
Cholinesterase Inhibition Activity, Alkaloid Profiling and Molecular Docking of Chilean Rhodophiala (Amaryllidaceae)

 ,
, 
Abstract
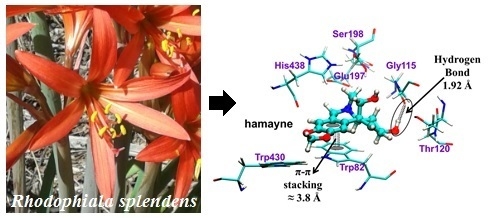
:
1. Introduction
2. Results and Discussion
2.1. AChE and BuChE Inhibitory Activities
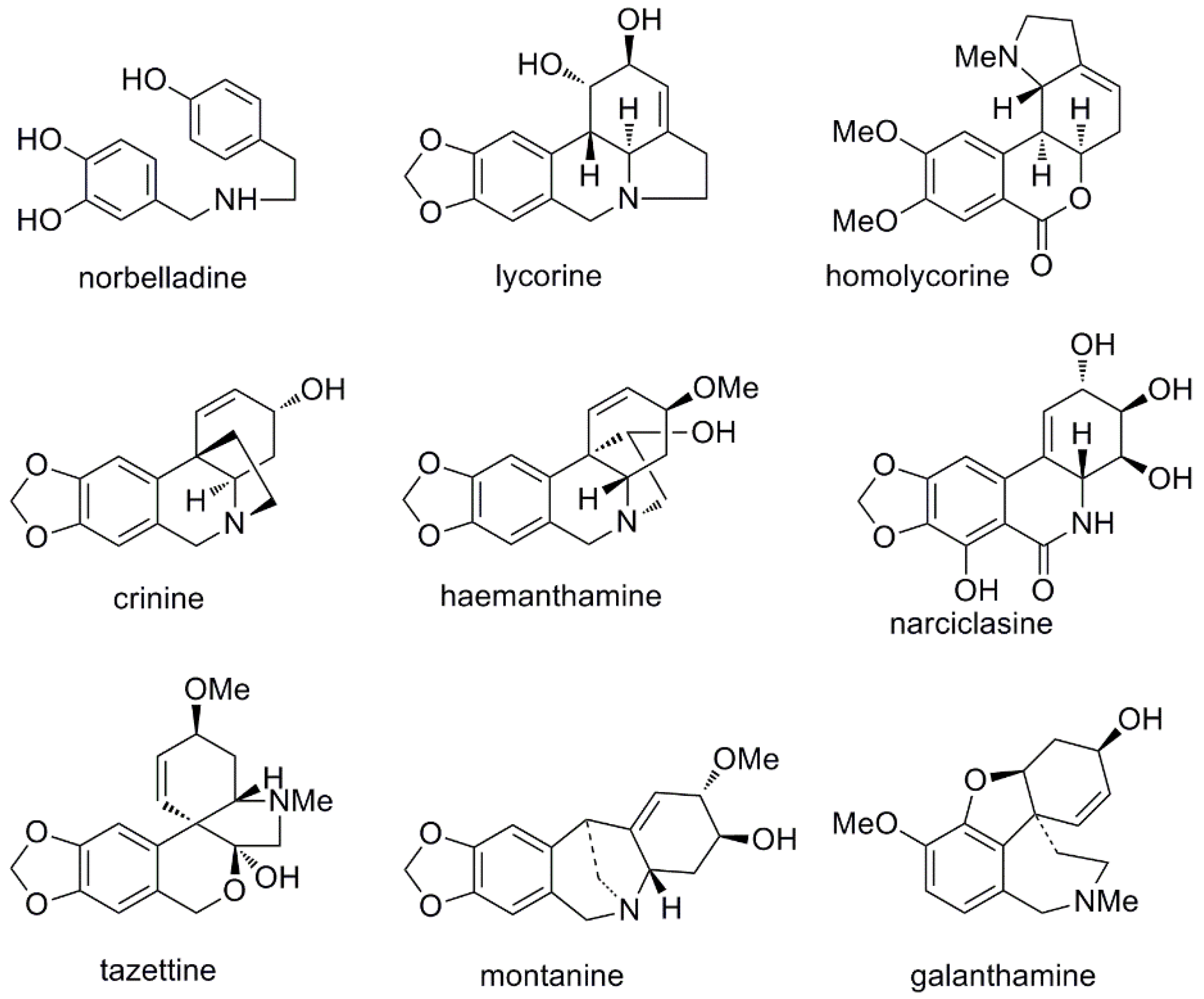
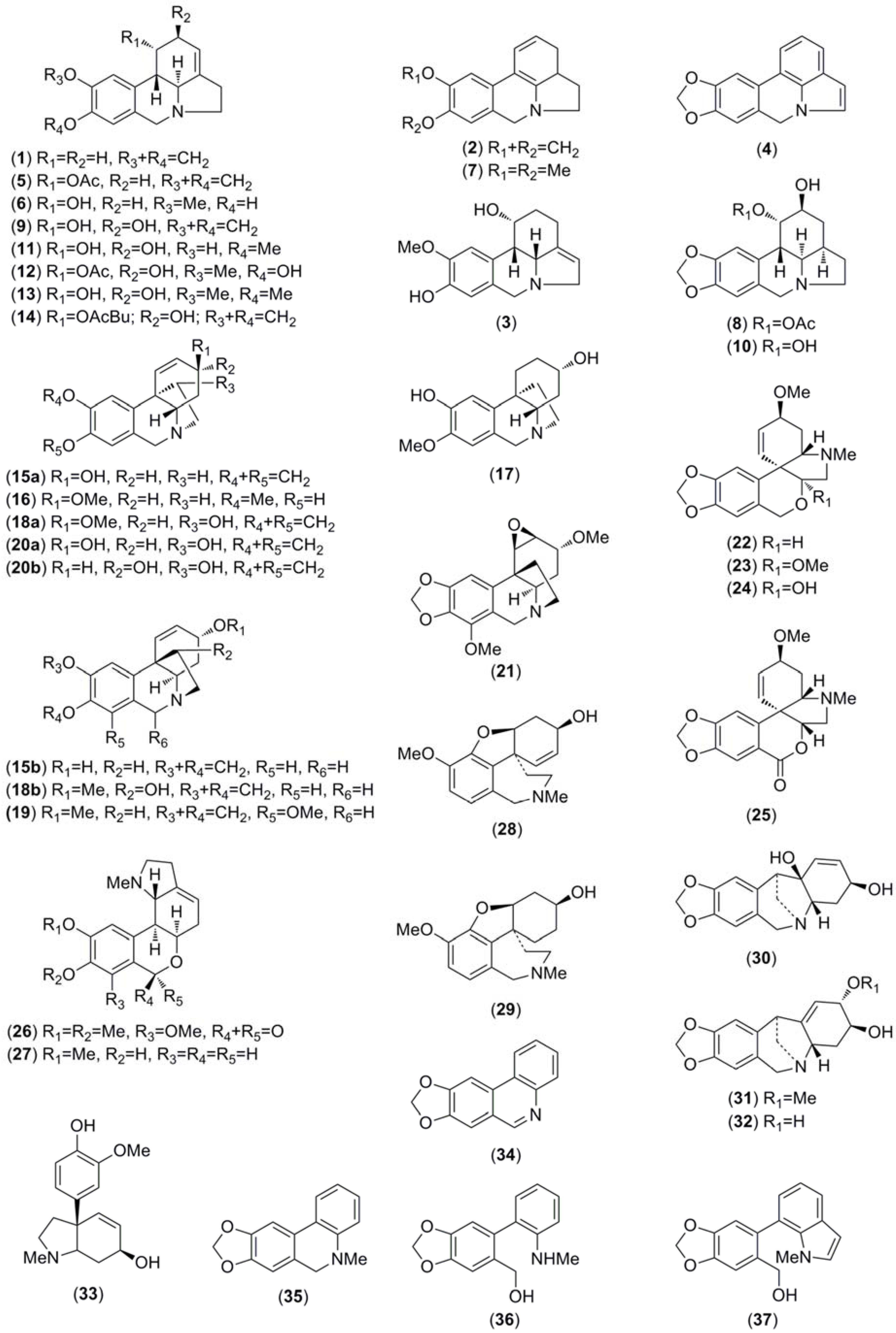
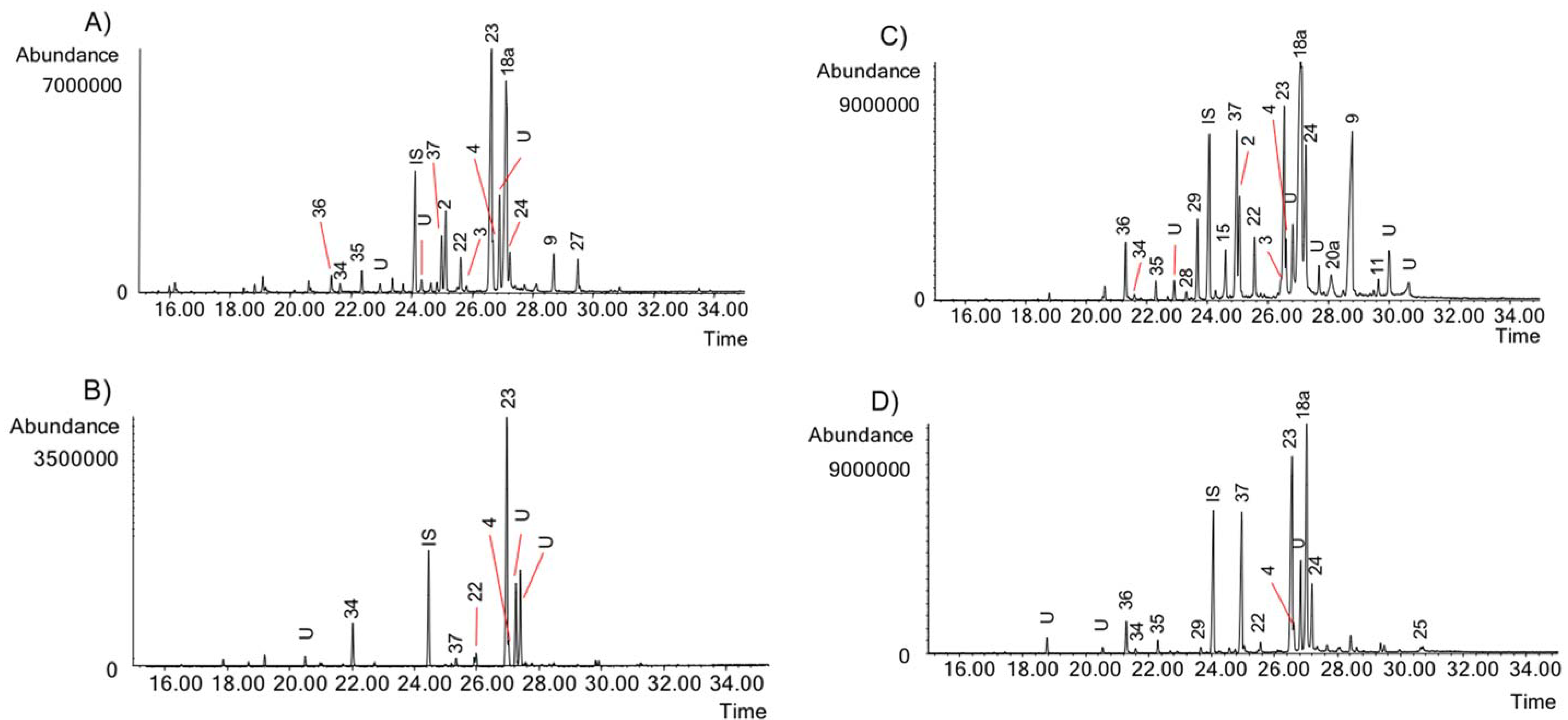
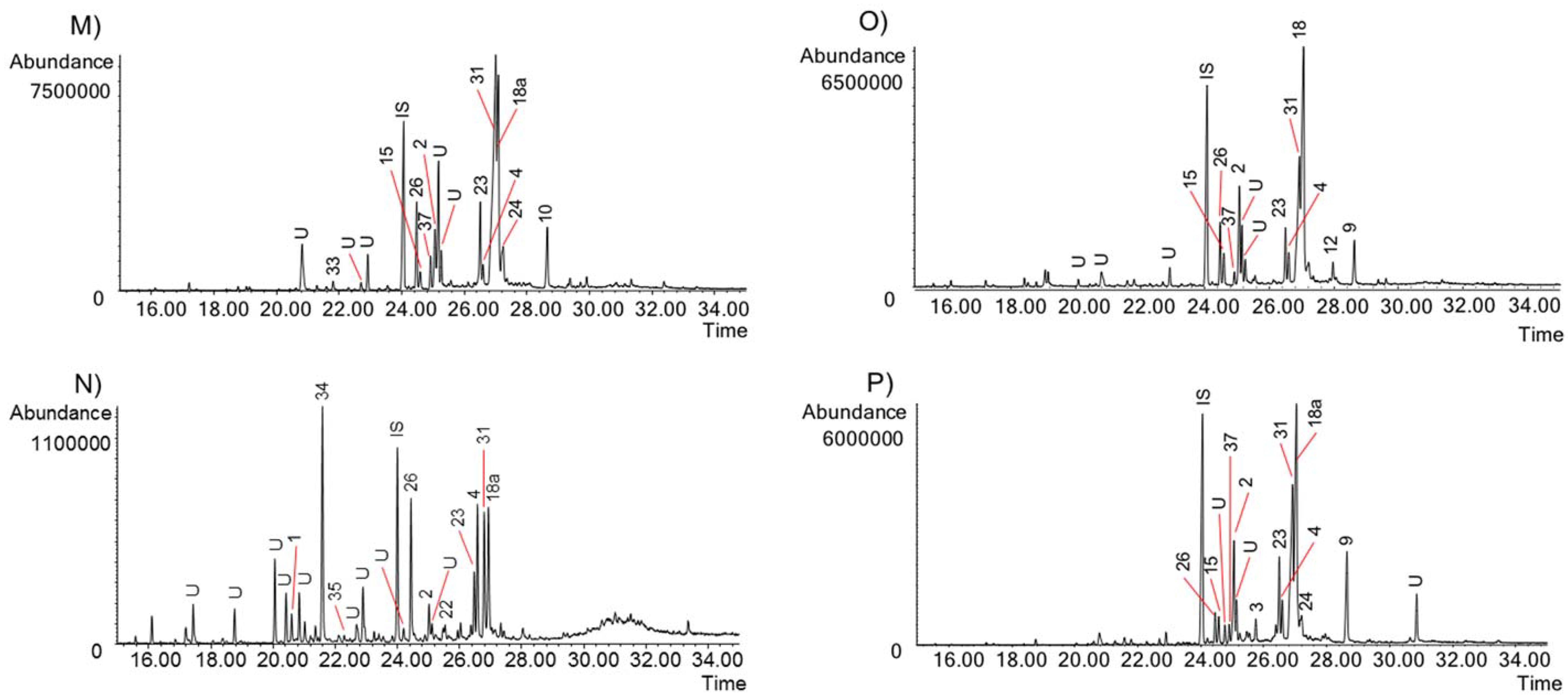
2.2. Alkaloid Identification by GC-MS
2.3. Molecular Docking
3. Materials and Methods
3.1. Plant Material
3.2. Extraction
3.3. Acetylcholinesterase (AChE) and Butyrylcholinesterase (BuChE) Inhibitory Activity
3.4. Alkaloids Identification and Quantification
3.4.1. Equipment
3.4.2. Alkaloids Identification
3.4.3. Alkaloid Quantification
3.5. Molecular Docking
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.-J.; Bao, J.-L.; Chen, X.-P.; Huang, M.; Wang, Y.-T. Alkaloids isolated from natural herbs as the anticancer agents. Evid.-Based Complement. Altern. Med. 2012, 2012, 485042. [Google Scholar] [CrossRef] [PubMed]
- Feher, M.; Schmidt, J.M. Property Distributions: Differences between drugs, natural products, and molecules from combinatorial chemistry. J. Chem. Inf. Comput. Sci. 2003, 43, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Bastida, J.; Lavilla, R.; Viladomat, F. Chemical and biological aspects of Narcissus alkaloids. In The Alkaloids: Chemistry and Physiology; Cordell, G.A., Ed.; Elsevier: Amsterdam, The Netherlands, 2006; Volume 63, pp. 87–179, eBook ISBN 9780080466552; Hardcover ISBN 9780124695634. [Google Scholar]
- Bastida, J.; Berkov, S.; Torras, L.; Pigni, N.B.; de Andrade, J.P.; Martínez, V.; Codina, C.; Viladomat, F. Chemical and biological aspects of Amaryllidaceae alkaloids. In Recent Advances in Pharmaceutical Sciences; Muñoz-Torrero, D., Ed.; Transworld Research Network: Kerala, India, 2011; pp. 65–100. ISBN 978-81-7895-528-5. [Google Scholar]
- Ingrassia, L.; Lefranc, F.; Mathieu, V.; Darro, F.; Kiss, R. Amaryllidaceae isocarbostyril alkaloids and their derivatives as promising antitumor agents. Transl. Oncol. 2008, 1, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Van Goiestsenoven, G.; Mathieu, V.; Lefranc, F.; Kornienko, A.; Evidente, A.; Kiss, R. Narciclasine as well as other Amaryllidaceae isocarnostyrils are promising GTP-ase targeting agents against brain cancers. Med. Res. Rev. 2013, 33, 439–455. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, M.; Teoh, H.L. Galanthamine from showdrop—The development of a modern drug against Alzheimer’s disease from local Caucasian knowledge. J. Ethnopharmacol. 2004, 9, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Maelicke, A.; Samochocki, M.; Jostock, R.; Fehrenbacher, A.; Ludwig, J.; Albuquerque, E.X.; Zerlin, M. Allosteric sensitization of nicotinic receptors by galanthamine, a new treatment strategy for Alzheimer’s disease. Biol. Psychiatry 2001, 49, 279–288. [Google Scholar] [CrossRef]
- Craig, L.A.; Hong, N.S.; McDonald, R.J. Revisiting the cholinergic hypothesis in the development of Alzheimer’s disease. Neurosci. Biobehav. Rev. 2011, 35, 1397–1409. [Google Scholar] [CrossRef] [PubMed]
- Querfurth, H.W.; LaFerla, F.M. Alzheimer’s disease. N. Engl. J. Med. 2010, 362, 329–344. [Google Scholar] [CrossRef] [PubMed]
- Alzheimer’s Disease International. Available online: https://www.alz.co.uk/research/WorldAlzheimerReport2016.pdf (accessed on 13 October 2017).
- Selkoe, D.J. Alzheimer’s disease: Genes, proteins, and therapy. Physiol. Rev. 2001, 81, 741–766. [Google Scholar] [CrossRef] [PubMed]
- Basiri, A.; Murugaiyah, V.; Osman, H.; Kumar, R.S.; Kia, Y.; Awang, K.B.; Ali, M.A. An expedient, ionic liquid mediated multi-component synthesis of novel piperidone grafted cholinesterase enzymes inhibitors and their molecular modeling study. Eur. J. Med. Chem. 2013, 67, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Greig, N.H.; Lahiri, D.K.; Sambamurti, K. Butyrylcholinesterase: An important new target in Alzheimer’s disease therapy. Int. Psychogeriatr. 2002, 14, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Giacobini, E. Cholinesterase inhibitors: New roles and therapeutic alternatives. Pharmacol. Res. 2004, 50, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Ballard, C.G. Advances in the treatment of Alzheimer’s disease: Benefits of dual cholinesterase inhibition. Eur. Neurol. 2002, 47, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Olate, E.; Bridgen, M. Techniques for the in vitro propagation of Rhodophiala and Leucocoryne spp. Acta Hortic. 2005, 673, 335–342. [Google Scholar] [CrossRef]
- Schiappacasse, F.; Peñailillo, P.; Yáñez, P. Propagación de Bulbosas Chilenas Ornamentals; Editorial Universidad de Talca: Talca, Chile, 2002; ISBN 956-7059-50-0. [Google Scholar]
- Muñoz, M.; Riegel, R.; Seemann, P.; Penailillo, P.; Schiappacasse, F.; Nunez, J. Phylogenetic relationships of Rhodolirium montanum Phil. and related species based on nucleotide sequences from ITS region and karyotype analysis. Gayana Bot. 2011, 68, 40–48. [Google Scholar] [CrossRef]
- Schiappacasse, F.; Peñailillo, P.; Basoalto, A.; Seemann, P.; Riegel, R.; Muñoz, M.; Jara, G.; Durán, C. Biotechnological applications on plant breeding of Chilean Rhodophiala species: Morphological and physiological studies. Agro Sur 2007, 35, 65–67. [Google Scholar] [CrossRef]
- Baeza, C.; Almendras, F.; Ruiz, E.; Peñailillo, P. Comparative karyotype studies in species of Miltinea Ravenna, Phycella Lindl. and Rhodophiala C. Presl (Amaryllidaceae) from Chile. Rev. Fac. Cienc. Agrar. 2012, 44, 197–209, ISSN printed 0370-4661, ISSN online 1853-8665. [Google Scholar]
- De Andrade, J.P.; Giordani, R.B.; Torras-Claveria, L.; Pigni, N.B.; Berkov, S.; Font-Bardia, M.; Calvet, T.; Konrath, E.; Bueno, K.; Sachett, L.G.; et al. The Brazilian Amaryllidaceae as a source of acetylcholinesterase inhibitory alkaloids. Phytochem. Rev. 2016, 15, 147–160. [Google Scholar] [CrossRef]
- Ortiz, J.E.; Berkov, S.; Pigni, N.B.; Theoduloz, C.; Roitman, G.; Tapia, A.; Bastida, J.; Feresin, G.E. Wild Argentinian Amaryllidaceae, a new renewable source of the acetylcholinesterase inhibitor galanthamine and other alkaloids. Molecules 2012, 17, 13473–13482. [Google Scholar] [CrossRef] [PubMed]
- Nicolet, Y.; Lockridge, O.; Masson, P.; Fontecilla-Camps, J.C.; Nachon, F. Crystal structure of human butyrylcholinesterase and of its complexes with substrate and products. J. Biol. Chem. 2003, 278, 41141–41147. [Google Scholar] [CrossRef] [PubMed]
- Cortes, N.; Alvarez, R.; Osorio, E.H.; Alzate, F.; Berkov, S.; Osorio, E. Alkaloid metabolite profiles by GC/MS and acetylcholinesterase inhibitory activities with binding-mode predictions of five Amaryllidaceae plants. J. Pharmaceut. Biomed. Anal. 2015, 102, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Cortes, N.; Sierra, K.; Alzate, F.; Osorio, E.H.; Osorio, E. Alkaloids of Amaryllidaceae as inhibitors of cholinesterases (AChEs and BuChEs): An integrated bioguided study. Phytochem. Anal. 2017, 29, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, P.; Silva, M.; Sammes, P.G.; Watson, W.H. Estudio químico de las Amaryllidaceae chilenas. Nuevos alcaloides de Hippeastrum ananuca. Bol. Soc. Chil. Quim. 1982, 27, 289. [Google Scholar]
- Pacheco, P.; Silva, M.; Steglich, W.; Watson, W.H. Alkaloids of Chilean Amaryllidaceae I hippeastidine and epi-homolycorine two novel alkaloids. Rev. Latinoam. Quim. 1978, 9, 28–32. [Google Scholar]
- Castilhos, T.S.; Giordani, R.; Dutilh, J.; Bastida, J.; de Carvalho, J.E.; Henriques, A.T.; Zuanazzi, J.A.S. Chemical and biological investigation of the alkaloids from Rhodophiala bífida (Herb.) Traub (Amaryllidaceae). In Proceedings of the 24° Reuniao Annual da Sociedade Brasileira de Química, Pocos de Caldas, Brazil, 28–31 May 2001. [Google Scholar]
- Castilhos, T.S.; Giordani, R.; Henriques, A.T.; Menezes, F.S.; Zuanazzi, J.A.S. Availacao in vitro das atividades antiinflamatoria, antioxidante e antimicrobiana do alcaloide montanina. Rev. Bras. Pharmacogn. 2007, 17, 209–214. [Google Scholar] [CrossRef]
- Vieira, P.; Giordani, R.; de Carli, G.; Tasca, T.; Zuanazzi, J. Screening and bioguided fractionation of Amaryllidaceae species with activity against Trichomonas vaginalis. Planta Med. 2010, 76, 470. [Google Scholar] [CrossRef]
- Kulhánková, A.; Cahlíková, L.; Novák, Z.; Macáková, K.; Kuneš, J.; Opletal, L.P. Alkaloids from Zephyranthes robusta Baker and their acetylcholinesterase- and butyrylcholinesterase-inhibitory activity. Chem. Biodivers. 2013, 10, 1120–1127. [Google Scholar] [CrossRef] [PubMed]
- Flora de Chile en su Habitat. Available online: www.floradechile.cl/monocoty/family/amaryllis.htm (accessed on 16 June 2018).
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef] [Green Version]
- López, S.; Bastida, J.; Viladomat, F.; Codina, C. Acetylcholinesterase inhibitory activity of some Amaryllidaceae alkaloids and Narcissus extracts. Life Sci. 2002, 71, 2521–2529. [Google Scholar] [CrossRef]
- De Andrade, J.P.; Pigni, N.B.; Torras-Claveria, L.; Berkov, S.; Codina, C.; Viladomat, F.; Bastida, J. Bioactive alkaloid extracts from Narcissus broussonetii: Mass spectral studies. J. Pharmaceut. Biomed. 2012, 70, 13–25. [Google Scholar] [CrossRef] [PubMed]
- De Andrade, J.P.; Guo, Y.; Font-Bardia, M.; Calvet, T.; Dutilh, J.; Viladomat, F.; Codina, C.; Nair, J.J.; Zuanazzi, J.A.S.; Bastida, J. Crinine-type alkaloids from Hippeastrum aulicum and H. calyptratum. Phytochemistry 2014, 103, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Torras-Claveria, L.; Berkov, S.; Codina, C.; Viladomat, F.; Bastida, J. Metabolomic analysis of bioactive Amaryllidaceae alkaloids of ornamental varieties of Narcissus by GC-MS combined with k-means cluster analysis. Ind. Crop. Prod. 2014, 56, 211–222. [Google Scholar] [CrossRef]
- Tallini, L.R.; de Andrade, J.P.; Kaiser, M.; Viladomat, F.; Nair, J.J.; Zuanazzi, J.A.S.; Bastida, J. Alkaloid constituents of the Amaryllidaceae plant Amaryllis belladonna L. Molecules 2017, 22, 1437. [Google Scholar] [CrossRef] [PubMed]
- Tallini, L.R.; Osorio, E.H.; dos Santos, V.D.; Borges, W.d.S.; Kaiser, M.; Viladomat, F.; Zuanazzi, J.A.S.; Bastida, J. Hippeastrum reticulatum (Amaryllidaceae): Alkaloids profiling, biological activities and molecular docking. Molecules 2017, 22, 2191. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Pigni, N.B.; Zheng, Y.; de Andrade, J.P.; Torras-Claveria, L.; Borges, W.S.; Viladomat, F.; Codina, C.; Bastida, J. Analysis of bioactive Amaryllidaceae alkaloid profiles in Lycoris species by GC-MS. Nat. Prod. Commun. 2014, 9, 1081–1086. [Google Scholar] [PubMed]
- Greenblatt, H.M.; Kryger, G.; Lewis, T.; Silman, I.; Sussman, J.L. Structure of acetylcholinesterase complexed with (−)-galanthamine at 2.3 Å resolution. FEBS Lett. 1999, 463, 321–326. [Google Scholar] [CrossRef] [Green Version]
- Nachon, F.; Carletti, E.; Ronco, C.; Trovaslet, M.; Nicolet, Y.; Jean, L.; Renard, P.-Y. Crystal structures of human cholinesterases in complex with huprine W and tacrine: Elements of specificity for anti-Alzheimer’s drugs targeting acetyl- and butyryl-cholinesterase. Biochem. J. 2013, 453, 393–399. [Google Scholar] [CrossRef] [PubMed]
- ChemCraft. Available online: http://www.chemcraftprog.com/citation.html (accessed on 9 October 2017).
- Adamo, C.; Barone, V. Toward reliable density functional methods without adjustable parameters: The PBE0 model. J. Chem. Phys. 1999, 110, 6158–6170. [Google Scholar] [CrossRef]
- Petersson, G.A.; Bennett, A.; Tensfeldt, T.G.; Al-Laham, M.A.; Shirley, W.A.; Mantzaris, J.; Mantzaris, J. A complete basis set model chemistry. I. The total energies of closed-shell atoms and hydrides of the first-row elements. J. Chem. Phys. 1988, 89, 2193–2218. [Google Scholar] [CrossRef]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Mennucci, B.; Petersson, G.A.; et al. Gaussian 09, Revis. E.01; Gaussian, Inc.: Wallingford, CT, USA, 2013. [Google Scholar]
- Moris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. Autodock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 16, 2785–2791. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, R.; Christensen, M.H. MolDock: A new technique for high-accuracy molecular docking. J. Med. Chem. 2006, 49, 3315–3321. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Names, Abbreviated Collection Place and Reference Letter | % Dry Weight *a | w/w Extraction Yield *b | % Alkaloid Extract *c | AChE IC50 (μg/mL) *d | BuChE IC50 (μg/mL) *d |
|---|---|---|---|---|---|
| Aerial parts | |||||
| R. andicola, SN (B) | 15.46 | 30.63 | 3.16 | 18.16 ± 2.94 | 138.27 ± 6.83 |
| R. andicola, NC (D) | 18.70 | 22.10 | 2.86 | 12.30 ± 0.74 | 43.41 ± 2.64 |
| R. andicola, VL (F) | 20.00 | 17.50 | 2.07 | 74.44 ± 5.53 | >200 |
| R. araucana, M, (H) | 17.70 | 25.30 | 3.78 | *e | *e |
| R. montana, LM (J) | 21.40 | 9.40 | 1.14 | 33.57 ± 2.16 | 16.38 ± 0.78 |
| R. pratensis, AC, RF, nl, sand dunes (L) | 14.84 | 27.08 | 1.93 | 72.59 ± 4.26 | >200 |
| R. pratensis, AC, RF, L (N) | 14.53 | 10.10 | 3.72 | 102.27 ± 6.61 | >200 |
| R. pratensis, AC, RF, nl (P) | 11.68 | 32.39 | 2.87 | 31.97 ± 3.24 | >200 |
| R. pratensis, AC, WF (R) | 10.67 | 16.47 | 1.15 | 8.39 ± 0.27 | >200 |
| R. splendens, LT, (T) | 14.81 | 44.26 | 1.53 | 5.78 ± 0.93 | 16.26 ± 3.34 |
| Bulbs | |||||
| R. andicola, SN (A) | 25.41 | 7.53 | 2.15 | 13.29 ± 1.01 | 45.76 ± 9.72 |
| R. andicola, NC (C) | 28.90 | 6.00 | 2.23 | 7.26 ± 0.16 | 47.38 ± 4.08 |
| R. andicola, VL (E) | 23.50 | 7.40 | 2.37 | 22.77 ± 1.57 | 113.24 ± 2.77 |
| R. araucana, M, (G) | 19.80 | 7.90 | 2.73 | 6.23 ± 0.24 | 45.71 ± 3.51 |
| R. montana, LM, (I) | 28.00 | 3.26 | 1.50 | 18.13 ± 0.51 | 40.05 ± 9.03 |
| R. pratensis, AC, RF, nl, sand dunes (K) | 18.87 | 19.73 | 1.50 | 11.81 ± 0.17 | >200 |
| R. pratensis, AC, RF, L (M) | 20.98 | 11.73 | 2.84 | 44.23 ± 4.08 | >200 |
| R. pratensis, AC, RF, nl (O) | 18.76 | 7.36 | 2.33 | 47.66 ± 1.78 | >200 |
| R. pratensis, AC, WF (Q) | 19.64 | 4.84 | 2.29 | 3.32 ± 0.26 | 52.16 ± 0.57 |
| R. splendens, LT, (S) | 21.35 | 35.05 | 1.60 | 3.62 ± 0.02 | 14.37 ± 1.94 |
| (A) | ||||||||||||||
| Alkaloid | M+ | BP | RI | B | D | F | H | J | L | N | P | R | T | |
| Lycorine-type | 8.6 | 7.5 | 25.1 | 46.0 | 46.9 | 35.5 | 37.3 | 35.4 | 60.8 | 13.1 | ||||
| lycorene (1) | 255 | 254 | 2346.8 | - | - | - | - | 9.4 | 5.1 | 7.2 | T | - | - | |
| anhydrolycorine (2) | 251 | 250 | 2543.1 | - | T | 5.9 | 11.2 | 9.6 | 9.9 | 8.3 | 11.8 | 13.2 | - | |
| kirkine (3) | 253 | 252 | 2588.2 | - | - | - | - | 7.2 | 5.3 | - | 6.0 | 7.2 | - | |
| 11,12-dehydroanhydrolycorine (4) | 249 | 248 | 2646.4 | 8.6 | 7.5 | 13.5 | 8.4 | 15.6 | 8.1 | 21.8 | 7.5 | 9.4 | 7.8 | |
| 1-O-acetylcaranine (5) | 313 | 226 | 2653.1 | - | - | - | - | - | - | - | - | - | - | |
| norpluviine (6) | 273 | 228 | 2683.6 | - | - | - | - | - | - | - | - | 6.2 | - | |
| assoanine (7) | 267 | 266 | 2708.9 | - | T | - | - | - | - | - | - | - | - | |
| 3,4-dihydro 1-acetyllycorine (8) | 331 | 330 | 2723.3 | - | - | - | - | - | - | - | - | - | - | |
| lycorine (9) | 287 | 226 | 2791.7 | - | T | 5.7 | 14.7 | 5.1 | 7.1 | - | 10.1 | 24.8 | 5.3 | |
| dihydrolycorine (10) | 289 | 288 | 2833.9 | - | - | - | - | - | - | - | T | T | - | |
| pseudolycorine (11) | 289 | 228 | 2856.4 | - | - | - | - | - | - | - | - | - | - | |
| sternbergine (12) | 331 | 228 | 2838.8 | - | - | - | 6.4 | - | - | - | T | - | - | |
| methylpseudolycorine (13) | 303 | 242 | 2911.2 | - | - | - | - | - | - | - | - | - | ||
| 1-O-(3′acetoxybutanoyl)lycorine (14) | 415 | 226 | 3248.8 | - | - | - | 5.3 | - | - | - | - | - | - | |
| Haemanthamine/crinine type | - | 11.6 | 53.1 | 34.4 | 14.8 | 17.2 | 21.4 | 38.7 | 35.7 | 48.0 | ||||
| vittatine (15a)/crinine (15b) | 271 | 271 | 2512.4 | - | T | 7.7 | - | - | - | - | 6.2 | 5.6 | - | |
| 8-O-demethylmaritidine (16) | 273 | 273 | 2540.0 | - | - | - | - | - | - | - | T | - | 6.4 | |
| deacetylcantabricine (17) | 275 | 275 | 2573.1 | - | - | - | - | - | - | - | - | - | T | |
| haemanthamine (18a)/crinamine (18b) | 301 | 272 | 2673.4 | - | 11.6 | 45.4 | 34.4 | 7.7 | 17.2 | 21.4 | 32.5 | 30.1 | 25.0 | |
| buphanidrine (19) | 315 | 315 | 2748.3 | - | - | - | - | - | - | - | - | - | - | |
| 11-hydroxyvittatine (20a)/hamayne (20b) | 287 | 258 | 2750.5 | - | - | - | - | - | - | - | - | - | 16.6 | |
| undulatine (21) | 331 | 331 | 2892.5 | - | - | - | - | 7.1 | - | - | - | - | - | |
| Tazettine-type | 60.7 | 59.1 | 95.8 | 22.3 | - | 11.2 | 16.3 | 16.0 | 29.9 | 55.6 | ||||
| deoxytazettine (22) | 315 | 231 | 2575.6 | 6.1 | 5.5 | 6.7 | 5.0 | - | T | 5.2 | - | 5.2 | - | |
| O-methyltazettine (23) | 345 | 261 | 2641.1 | 54.6 | 36.0 | 66.2 | 10.3 | - | 11.2 | 11.1 | 10.1 | 14.9 | 30.7 | |
| tazettine (24) | 331 | 247 | 2685.1 | - | 12.5 | 17.0 | 7.0 | - | - | - | 5.9 | 9.8 | 24.9 | |
| epimacronine (25) | 329 | 245 | 2848.0 | - | 5.1 | 5.9 | T | - | T | - | - | T | T | |
| Homolycorine-type | - | - | 9.1 | - | - | 8.6 | 22.4 | 6.5 | 19.5 | - | ||||
| nerinine (26) | 347 | 109 | 2513.5 | - | - | - | - | - | 8.6 | 22.4 | 6.5 | 19.5 | - | |
| 8-O-demethylhomolycorine (27) | 301 | 109 | 2856.4 | - | - | 9.1 | - | - | - | - | - | - | - | |
| Galanthamine-type | - | 5.1 | 14.1 | 7.1 | 5.8 | - | - | - | - | - | ||||
| galanthamine (28) | 287 | 286 | 2519.9 | - | - | 8.6 | 7.1 | 5.8 | - | - | - | - | - | |
| lycoramine (29) | 289 | 288 | 2544.6 | - | 5.1 | 5.5 | - | - | - | - | - | - | - | |
| Montanine-type | - | - | - | - | 8.1 | 28.3 | 19.7 | 15.2 | 26.7 | - | ||||
| pancratinine C (30) | 287 | 176 | 2623.5 | - | - | - | - | - | - | - | - | - | ||
| montanine (31) | 301 | 301 | 2663.1 | - | - | - | - | 8.1 | 28.3 | 19.7 | 15.2 | 26.7 | - | |
| pancracine (32) | 287 | 287 | 2737.4 | - | - | - | - | - | - | - | - | - | - | |
| Mesembrenone-type | - | - | - | - | - | - | - | - | - | - | ||||
| demethylmesembrenol (33) | 275 | 205 | 2343.2 | - | - | - | - | - | T | - | T | T | - | |
| Narciclasine-type | 12.7 | 11.2 | 13.5 | 5.8 | 6.2 | 5.5 | 44.5 | - | 10.4 | 32.2 | ||||
| trisphaeridine (34) | 223 | 223 | 2322.9 | 12.7 | 5.1 | 6.0 | 5.8 | 6.2 | 5.5 | 39.4 | T | 5.2 | 22.7 | |
| dihydrobicolorine (35) | 239 | 238 | 2366.1 | T | 6.1 | 7.5 | T | - | T | 5.1 | T | 5.2 | 9.5 | |
| Other-type | 5.2 | 32.9 | 62.1 | 8.5 | - | 5.9 | - | 5.7 | 17.7 | 34.5 | ||||
| ismine (36) | 257 | 238 | 2304.6 | - | 7.6 | 17.0 | T | - | T | - | T | 6.0 | 14.3 | |
| galanthindole (37) | 281 | 281 | 2534.8 | 5.2 | 25.3 | 45.1 | 8.5 | - | 5.9 | - | 5.7 | 11.7 | 20.2 | |
| Not identified | 46.0 | 26.8 | 46,1 | 42.0 | 126.9 | 35.9 | 78.5 | 20.6 | 73.4 | 21.8 | ||||
| unknown (ismine-derivate) * | 227 | 226 | 2232.2 | - | - | - | - | - | - | 8.8 | - | - | - | |
| unknown (ismine-derivate) * | 227 | 225 | 2232.2 | - | 5.6 | 6.8 | 5.6 | - | - | 8.9 | - | - | 6.2 | |
| unknown | 269 | 238 | 2258.9 | - | 5.1 | 6.9 | - | - | - | - | - | - | 6.4 | |
| unknown (mesembrenone-type) * | 245 | 175 | 2280.8 | - | - | - | - | 6.0 | - | - | 10.5 | - | ||
| unknown | 269 | 268 | 2285.5 | - | - | - | - | - | - | 9.7 | - | - | - | |
| unknown | 253 | 252 | 2313.1 | 5.9 | T | - | - | 5.9 | - | 13.7 | T | - | - | |
| unknown | 251 | 251 | 2335.2 | - | - | - | - | 6.2 | - | 9.9 | - | - | - | |
| unknown (lycorine-type) * | 257 | 256 | 2379.0 | - | - | - | - | - | - | - | - | - | - | |
| unknown | 253 | 252 | 2405.0 | - | - | - | - | 8.9 | 6.2 | 10.3 | - | 5.5 | - | |
| unknown (lycorine-type) * | 269 | 211 | 2480.5 | - | - | - | T | 28.1 | - | 6.1 | T | 7.1 | - | |
| unknown | 271 | 238 | 2492.7 | - | - | - | - | - | - | - | - | - | - | |
| unknown (homolycorine-type) * | 331 | 109 | 2557.5 | - | - | - | - | - | 5.3 | 5.7 | 7.0 | 33.0 | - | |
| unknown (crinine/haemanthamine-type) * | 329 | 329 | 2564.1 | - | - | - | - | - | 5.3 | - | T | 7.3 | - | |
| unknown | 257 | 225 | 2579.2 | - | - | - | T | - | - | 5.4 | - | - | - | |
| unknown | 299 | 238 | 2584.0 | - | - | - | - | - | - | - | - | - | - | |
| unknown (crinine/haemanthamine-type) * | 315 | 254 | 2616.4 | - | - | - | - | 17.5 | 6.0 | - | 5.7 | - | - | |
| unknown (tazettine-type) * | 315 | 231 | 2616.9 | - | - | - | - | - | - | - | - | - | - | |
| unknown (lycorine-type) * | 329 | 268 | 2646.1 | - | - | - | - | 5.1 | - | - | - | - | - | |
| unknown | 297 | 297 | 2655.7 | - | - | - | - | 5.4 | - | - | - | - | - | |
| unknown (tazettine-type) * | 345 | 261 | 2662.6 | 17.9 | 16.1 | 24.6 | 5.6 | - | - | - | - | - | 9.2 | |
| unknown (crinine/haemanthamine-type) * | 345 | 272 | 2671.8 | 22.2 | - | - | - | - | - | - | - | - | - | |
| unknown (crinine/haemanthamine-type) * | 283 | 283 | 2692.5 | - | - | - | - | 6.9 | - | - | - | - | - | |
| unknown | 303 | 302 | 2703.2 | - | - | - | - | - | 7.1 | - | - | - | - | |
| unknown (crinine/haemanthamine-type) * | 315 | 315 | 2724.2 | - | - | - | 16.8 | - | - | - | - | - | ||
| unknown (lycorine-type) * | 253 | 252 | 2735.3 | - | - | - | - | - | - | - | - | - | - | |
| unknown (homolycorine-type) * | 345 | 109 | 2735.6 | - | - | - | - | 5.9 | - | - | - | - | - | |
| unknown (crinine/haemanthamine-type) * | 347 | 331 | 2795.5 | - | - | - | - | 6.5 | - | - | - | - | - | |
| unknown (crinine/haemanthamine-type) * | 345 | 331 | 2795.7 | - | - | - | 5.6 | - | - | - | - | - | - | |
| unknown (crinine/haemanthamine-type) * | 347 | 331 | 2795.8 | - | - | - | - | - | - | - | - | - | - | |
| unknown (lycorine-type) * | 251 | 250 | 2846.6 | - | - | 7.8 | - | - | - | - | - | - | - | |
| unknown | 335 | 335 | 2860.6 | - | - | - | - | - | - | - | - | - | - | |
| unknown (lycorine-type) * | 345 | 242 | 2868.8 | - | - | - | 5.3 | - | - | - | - | - | - | |
| unknown | 373 | 372 | 2881.9 | - | - | - | - | - | - | - | - | - | - | |
| unknown (lycorine-type) * | 357 | 356 | 2942.1 | - | - | - | - | - | - | - | 7.9 | 10.0 | - | |
| unknown (lycorine-type) * | 279 | 278 | 3016.0 | - | - | - | - | 13.7 | - | - | - | - | - | |
| unknown (galanthamine-type) * | 375 | 330 | 3027.5 | - | - | - | - | - | - | - | - | - | - | |
| unknown (lycorine-type) * | 267 | 266 | 3032.9 | - | - | - | - | - | - | - | - | - | - | |
| unknown (lycorine-type) * | 375 | 374 | 3049.2 | - | - | - | - | - | - | - | - | - | - | |
| unknown (lycorine-type) * | 355 | 226 | 3066.6 | - | - | - | 6.1 | - | - | - | - | - | - | |
| unknown (lycorine-type) * | 373 | 226 | 3161.0 | - | - | - | 13.8 | - | - | - | - | - | - | |
| Total | 133.2 | 154.2 | 311.1 | 166.1 | 208.7 | 148.1 | 240.1 | 138.1 | 274.1 | 205.3 | ||||
| (B) | ||||||||||||||
| Alkaloid | M+ | BP | RI | A | C | E | G | I | K | M | O | Q | S | |
| Lycorine-type | 44.1 | 58.2 | 43.2 | 79.0 | 78.6 | 42.6 | 26.1 | 33.8 | 41.2 | 38.5 | ||||
| lycorene (1) | 255 | 254 | 2346.8 | - | - | - | - | 7.6 | - | T | - | - | - | |
| anhydrolycorine (2) | 251 | 250 | 2543.1 | 18.1 | 15.4 | 25.0 | 27.3 | 24.6 | 9.4 | 10.1 | 13.3 | 14.9 | 7.7 | |
| kirkine (3) | 253 | 252 | 2588.2 | 5.0 | 6.0 | - | 5.1 | T | 6.1 | - | - | - | T | |
| 11,12-dehydroanhydrolycorine (4) | 249 | 248 | 2646.4 | 11.0 | 8.6 | 8.1 | 8.5 | 19.0 | 6.6 | 6.4 | 7.3 | 6.6 | 8.5 | |
| 1-O-acetylcaranine (5) | 313 | 226 | 2653.1 | - | - | - | - | 17.4 | - | - | - | - | - | |
| norpluviine (6) | 273 | 228 | 2683.6 | - | - | - | - | - | - | - | - | - | - | |
| assoanine (7) | 267 | 266 | 2708.9 | - | - | - | T | - | - | - | - | - | - | |
| 3,4-dihydro 1-acetyllycorine (8) | 331 | 330 | 2723.3 | - | - | - | - | - | - | T | - | - | T | |
| lycorine (9) | 287 | 226 | 2791.7 | 10.0 | 21.8 | 10.1 | 27.5 | 10.0 | 15.4 | T | 7.4 | 19.7 | 16.3 | |
| dihydrolycorine (10) | 289 | 288 | 2833.9 | - | - | - | - | - | T | 9.6 | T | T | T | |
| pseudolycorine (11) | 289 | 228 | 2856.4 | - | 6.4 | - | 10.6 | - | 5.1 | - | - | - | 6.0 | |
| sternbergine (12) | 331 | 228 | 2838.8 | - | - | - | - | - | - | - | 5.8 | - | - | |
| methylpseudolycorine (13) | 303 | 242 | 2911.2 | - | - | - | T | - | - | - | - | - | - | |
| 1-O-(3′acetoxybutanoyl)lycorine (14) | 415 | 226 | 3248.8 | - | - | - | - | - | - | - | - | - | - | |
| Haemanthamine/crinine type | 60.0 | 49.5 | 31.7 | 43.7 | 17.4 | 28.6 | 33.8 | 45.5 | 44.9 | 36.9 | ||||
| vittatine (15a)/crinine (15b) | 271 | 271 | 2512.4 | - | 10.8 | - | 7.1 | T | 5.6 | 6.2 | 7.3 | 7.3 | 6.3 | |
| 8-O-demethylmaritidine (16) | 273 | 273 | 2540.0 | - | T | - | - | - | - | - | - | - | 6.0 | |
| deacetylcantabricine (17) | 275 | 275 | 2573.1 | - | - | - | - | - | - | - | - | - | - | |
| haemanthamine (18a)/crinamine (18b) | 301 | 272 | 2673.4 | 60.0 | 31.7 | 31.7 | 36.6 | 5.8 | 23.0 | 27.6 | 38.2 | 31.0 | 16.4 | |
| buphanidrine (19) | 315 | 315 | 2748.3 | - | - | - | - | 5.9 | - | - | - | - | - | |
| 11-hydroxyvittatine (20a)/ hamayne (20b) | 287 | 258 | 2750.5 | - | 7.00 | - | T | - | - | - | - | 6.6 | 8.2 | |
| undulatine (21) | 331 | 331 | 2892.5 | - | - | - | - | 5.7 | - | - | - | - | - | |
| Tazettine-type | 74.8 | 56.1 | 85.9 | 33.1 | - | 21.8 | 18.0 | 8.9 | 17.9 | 21.9 | ||||
| deoxytazettine (22) | 315 | 231 | 2575.6 | 8.3 | 10.7 | 13.1 | 9.6 | - | - | - | - | - | T | |
| O-methyltazettine (23) | 345 | 261 | 2641.1 | 58.7 | 23.0 | 61.7 | 14.8 | - | 10.9 | 12.7 | 8.9 | 9.9 | 14.3 | |
| tazettine (24) | 331 | 247 | 2685.1 | 7.8 | 22.4 | 11.1 | 8.7 | - | 10.9 | 5.3 | T | 8.0 | 7.6 | |
| epimacronine (25) | 329 | 245 | 2848.0 | - | T | T | T | - | - | - | - | - | - | |
| Homolycorine-type | 9.4 | - | 10.6 | - | - | 13.9 | 14.1 | 10.1 | 28.5 | 9.8 | ||||
| nerinine (26) | 347 | 109 | 2513.5 | - | - | - | - | - | 13.9 | 14.1 | 10.1 | 21.1 | - | |
| 8-O-demethylhomolycorine (27) | 301 | 109 | 2856.4 | 9.4 | - | 10.6 | - | - | - | - | - | 7.4 | 9.8 | |
| Galanthamine-type | - | 19.0 | 11.9 | 12.3 | - | - | - | - | - | - | ||||
| galanthamine (28) | 287 | 286 | 2519.9 | - | 5.1 | 6.8 | 5.4 | T | - | - | - | - | - | |
| lycoramine (29) | 289 | 288 | 2544.6 | - | 13.9 | 5.1 | 6.9 | - | - | - | - | - | - | |
| Montanine-type | - | - | - | 5.2 | 7.8 | 41.7 | 23.1 | 12.5 | 24.7 | 6.4 | ||||
| pancratinine C (30) | 287 | 176 | 2623.5 | - | - | - | - | T | 5.3 | T | - | - | - | |
| montanine (31) | 301 | 301 | 2663.1 | - | - | - | 5.2 | 7.8 | 29.8 | 23.1 | 12.5 | 24.7 | - | |
| pancracine (32) | 287 | 287 | 2737.4 | - | - | - | - | - | 6.6 | - | - | - | 6.4 | |
| Mesembrenone-type | - | - | - | - | - | 7.2 | 5.2 | - | 5.1 | - | ||||
| demethylmesembrenol (33) | 275 | 205 | 2343.2 | - | - | - | - | - | 7.2 | 5.2 | T | 5.1 | - | |
| Narciclasine-type | 12.9 | 12.7 | 13.3 | 5.8 | 5.7 | - | - | - | - | 5.1 | ||||
| trisphaeridine (34) | 223 | 223 | 2322.9 | 5.5 | 6.3 | 5.3 | T | 5.7 | T | T | T | T | 5.1 | |
| dihydrobicolorine (35) | 239 | 238 | 2366.1 | 7.4 | 6.4 | 8.0 | 5.8 | T | T | T | T | T | T | |
| Other-type | 19.2 | 35.6 | 28.2 | 21.8 | - | 7.9 | 7.8 | 5.3 | 8.0 | 7.3 | ||||
| ismine (36) | 257 | 238 | 2304.6 | 6.6 | 9.9 | 9.5 | 6.6 | - | T | T | T | T | T | |
| galanthindole (37) | 281 | 281 | 2534.8 | 12.6 | 25.7 | 18.7 | 15.2 | - | 7.9 | 7.8 | 5.3 | 8.0 | 7.3 | |
| Not identified | 29.9 | 41.4 | 41.2 | 79.8 | 76.8 | 92.6 | 50.5 | 32.0 | 76.0 | 36.0 | ||||
| unknown (ismine-derivate) * | 227 | 226 | 2232.2 | - | - | - | - | - | - | - | - | - | - | |
| unknown (ismine-derivate) * | 227 | 225 | 2232.2 | - | T | 6.2 | 5.1 | - | - | - | T | - | - | |
| unknown | 269 | 238 | 2258.9 | - | T | - | - | - | - | - | - | - | - | |
| unknown (mesembrenone-type) * | 245 | 175 | 2280.8 | - | - | - | - | - | 15.3 | 11.8 | 6.1 | 11.4 | - | |
| unknown | 269 | 268 | 2285.5 | - | - | - | - | - | - | - | - | - | - | |
| unknown | 253 | 252 | 2313.1 | 5.4 | - | - | - | 6.7 | - | - | 5.1 | - | - | |
| unknown | 251 | 251 | 2335.2 | - | - | - | - | 6.2 | - | - | - | - | - | |
| unknown (lycorine-type) * | 257 | 256 | 2379.0 | - | - | - | 8.0 | - | - | - | - | - | - | |
| unknown | 253 | 252 | 2405.0 | - | 6.4 | - | 5.1 | 6.9 | 8.3 | 8.0 | 6.0 | 5.4 | 5.5 | |
| unknown (lycorine-type) * | 269 | 211 | 2480.5 | - | - | - | T | 34.5 | 5.3 | 5.1 | - | - | - | |
| unknown | 271 | 238 | 2492.7 | 5.7 | - | - | - | - | - | - | - | - | ||
| unknown (homolycorine-type) * | 331 | 109 | 2557.5 | - | - | - | - | - | 19.7 | 18.5 | 8.7 | 46.7 | - | |
| unknown (crinine/haemanthamine-type) * | 329 | 329 | 2564.1 | - | - | - | - | 6.2 | 7.1 | 6.1 | 7.2 | - | ||
| unknown | 257 | 225 | 2579.2 | - | - | - | - | - | - | - | - | - | - | |
| unknown | 299 | 238 | 2584.0 | - | - | T | - | - | - | - | - | - | - | |
| unknown (crinine/haemanthamine-type) * | 315 | 254 | 2616.4 | - | - | - | - | 5.4 | 8.6 | - | - | - | - | |
| unknown (tazettine-type) * | 315 | 231 | 2616.9 | - | - | 5.2 | - | - | - | - | - | - | - | |
| unknown (lycorine-type) * | 329 | 268 | 2646.1 | - | - | - | - | - | 7.9 | - | - | 5.3 | - | |
| unknown | 297 | 297 | 2655.7 | - | - | - | - | - | - | - | - | - | - | |
| unknown (tazettine-type) * | 345 | 261 | 2662.6 | 18.8 | 11.2 | 23.0 | - | - | - | - | - | - | 7.3 | |
| unknown (crinine/haemanthamine-type) * | 345 | 272 | 2671.8 | - | - | - | - | - | - | - | - | - | - | |
| unknown (crinine/haemanthamine-type) * | 283 | 283 | 2692.5 | - | - | - | - | - | - | - | - | - | - | |
| unknown | 303 | 302 | 2703.2 | - | - | - | - | - | 21.3 | - | - | - | - | |
| unknown (crinine/haemanthamine-type) * | 315 | 315 | 2724.2 | - | - | - | - | - | - | - | - | - | - | |
| unknown (lycorine-type) * | 253 | 252 | 2735.3 | - | - | - | 17.1 | - | - | - | - | - | - | |
| unknown (homolycorine-type) * | 345 | 109 | 2735.6 | - | - | - | - | - | - | - | - | - | - | |
| unknown (crinine/haemanthamine-type) * | 347 | 331 | 2795.5 | - | - | - | - | - | - | - | - | - | - | |
| unknown (crinine/haemanthamine-type) * | 345 | 331 | 2795.7 | - | - | - | - | - | - | - | - | - | - | |
| unknown (crinine/haemanthamine-type) * | 347 | 331 | 2795.8 | - | - | - | - | 6.9 | - | - | - | - | - | |
| unknown (lycorine-type) * | 251 | 250 | 2846.6 | - | - | 6.8 | 9.9 | - | - | - | - | - | - | |
| unknown | 335 | 335 | 2860.6 | - | 7.6 | - | - | - | - | - | - | - | - | |
| unknown (lycorine-type) * | 345 | 242 | 2868.8 | - | - | - | - | - | - | - | - | - | - | |
| unknown | 373 | 372 | 2881.9 | - | - | - | - | - | - | - | - | - | 7.8 | |
| unknown (lycorine-type) * | 357 | 356 | 2942.1 | - | - | - | - | - | - | - | - | - | 7.0 | |
| unknown (lycorine-type) * | 279 | 278 | 3016.0 | - | - | - | - | 10.2 | - | - | - | - | - | |
| unknown (galanthamine-type) * | 375 | 330 | 3027.5 | - | 7.0 | - | - | - | - | - | - | - | - | |
| unknown (lycorine-type) * | 267 | 266 | 3032.9 | - | 9.2 | - | 7.3 | - | - | - | - | - | - | |
| unknown (lycorine-type) * | 375 | 374 | 3049.2 | - | - | - | - | - | - | - | - | - | 8.4 | |
| unknown (lycorine-type) * | 355 | 226 | 3066.6 | - | - | - | 5.8 | - | - | - | - | - | - | |
| unknown (lycorine-type) * | 373 | 226 | 3161.0 | - | - | - | 21.5 | - | - | - | - | - | - | |
| Total | 250.3 | 272.5 | 265.9 | 280.4 | 186.0 | 256.3 | 178.6 | 148.1 | 246.3 | 161.9 | ||||
| Alkaloid | AChE | BuChE |
|---|---|---|
| 11-hydroxyvittatine (20a) | 122.17 ± 22.03 | >200 |
| lycorine (9) | 101.70 ± 23.79 | >200 |
| 8-O-demethylmaritidine (16) | 113.21 ± 8.21 | 127.87 ± 2.45 |
| hamayne (20b) | 135.09 ± 15.33 | 48.40 ± 1.13 |
| deacetylcantabricine (17) | >200 | >200 |
| haemanthamine (18a) | 184.68 ± 11.58 | >200 |
| galanthamine (28) | 0.48 ± 0.07 | 3.70 ± 0.24 |
| Alkaloid | AChE *a | BuChE *b |
|---|---|---|
| 11-hydroxyvittatine (20a) | −8.43 | −9.03 |
| lycorine (9) | −8.82 *c | −8.94 |
| 8-O-demethylmaritidine (16) | −8.74 *d | −8.93 |
| hamayne (20b) | −8.28 | −8.54 |
| deacetylcantabricine (17) | −7.90 | −8.43 |
| haemanthamine (18a) | −8.80 | −8.34 |
| galanthamine (28) | −9.55 *c | −8.23 *c |
| epimacronine (25) | −9.36 *c | −7.63 |
| tazettine (24) | −8.66 *c | −7.87 |
| O-methyltazettine (23) | −8.54 | −7.87 |
| 11,12-dehydroanhydrolycorine (4) | −8.41 | −7.44 |
| galanthindole (37) | −7.81 | −7.41 |
| trisphaeridine (34) | −7.38 | −7.27 |
| dihydrobicolorine (35) | −7.33 | −7.38 |
| ismine (36) | −6.78 | −7.08 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tallini, L.R.; Bastida, J.; Cortes, N.; Osorio, E.H.; Theoduloz, C.; Schmeda-Hirschmann, G. Cholinesterase Inhibition Activity, Alkaloid Profiling and Molecular Docking of Chilean Rhodophiala (Amaryllidaceae). Molecules 2018, 23, 1532. https://doi.org/10.3390/molecules23071532
Tallini LR, Bastida J, Cortes N, Osorio EH, Theoduloz C, Schmeda-Hirschmann G. Cholinesterase Inhibition Activity, Alkaloid Profiling and Molecular Docking of Chilean Rhodophiala (Amaryllidaceae). Molecules. 2018; 23(7):1532. https://doi.org/10.3390/molecules23071532
Chicago/Turabian StyleTallini, Luciana R., Jaume Bastida, Natalie Cortes, Edison H. Osorio, Cristina Theoduloz, and Guillermo Schmeda-Hirschmann. 2018. "Cholinesterase Inhibition Activity, Alkaloid Profiling and Molecular Docking of Chilean Rhodophiala (Amaryllidaceae)" Molecules 23, no. 7: 1532. https://doi.org/10.3390/molecules23071532