 114.9 ± 4.8abcdefg | 104.4 ± 4.8ab |
| PAH3 | 107.4 ± 4.6abcd | 105.4 ± 1.9a | 107.5 ± 3.1abcdef | 122.7 ± 4.3cde | 124.0 ± 3.6abcde | 115.9 ± 5.4abcdef | 114.8 ± 4.5defg | 115.1 ± 5.5abcdefg | 111.4 ± 5.8bcdefgh |
| PNH1 | 106.1 ± 4.1abcd | 110.7 ± 3.0abcd | 109.7 ± 1.4bcdefg | 115.7 ± 4.6abcde | 121.9 ± 4.3abcde | 113.3 ± 5.4abc | 112.4 ± 1.7bcdefg | 111.8 ± 5.4abcd | 116.1 ± 3.4fghij |
| PNH2 | 111.6 ± 4.6bcdef | 116.5 ± 5.2bcdefg | 111.6 ± 3.5defgh | 121.8 ± 2.4cde | 124.9 ± 5.3bcde | 117.9 ± 3.4abcdefg | 117.1 ± 3.5fg | 121.0 ± 2.6efghi | 108.7 ± 2.3abcde |
| PNH3 | 112.4 ± 4.6cdef | 119.7 ± 4.9efg | 114.9 ± 2.9ghij | 124.0 ± 4.1e | 127.9 ± 2.4def | 118.9 ± 2.2bcdefg | 109.4 ± 4.3abcde | 116.7 ± 4.8bcdefg | 111.8 ± 5.0cdefgh |
| PPH1 | 107.6 ± 5.9abcd | 115.5 ± 5.6bcdefg | 106.9 ± 4.7abcdef | 115.6 ± 5.1abcd | 122.9 ± 3.3abcde | 115.3 ± 4.5abcde | 109.5 ± 3.0abcde | 113.3 ± 3.4abcdef | 102.4 ± 4.8a |
| PPH2 | 111.8 ± 5.9bcdef | 117.4 ± 4.4cdefg | 110.4 ± 4.0cdefgh | 118.5 ± 5.1abcde | 126.8 ± 2.6de | 120.2 ± 3.6cdefg | 112.8 ± 4.3cdefg | 117.9 ± 5.2cdefgh | 113.2 ± 2.9defghij |
| PPH3 | 113.7 ± 4.6def | 120.3 ± 4.6fg | 116.4 ± 2.8hij | 119.0 ± 5.4abcde | 134.4 ± 4.3fg | 124.9 ± 2.3g | 116.1 ± 5.0efg | 128.5 ± 5.1i | 119.6 ± 4.4j |
| TAH1 | 111.2 ± 5.3bcdef | 112.0 ± 5.9abcdef | 109.4 ± 2.9bcdefg | 114.4 ± 3.3abc | 121.8 ± 4.7abcde | 113.0 ± 5.3abc | 111.6 ± 3.5abcdef | 119.6 ± 2.3defgh | 110.7 ± 2.3bcdefg |
| TAH2 | 111.9 ± 4.1bcdef | 113.5 ± 3.6abcdefg | 111.3 ± 4.3defgh | 121.2 ± 1.5bcde | 123.7 ± 3.5abcde | 116.0 ± 1.0abcdef | 110.2 ± 3.3abcdef | 116.1 ± 5.5abcdefg | 110.9 ± 3.1bcdefg |
| TAH3 | 118.3 ± 2.6f | 119.9 ± 3.7efg | 118.2 ± 5.3ij | 122.0 ± 4.5cde | 135.5 ± 3.9g | 121.3 ± 2.8defg | 119.7 ± 2.1g | 121.6 ± 2.7fghi | 118.6 ± 3.2hij |
| TNH1 | 106.5 ± 4.4abcd | 112.9 ± 2.1abcdefg | 106.3 ± 3.8abcde | 111.8 ± 3.9a | 122.6 ± 4.2abcde | 119.0 ± 4.2bcdefg | 107.7 ± 5.6abcd | 115.7 ± 2.9abcdefg | 112.9 ± 3.0defghij |
| TNH2 | 106.9 ± 3.9abcd | 114.3 ± 2.9bcdefg | 112.9 ± 2.2efghij | 118.3 ± 5.8abcde | 125.8 ± 5.0cde | 122.6 ± 4.7efg | 108.1 ± 4.3abcd | 116.6 ± 5.8bcdefg | 114.0 ± 2.1defghij |
| TNH3 | 108.9 ± 4.4abcde | 116.6 ± 4.4bcdefg | 112.1 ± 2.2defghi | 123.6 ± 3.2de | 129.0 ± 4.2efg | 124.8 ± 3.4g | 110.2 ± 2.4abcdef | 122.1 ± 2.1ghi | 119.0 ± 4.7ij |
| TPH1 | 108.3 ± 5.3abcd | 111.4 ± 5.5abcde | 104.3 ± 4.3abc | 115.4 ± 4.8abcd | 122.3 ± 4.3abcde | 110.9 ± 1.6a | 110.3 ± 3.9abcdef | 113.9 ± 6.3abcdefg | 109.3 ± 3.6abcdef |
| TPH2 | 107.9 ± 4.6abcd | 119.2 ± 2.8defg | 113.4 ± 2.3fghij | 115.1 ± 5.7abc | 124.1 ± 3.8abcde | 123.3 ± 3.7fg | 106.1 ± 4.9abc | 119.6 ± 4.6defgh | 117.0 ± 4.2ghij |
| TPH3 | 110.7 ± 2.7bcdef | 120.8 ± 3.7g | 118.7 ± 2.6j | 118.4 ± 4.4abcde | 135.7 ± 2.8g | 125.3 ± 4.2g | 109.2 ± 4.6abcde | 125.4 ± 5.2hi | 119.2 ± 3.0ij |
| Amino Acids | BAH3 | BNH3 | BPH2 | PAH3 | PNH3 | PPH3 | TAH3 | TNH3 | TPH3 |
|---|---|---|---|---|---|---|---|---|---|
| Ala | 98.7 | 98.9 | 94.1 | 100.0 | 98.6 | 99.6 | 103.9 | 101.6 | 101.3 |
| Arg | 98.7 | 98.9 | 94.1 | 96.4 | 96.8 | 95.9 | 100.4 | 101.6 | 97.6 |
| Asp | 40.2 | 39.6 | 40.0 | 41.1 | 48.4 | 40.6 | 40.5 | 35.7 | 39.8 |
| Cys | 7.3 | 7.2 | 7.1 | 7.1 | 7.2 | 7.4 | 7.0 | 7.1 | 7.2 |
| Glu | 87.8 | 88.1 | 89.4 | 89.3 | 91.4 | 88.6 | 88.0 | 87.3 | 86.8 |
| Gly | 223.0 | 226.6 | 221.2 | 223.2 | 222.2 | 225.1 | 221.8 | 228.2 | 220.6 |
| His | 5.5 | 5.4 | 4.7 | 5.4 | 5.4 | 5.5 | 5.3 | 5.3 | 5.4 |
| 4-Hyp | 115.9 | 118.4 | 117.9 | 119.7 | 120.4 | 122.6 | 112.4 | 113.8 | 119.3 |
| Ile | 14.6 | 14.4 | 14.1 | 12.5 | 12.5 | 12.9 | 14.1 | 14.3 | 14.5 |
| Leu | 32.9 | 32.4 | 32.9 | 32.1 | 32.3 | 33.2 | 31.7 | 32.1 | 30.7 |
| Lys | 34.7 | 34.2 | 35.3 | 33.9 | 35.8 | 35.1 | 33.5 | 33.9 | 34.4 |
| Met | 12.8 | 12.6 | 11.8 | 10.7 | 10.8 | 11.1 | 14.1 | 14.3 | 14.5 |
| Phe | 18.3 | 18.0 | 18.8 | 17.9 | 19.7 | 18.5 | 19.4 | 19.6 | 18.1 |
| Pro | 133.5 | 134.9 | 131.8 | 137.5 | 127.2 | 136.5 | 132.0 | 133.7 | 130.2 |
| Ser | 27.4 | 25.2 | 25.9 | 26.8 | 30.5 | 24.0 | 28.2 | 23.2 | 28.9 |
| Thr | 7.3 | 9.0 | 9.4 | 10.7 | 12.5 | 11.1 | 10.6 | 14.3 | 10.8 |
| Tyr | 5.5 | 5.4 | 4.7 | 3.6 | 3.6 | 3.7 | 3.5 | 3.6 | 3.6 |
| Val | 20.1 | 21.6 | 23.5 | 19.6 | 21.5 | 22.1 | 19.4 | 19.6 | 19.9 |
| Genes | Primers (5′-3′) |
|---|---|
| Wnt 3a | F-GGTGGCTGTAGCGAGGACAT R-TTGTTGTGGCGGTTCATGGC |
| Wnt 10b | F-TCCTGACTTCTGTGAGCGAGACC R-CATAGCAGCACCAGTGGAAGCG |
| β-catenin | F-AGAAGGTCCGAGTGCTGCTC R-CTGAGCTGGCTGTTGACCAC |
| GSK 3β | F-TTCCTCCTCATGCTCGGATT R-CAGGTGGAGTTGGAAGCTGA |
| Cyclin D1 | F-CAGAAGTGCGAGGAGGAGGT R-TAGAGGCCACGAACATGCAA |
| c-Myc | F-CGAGGAGAATGTCAAGAGGCGAAC R-GCTTGGACGGACAGGATGTATGC |
| ALP | F-CGAGTGAACAGGAACAACG R-AATTCTGCCTCCTTCCACCA |
| Osteocalcin | F-CCAGGCGCTACCTGTATCAA R-GGTCAGCCAACTCGTCACAG |
| β-Actin | F-CCTGGCACCCAGCACAAT R-GGGCCGGACTCGTCATAC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, M.; Zhao, X.-H. The Growth Proliferation, Apoptotic Prevention, and Differentiation Induction of the Gelatin Hydrolysates from Three Sources to Human Fetal Osteoblasts (hFOB 1.19 Cells). Molecules 2018, 23, 1287. https://doi.org/10.3390/molecules23061287
Lu M, Zhao X-H. The Growth Proliferation, Apoptotic Prevention, and Differentiation Induction of the Gelatin Hydrolysates from Three Sources to Human Fetal Osteoblasts (hFOB 1.19 Cells). Molecules. 2018; 23(6):1287. https://doi.org/10.3390/molecules23061287
Chicago/Turabian StyleLu, Ming, and Xin-Huai Zhao. 2018. "The Growth Proliferation, Apoptotic Prevention, and Differentiation Induction of the Gelatin Hydrolysates from Three Sources to Human Fetal Osteoblasts (hFOB 1.19 Cells)" Molecules 23, no. 6: 1287. https://doi.org/10.3390/molecules23061287
APA StyleLu, M., & Zhao, X.-H. (2018). The Growth Proliferation, Apoptotic Prevention, and Differentiation Induction of the Gelatin Hydrolysates from Three Sources to Human Fetal Osteoblasts (hFOB 1.19 Cells). Molecules, 23(6), 1287. https://doi.org/10.3390/molecules23061287