Melatonin and Its Effects on Plant Systems
by
 , , , , ,
, , , , ,
Rahat Sharif
1, 
Chen Xie
1 ,
,
Haiqiang Zhang
1,
Marino B. Arnao
2,
Muhammad Ali
1 ,
,
Qasid Ali
3,
Izhar Muhammad
4,
Abdullah Shalmani
4,
Muhammad Azher Nawaz
5,
Peng Chen
6,* and
Yuhong Li
1,* 1
College of Horticulture, Northwest A&F University, Yangling 712100, China
2
Department of Plant Biology (Plant Physiology), Faculty of Biology, University of Murcia, Campus de Espinardo, 30100 Murcia, Spain
3
Department of Horticulture, Faculty of Agriculture, Akdeniz University, 07059 Antalya, Turkey
4
State Key Laboratory of Crop Stress Biology in Arid Areas, College of Life Sciences, Northwest A&F University, Yangling 712100, China
5
Department of Horticulture, University college of Agriculture, University of Sargodha, Sargodha 40100, Pakistan
6
College of Life Science, Northwest A&F University, Yangling 712100, China
*
Authors to whom correspondence should be addressed.
Molecules 2018, 23(9), 2352; https://doi.org/10.3390/molecules23092352
Submission received: 13 June 2018
/
Revised: 11 July 2018
/
Accepted: 20 July 2018
/
Published: 14 September 2018
(This article belongs to the Special Issue Melatonin as an Antioxidant and a Functionally Pleiotropic Molecule: Synthesis, Metabolism and Activities in Organisms)
Abstract
:Melatonin (N-acetyl-5-methoxytryptamine) is a nontoxic biological molecule produced in a pineal gland of animals and different tissues of plants. It is an important secondary messenger molecule, playing a vital role in coping with various abiotic and biotic stresses. Melatonin serves as an antioxidant in postharvest technology and enhances the postharvest life of fruits and vegetables. The application of exogenous melatonin alleviated reactive oxygen species and cell damage induced by abiotic and biotic stresses by means of repairing mitochondria. Additionally, the regulation of stress-specific genes and the activation of pathogenesis-related protein and antioxidant enzymes genes under biotic and abiotic stress makes it a more versatile molecule. Besides that, the crosstalk with other phytohormones makes inroads to utilize melatonin against non-testified stress conditions, such as viruses and nematodes. Furthermore, different strategies have been discussed to induce endogenous melatonin activity in order to sustain a plant system. Our review highlighted the diverse roles of melatonin in a plant system, which could be useful in enhancing the environmental friendly crop production and ensure food safety.
Keywords:
melatonin; abiotic stress; biotic stress; antioxidants; gene expression; postharvest; mitochondria1. Introduction
Melatonin was discovered in the bovine pineal gland and called as vertebrate pineal secretory molecule [1,2]. The pineal gland is an organ present in animals’ bodies, responsible for the production of melatonin in order to control the behavior of the body toward changing photoperiod and also serve as a neuronal protective antioxidant [3]. However, in plants, the existence of melatonin has been reported in more than 20 dicotyledonous and monocotyledonous plant families [4,5]. The name melatonin was given to this biomolecule after Lerner et al. [6] stated that an indole molecule is responsible for causing skin lighting in the frog. In addition to that, it is the most important factor for controlling the circadian cycle in different vertebrates; the secretion of melatonin reaches to the highest level during the night time, which makes it the peak signaling molecule of darkness [2]. Moreover, melatonin is an important antioxidant that can be taken in diet and also the body produces it endogenously, though its production degrades gradually with increasing age [2,7,8]. In plants, the role of melatonin has been extensively studied [9,10,11].
Since the discovery of melatonin in 1965, about 34,000 research materials regarding melatonin are available on Scopus database; this highlights the importance of this molecule in that it has been studied extensively. Because of its significant effects on plant systems, it attracts scientists and young researchers from the diverse field of plant sciences [12,13,14,15]. It is considered as a central indoleamine neurotransmitter, largely involved in the diverse biological process and accepted as an important plant metabolite [16,17]. Additionally, melatonin has been reported for its involvement in improving seed germination, fruit ripening, photosynthesis, biomass production, circadian rhythm, redox network, membrane integrity, root development, leaf senescence, osmoregulation, abiotic stress (salt, drought, cold, heat, oxidative, heavy metals) [18,19,20,21,22,23,24,25,26]. Moreover, it has been reported to play a beneficial role in the protection of plants against biotic stresses [14]. Furthermore, melatonin induces gene expression which helps the plant to cope with biotic and abiotic stresses [27]. Therefore, it could be of great importance to utilize melatonin as a bio-stimulator for sustainable crop production without affecting the external environment.
The purpose of this review is to highlight the various aspects of melatonin from the plethora of research available over its role in protecting plants from abiotic, biotic and post-harvest stresses. Additionally, the contribution of melatonin in regulating gene expression has been presented here. The melatonin defense mechanism has also been presented in this review with illustration and diagrammatic sketch (Figure 1). Furthermore, we discussed the melatonin concentration in different plants as well as the strategies to improve the endogenous melatonin content to restore the defense in plants.
2. The Occurrence of Melatonin in Plants
Melatonin is an ancient molecule derived from cyanobacteria which was introduced to animals and plants through evolution [28]. Melatonin is an indolic compound derived from tryptophan and it was found during 1995 in plants. It is found in almost all the plant species, varying in amount according to the plant tissues in light depending manner [29,30]. Though the level of melatonin is higher in aromatic plants and leaves than that of seeds [29]. In higher plants, melatonin was firstly discovered in 1993 in the Convolvulaceae ivy morning glory (Pharbitis nil L., syn. Ipomoeanil L.) and in tomato fruits (Solanum lycopersicum L.), however, due to some unknown reasons, the results were published two years later in 1995 [31,32]. Moreover, the majority of the plants reported to contain melatonin contents are from family Rosaceae, Vitaceae, Poaceae, Apiaceae and Brassicaceae, however, plants from other species have also exhibited higher endogenous melatonin contents. During the last decade, several researchers revealed that the concentration of melatonin also differs between the varieties of the same species, depending on location, growth stage, organ and harvest timing [33,34,35]. Endogenous melatonin was reported to play an important role in the regulation of plant growth attributes in various species [36]. For that reason, several detection methods and assays have been reported for the determination of melatonin content in plant samples that includes radioimmunoassay (RIA), enzyme-linked immunosorbent assay (ELISA), gas chromatography-mass spectrometry (GC-MS), and high-performance liquid chromatography (HPLC) with electrochemical detection (HPLC-ECD), fluorescence detection (HPLC-FD), or high-performance liquid chromatography-mass spectrometry HPLC-MS [37]. However, RIA has been reported to be less reliable in detecting melatonin content compared with other methods [13]. Therefore, we enlisted the reported melatonin contents and its detection methods in some of the important agronomic and horticultural plants (Table 1). However, these values could be changed as melatonin production is dependent on the day length and also seasonal changes. For example, its production is low during the daytime and high during the night time [38,39]. Thus, the values stated in Table 1 are uncertain unless the circadian, seasonal and plant age variations are known. We believe this information could be helpful for future research programs regarding the detection of endogenous melatonin level.
3. Role of Melatonin in Regulating Plant Growth and Physiology
The primary functions of melatonin are as an antioxidant because it is soluble in water and fats and moves freely across the body to any aqueous section [28,51]. Though it has been reported that melatonin improves the overall growth of plants [42,52]. It enhances the coleoptile length of canary grass, barley and wheat [42]. The melatonin-treated maize seeds resulted in better seed vigor and quality and improved seed storage proteins [53]. According to another report, a coating of soybean seed with melatonin significantly improved the leaf growth, plant height, number of pod plants−1 and number of seeds per pod [54]. A similar role of melatonin was also observed in etiolated Lupinus albus L., where it was found to be responsible for the promotion of vegetative growth, and regenerations of lateral and adventitious roots [55,56]. While in cucumber plants, an increase in the seedling growth, improvement in the nutrient uptake efficiency and enhancement in nitrogen metabolism were perceived after treatment with melatonin, particularly under salt stress conditions [57]. Moreover, it was reported that melatonin treatment improved photosynthetic activity, enhanced redox homeostasis, regulated root growth and development, and seminal root elongation in barley, wheat, sweet cherry and rice [47,58,59,60,61,62]. The plant hormones such as auxin, ethylene, cytokinin, gibberellins, IAA (indole 3-acetic acid) and brassinosteroids are extensively involved in regulating plant growth and development [63]. The effects of these plant hormones can be regulated with the application of exogenous melatonin application [64]. Among them, IAA shared similarities in structure and functions [42,65]. In line with that, exogenous treatment of melatonin enhances the production of IAA [66]. While on the other hand both melatonin and IAA work in the combined and similar fashion as they were reported for enhancing root morphogenesis [52]. In brassica juncea plant, exogenously applied melatonin enhanced the IAA level, which further resulted in better root activity [67]. Whereas it influenced the root organogenesis positively in Mimosa pudica L. [68]. Therefore, it is assumed that melatonin influenced signal transduction and also had a role in regulating plant physiological and biological processes. To sum up, melatonin could be considered as a biological plant growth regulator to improve the production capacity of a plant.
4. Melatonin Effect on Postharvest Produce
The shelf-life and quality of postharvest produce decline due to the deterioration. For this reason, many treatments have been implemented to maintain the quality and shelf life of postharvest fruits and vegetables [69,70,71,72]. Usually, the produce is stored in a cold environment which induces oxidative stress by elevating the production of ROS; this is the main drawback of cold storage [73]. However, treatment with melatonin alleviates the ROS activity and increases the antioxidant enzymes production [70]. In other cases, the application of exogenous melatonin triggered the endogenous melatonin biosynthetic activity via the antagonistic crosstalk with calcium, preventing the product from postharvest deterioration [74]. Additionally, the postharvest quality of horticultural produce is mainly dependent on the preharvest factors as it cannot be increased after harvesting but can only be maintained [75]. In line with that, the tomato seeds fertigated with melatonin had not only increased their yield but also kept the postharvest quality by exhibiting an increase of vitamin C, lycopene and calcium contents. The treated plants also recorded for more soluble solids and P content than that of control [76]. In another study, the exogenous application of melatonin on the clusters of grapes attached to the vine had altered metabolism of polyphenol, carbohydrate biosynthesis and more importantly ethylene signaling in berries of grapes. The restricted ethylene production resulted in better antioxidant activity [74], which is an important factor for maintaining postharvest quality. Moreover, melatonin regulates salicylic acid, jasmonic acid, nitric oxide and ethylene which collectively generate the resistance against diseases in a very familiar action [64]. The cooperative or antagonistic approach of ethylene and jasmonate is mainly dependent on the interaction of their downstream signaling pathway [77]. Jasmonic acid encourages the synthesis of lycopene in tomato independently to ethylene and exogenously applied ethylene is widely used to trigger and initiate ripening in climacteric fruits [78,79]. Correspondingly, ethylene does not only affect the biochemical structure but also increases the respiration rate of fruit and vegetables [80]. Likewise, the exogenously applied melatonin influenced the ethylene biosynthesis pathway and conferred better aroma, color, sugar and overall postharvest quality of tomato [81]. The research provides a good base for utilizing melatonin in keeping the postharvest quality of produces. Both of these hormones regulated by melatonin play an important role in defining the postharvest status of produce by means of their possible involvement in providing resistance against postharvest diseases and deterioration. Still, not a great deal of research material is available on melatonin postharvest application. However, melatonin may be considered as a potential substance to reduce the percentage of postharvest losses and enhance the shelf life of postharvest produce. According to a recent report, the silencing of fruit shelf-life regulator (SIFR) gene has been reported for controlling the postharvest ripeness in tomato and also extended the fruit shelf life by inhibiting the ethylene production [82]. For that reason, it will be interesting to see how exogenous melatonin affects the postharvest maturity by regulating the expression level of SIFR gene. Furthermore, Table 2 represents the reported studies on melatonin application over postharvest products.
5. Role of Melatonin in Mitigating Abiotic Stresses
In recent times, melatonin as a biostimulant and plant growth regulator attracts the interest of plant biologists [14]. For instance, it provides physiological and molecular resistance against many abiotic stresses by means of its involvement in regulating stress signaling [92,93]. Additionally, its beneficial effect on photosynthesis and other growth-related factors amongst different crops under the diverse abiotic stresses is another promising aspect of melatonin application [94,95]. Exogenous melatonin significantly induced the level of endogenous Abscisic acid (ABA) and Gibberellic acid (GA) in cucumber seedling under the saline condition, due to which the resistance against salinity was improved [96]. While in plants affected by heat stress, the level of cytokinin (CK) was degraded gradually. Though, induction in the level of CK biosynthesis was observed after the plants were treated with exogenous melatonin. The study further reported that the resistance against heat stress was perceived in the melatonin-treated plants due to enhanced CK level [97]. In short, the main action of mechanism is the improvement of the antioxidant defense system and enhancing photosynthetic activity (Table 3). Moreover, melatonin has been described by many scientists to significantly influence the overall plant growth against abiotic stresses [87,97,98,99] with minimum effects on the surrounding environment.
6. Melatonin Role in Suppressing Biotic Stresses
To the best of our knowledge, the first study conducted on melatonin’s ability to increase resistance to biotic stress in plants was reported by Yin et al. [129]. In their study, they successfully used melatonin to mimic the harmful effects of Diplocarpon mali in apple tree through the root irrigation method. In another study, the SNAT mutant line in Arabidopsis suffered from avirulent pathogen Pseudomonas syringae pv due to the reduced induction capacity of defense genes (PR1, ICS1, and PDF 1.2). However, the induction was restored with the application of exogenous melatonin, confirming the role of melatonin in suppressing biotic stress [20]. Similarly, in apple juice, the melatonin showed excellent anti-microbes activity by reducing the percentage to 19% compared with control [83]. Moreover, the application of exogenous melatonin on the Arabidopsis plant augmented the level of endogenous melatonin and nitric oxide (NO) that was lethal against the pathogen Pseudomonas syringe pv. tomato (Pst) DC3000 [24]. So far it is known that melatonin expresses the activity of chitinase genes [129], which is an important factor in restricting the lesion expansion and inhibiting the growth of pathogen [69]. However, the induction in endogenous melatonin activity also plays an important role in sustaining the defense system [24]. More generally, that melatonin induces resistance against biotic stress is a collective action of endogenous hormonal [20], antioxidant enzymes activities and expression of PR and Chitinase genes.
Furthermore, the melatonin-directed regulation of other phytohormones could provide resistance against biotic stresses, which is yet to be examined in plant [64]. For example, ethylene enhances the infection capacity and symptoms development as observed for cucumber inoculated with Cucumber mosaic virus (CMV) [130,131]. However, treatment with exogenous melatonin suppresses the ethylene activity [73] and could help in sustaining the plant defense system against that particular virus. A similar case is with other phytohormones as they are highly involved in keeping the balance of the plant defense system [63,132,133] but virus infection inanimates the plant hormones system, which helps them to replicate quickly [134]. Therefore, it could be of great importance to unravel the role of melatonin mediated defense response against plant viruses and other biotic stress factors. The reported studies in Table 4 demonstrated the potential role of melatonin application against biotic stress. However, more research is needed on the melatonin application against plant viruses, nematodes, and insects.
7. Regulation of Gene Expression
Gene expression is defined as a biological process which changes according to environmental stimuli. Sometimes these environmental stimuli induce positive and negative gene expression which further delimits the biological processes and production capacity of plants. Recently, various biological gene regulators have been reported [69,137,138]. Here we discussed the melatonin role in regulating and expressing gene activities in different plants under various stress conditions. Additionally, an Arabidopsis mutant, SNAT, was subjected to cold stress, whose anthocyanin producing ability was quite low compared to that of wild-type. However, after the application of exogenous melatonin, a restoration process was upheld due to the up-regulation of anthocyanin biosynthesis genes, which confirmed that the melatonin improves plant growth via the induction of anthocyanin activities [27]. In another report, cucumber seeds primed with melatonin have been employed to the RNA-seq approach. The study reported that 121 and 196 genes were up and down-regulated respectively. Among them, genes responsible for carbohydrate metabolism, cell wall synthesis, and lateral root formation were reported for exhibiting mix expression pattern [139]. The study made inroads for further functional characterization of the genes involved in lateral root formation, other biological processes and highlighted the all-important role of melatonin involvement in gene expression and regulation. Besides that, a stability analysis of reference genes was performed for the anti-cancerous medicinal plant Catharanthus roseus under exogenous melatonin treatment. The study confirmed that EXP and EXPR were the most stable genes under melatonin treatment, which is important for the future research in order to achieve an accurate expression pattern [140]. Melatonin is also involved in regulating stress-specific genes. For example, the Arabidopsis plant under iron deficient condition restored its tolerance to iron deficiency by regulating FIT1, FRO2, and IRT1 genes after melatonin treatment [141]. The study confirmed that melatonin can increase the tolerance of the plant to iron deficiency by upregulating the iron stress-specific genes. While in Apple, the exogenously applied melatonin delayed leaf senescence by suppressing the chlorophyll degradation genes namely senescence-associated gene 12 (SAG12) and AUXIN RESISTANT 3 (AXR3)/INDOLE-3-ACETIC ACID INDUCIBLE 17 (IAA17) [122]. The same mechanism could be used for other stresses as well, though no study has stated it so far. In another report, the melatonin significantly upregulated the expression of CmSOD, CmPOD, and CmCAT and MYB, bHLH, WD40 genes in melon and cabbage under cold and oxidative stress respectively [27,142]. The restoration of antioxidant enzymes activity is at the center of this induced resistance by these several expressed genes reported in Table 5. Moreover, there is a significant amount of study available on melatonin-induced gene expression, however, no study has examined the role of melatonin in regulating the MLO clade V genes, which is responsible for bringing susceptibility to powdery and downy mildew disease in different plants [143,144]. It could be worth studying the effect of melatonin on these important economic fungal diseases and also the genes responsible for their induction, as melatonin does show the potential of inducing resistance against fungal diseases [14,136]. Furthermore, gene expression in different plant species regulated by melatonin is listed in Table 5.
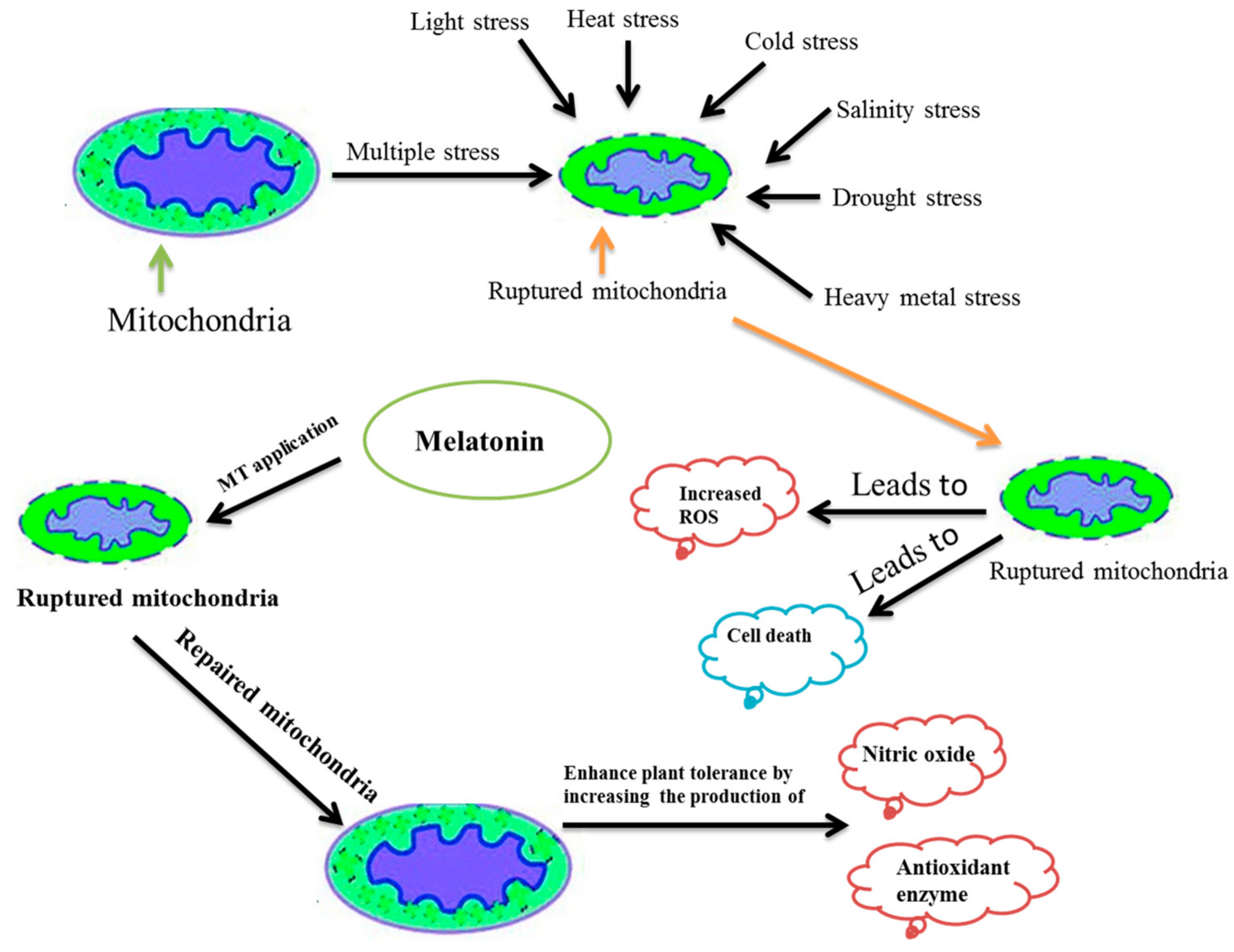
8. Melatonin Defense Mechanism
The defense mechanism of melatonin continues to puzzle researchers as no definite defense mechanism has been proposed so far. Nevertheless, there are many suggested mechanisms of action. For example, the ability to scavenge H2O2 and the induction of antioxidant enzymes activities by melatonin helps to recover plants from abiotic stresses [112,116,147]. In another report, the melatonin was proposed to up-regulate the expression of heat shock protein (HSP) to mitigate the high-temperature stress [92]. While for biotic stress, the melatonin was anticipated for activating the NO and salicylic acid (SA) mediated defense signaling pathway by expressing the PR-protein (pathogenesis-related protein) immediately [24,148]. Mitochondria are the main powerhouse for energy production through aerobic respiration and play a key role in plant growth and development [149]. Additionally, mitochondria and chloroplast are referred to as the original site of melatonin synthesis in plants [28]. In another study, mitochondria were pinpointed as a major generation site for NO and ROS [150,151] and could be important in playing a key role in mitigating various stresses via NO accumulation and ROS regulation [151,152,153]. Though, the mitochondria can be damaged due to the over-production of ROS under environmental stresses [151]. However, melatonin was reported to recover the damaged mitochondria [154]. Therefore, from the recent research reported, we have proposed a new model of melatonin defense mechanism (Figure 1).
9. Approaches to Inducing Endogenous Melatonin Level
9.1. Transgenic Approaches to Inducing Endogenous Melatonin Level
The enzymes responsible for regulating the melatonin biosynthesis pathway has been successfully overexpressed in various crops and showed good results by boosting the level of endogenous melatonin. These enzymes include tryptophan decarboxylase (TDC), tryptamine5-hydroxylase, arylalkylamine N-acetyltransferase (AANAT)/serotonin N-acetyltransferase, and N-acetylserotonin methyltransferase/hydroxyindole-O-methyltransferase (HIOMT) [11,26,155,156,157,158,159,160]. In line with this, Apple MZASMT1 overexpression in Arabidopsis increased the production of melatonin and enhanced the resistance against drought stress by lowering the activity of ROS [161]. In another report, the tomato lines overexpressing the N-acetylserotonin methyltransferase (ASMT) gene improved the level of endogenous melatonin. Additionally, it increased the cellular protection through the generation of heat shock protein (HSP) and activation of autophagy to refold denatured proteins that triggered heat resistance [92]. Moreover, the rice chloroplast caffeic acid O-methyltransferase COMT transgenic lines produced a higher level of melatonin by regulating the 5-methoxytryptamine (5-MT) pathway, which further resulted in improved seedling growth under continuous lighted environment [162]. Similarly, in tomato, the upregulation in COMT1 transcription factor occurred in the plants due to the overexpression of the HsfA1a gene. Additionally, the induction of melatonin biosynthesis and resistance against cadmium stress was also observed in the overexpressed plants [163]. The overexpression of tryptophan decarboxylase 2 (MeTDC2)-interacting proteins, N-acetylserotonin O-methyltransferase 2 (MeASMT2) interacting proteins, and N-acetylserotonin O-methyltransferase 3 (MeASMT3) produced more melatonin than any other enzyme in cassava protoplast. Furthermore, the co-overexpression of these three enzymes with MeWRKY20/75 activated the MeWRKY20 and MeWRKY75 W-box transcriptional activities along with the positive effect on endogenous melatonin level [36]. Taken together, these findings suggested that the transgenic induction of the endogenous melatonin level could be a useful and reliable approach to understand the mechanism of this important signaling molecule in a plant system.
9.2. Some Other Strategies to Induce Endogenous Melatonin Level
Apart from the transgenic approach, there are other ways to induce the endogenous melatonin level in plants. An Arabidopsis melatonin mutant was unable to tackle the avirulent pathogen due to the decrease in melatonin level. However, exogenous melatonin recovers the plant defense mechanism and restores resistance against a pathogen by recovering the endogenous melatonin level [20]. In another study, the endophytic bacterium Pseudomonas fluorescens RG11 strain was used successfully to induce endogenous melatonin level in grapes. The inoculated grape plants showed resistance to salt stress by decreasing the reactive oxygen species burst and cell damage [164]. Similarly, the Bacillus amyloliquefaciens SB-9 endophytic bacterium strain from grapevine root promotes the endogenous melatonin production and also ameliorates the adverse effect of salt and drought stress via scavenging H2O2 activities [165]. Altogether, these environmentally friendly approaches of inducing endogenous melatonin can be utilized against biotic and abiotic stresses in agronomic and horticultural crops.
10. Conclusions
The plethora of research available about melatonin proves that it is an indispensable signaling molecule. The plant produces melatonin endogenously and research showed that it is highly important in maintaining plant growth and development. Additionally, the mitigation of abiotic and biotic stresses makes it a more versatile molecule. Moreover, it significantly reduces the percentage of losses during postharvest storage among different fruits and vegetables. Furthermore, the regulation of gene expression and crosstalk with other phytohormones is another important factor of melatonin, which contributes greatly to many plant biological processes under both normal and unfriendly environmental conditions. However, the endogenously produced melatonin sometimes is not enough to tackle harsh scenarios. For that reason, exogenous melatonin and some other approaches were implemented to induce the level of endogenous melatonin in order to sustain plant immunity and normal growth capacity. In addition, melatonin is considered as a nontoxic biodegradable molecule, which could be used for the promotion of organic farming [166]. To sum up, melatonin showed great importance across different plant science sectors; however, there is still no evidence available regarding the use of melatonin against viruses, nematodes, or insects; this requires further investigation.
Author Contributions
Conceptualization, R.S., C.X. and H.Z.; Methodology, M.B.A., Q.A. and M.A.; Formal Analysis, I.M.; Writing—Original Draft Preparation, R.S., A.S., I.M. and M.A.N.; Writing—Review & Editing, P.C.; Supervision, Y.L.; Project Administration, Y.L.
Funding
This research was funded by the National Key R&D Project, China (2016YFD0101705) and the National Natural Science Foundation of China (31471891and 31772300).
Acknowledgments
We apologize to the authors whose works are not cited because of space constraints.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lerner, A.B.; Case, J.D.; Takahashi, Y.; Lee, T.H.; Mori, W. Isolation of melatonin, the pineal gland factor that lightens melanocyte S1. J. Am. Chem. Soc. 1958, 80, 2587. [Google Scholar] [CrossRef]
- Reiter, R.J. Pineal melatonin: Cell biology of its synthesis and of its physiological interactions. Endocr. Rev. 1991, 12, 151–180. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Xu, B.; Zhou, X.; Reiter, R.J. Pineal Calcification, Melatonin Production, Aging, Associated Health Consequences and Rejuvenation of the Pineal Gland. Molecules 2018, 23, E301. [Google Scholar] [CrossRef] [PubMed]
- Posmyk, M.M.; Janas, K.M. Melatonin in plants. Acta Physiol. Plant. 2009, 31. [Google Scholar] [CrossRef]
- Nawaz, M.A.; Huang, Y.; Bie, Z.; Ahmed, W.; Reiter, R.J.; Niu, M.; Hameed, S. Melatonin: Current status and future perspectives in plant science. Front. Plant Sci. 2016, 6, 1230. [Google Scholar] [CrossRef] [PubMed]
- Lerner, A.; Case, J. Pigment cell regulatory factors. J. Investig. Dermatol. 1959, 32, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Sack, R.L.; Lewy, A.J.; Erb, D.L.; Vollmer, W.M.; Singer, C.M. Human melatonin production decreases with age. J. Pineal Res. 1986, 3, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Melatonin and the theories of aging: A critical appraisal of melatonin’s role in antiaging mechanisms. J. Pineal Res. 2013, 55, 325–356. [Google Scholar] [CrossRef] [PubMed]
- Tal, O.; Haim, A.; Harel, O.; Gerchman, Y. Melatonin as an antioxidant and its semi-lunar rhythm in green macroalga Ulva sp. J. Exp. Bot. 2011, 62, 1903–1910. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Growth conditions determine different melatonin levels in Lupinus albus L. J. Pineal Res. 2013, 55, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Byeon, Y.; Back, K. Melatonin synthesis in rice seedlings in vivo is enhanced at high temperatures and under dark conditions due to increased serotonin N-acetyltransferase and N-acetylserotonin methyltransferase activities. J. Pineal Res. 2014, 56, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.-X.; Zhou, Z.; Cruz, M.H.C.; Fuentes-Broto, L.; Galano, A. Phytomelatonin: Assisting plants to survive and thrive. Molecules 2015, 20, 7396–7437. [Google Scholar] [CrossRef] [PubMed]
- Kolář, J.; Macháčková, I. Melatonin in higher plants: Occurrence and possible functions. J. Pineal Res. 2005, 39, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Functions of melatonin in plants: A review. J. Pineal Res. 2015, 59, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Pape, C.; Lüning, K. Quantification of melatonin in phototrophic organisms. J. Pineal Res. 2006, 41, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Manchester, L.C.; Tan, D.-X.; Reiter, R.J.; Park, W.; Monis, K.; Qi, W. High levels of melatonin in the seeds of edible plants: Possible function in germ tissue protection. Life Sci. 2000, 67, 3023–3029. [Google Scholar] [CrossRef]
- Murch, S.J.; Saxena, P.K. Mammalian neurohormones: Potential significance in reproductive physiology of St. John’s wort (Hypericum perforatum L.)? Naturwissenschaften 2002, 89, 555–560. [Google Scholar] [PubMed]
- Kolár, J.; Johnson, C.H.; Machácková, I. Presence and possible role of melatonin in a short-day flowering plant, Chenopodium rubrum. In Melatonin after Four Decades: An Assessment of Its Potential; Olcese, J., Ed.; Springer: Berlin, Germany, 2002; pp. 391–393. [Google Scholar]
- Lee, H.Y.; Byeon, Y.; Back, K. Melatonin as a signal molecule triggering defense responses against pathogen attack in Arabidopsis and tobacco. J. Pineal Res. 2014, 57, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Byeon, Y.; Tan, D.X.; Reiter, R.J.; Back, K. Arabidopsis serotonin N-acetyltransferase knockout mutant plants exhibit decreased melatonin and salicylic acid levels resulting in susceptibility to an avirulent pathogen. J. Pineal Res. 2015, 58, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Jiang, C.; Ye, T.; Tan, D.-X.; Reiter, R.J.; Zhang, H.; Liu, R.; Chan, Z. Comparative physiological, metabolomic, and transcriptomic analyses reveal mechanisms of improved abiotic stress resistance in bermudagrass [Cynodon dactylon (L). Pers.] by exogenous melatonin. J. Exp. Bot. 2014, 66, 681–694. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Reiter, R.J.; Tan, D.X.; Chan, Z. INDOLE-3-ACETIC ACID INDUCIBLE 17 positively modulates natural leaf senescence through melatonin-mediated pathway in Arabidopsis. J. Pineal Res. 2015, 58, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Tan, D.X.; Reiter, R.J.; Ye, T.; Yang, F.; Chan, Z. Melatonin induces class A1 heat-shock factors (HSFA1s) and their possible involvement of thermotolerance in Arabidopsis. J. Pineal Res. 2015, 58, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Chen, Y.; Tan, D.X.; Reiter, R.J.; Chan, Z.; He, C. Melatonin induces nitric oxide and the potential mechanisms relate to innate immunity against bacterial pathogen infection in Arabidopsis. J. Pineal Res. 2015, 59, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Tan, D.-X.; Reiter, R.J.; Shi, H. Comparative metabolomic analysis highlights the involvement of sugars and glycerol in melatonin-mediated innate immunity against bacterial pathogen in Arabidopsis. Sci. Rep. 2015, 5, 15815. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Sun, Q.; Zhang, H.; Cao, Y.; Weeda, S.; Ren, S.; Guo, Y.-D. Roles of melatonin in abiotic stress resistance in plants. J. Exp. Bot. 2014, 66, 647–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, N.; Sun, Q.; Li, H.; Li, X.; Cao, Y.; Zhang, H.; Li, S.; Zhang, L.; Qi, Y.; Ren, S. Melatonin improved anthocyanin accumulation by regulating gene expressions and resulted in high reactive oxygen species scavenging capacity in cabbage. Front. Plant Sci. 2016, 7, 197. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Manchester, L.C.; Liu, X.; Rosales-Corral, S.A.; Acuna-Castroviejo, D.; Reiter, R.J. Mitochondria and chloroplasts as the original sites of melatonin synthesis: A hypothesis related to melatonin’s primary function and evolution in eukaryotes. J. Pineal Res. 2013, 54, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B. Phytomelatonin: Discovery, content, and role in plants. Adv. Bot. 2014, 2014. [Google Scholar] [CrossRef]
- Zhao, D.; Wang, R.; Liu, D.; Wu, Y.; Sun, J.; Tao, J. Melatonin and Expression of Tryptophan Decarboxylase Gene (TDC) in Herbaceous Peony (Paeonia lactiflora Pall.) Flowers. Molecules 2018, 23, 1164. [Google Scholar] [CrossRef] [PubMed]
- Vantasselv, D.; Li, J.; Oneill, S. Melatonin-identification of a potential dark signal in plants. Plant Physiol. 1993, 102, 117. [Google Scholar]
- Vantassel, D.; Roberts, N.; Oenill, S. Melatonin from higher-plants-isolation and identification of N-acetyl 5-methoxytryptamine. Plant Physiol. 1995, 108, 101. [Google Scholar]
- Burkhardt, S.; Tan, D.X.; Manchester, L.C.; Hardeland, R.; Reiter, R.J. Detection and quantification of the antioxidant melatonin in Montmorency and Balaton tart cherries (Prunus cerasus). J. Agric. Food. Chem. 2001, 49, 4898–4902. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; Alan, A.R.; Cao, J.; Saxena, P.K. Melatonin and serotonin in flowers and fruits of Datura metel L. J. Pineal Res. 2009, 47, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, M.; Ezura, H. Profiling of melatonin in the model tomato (Solanum lycopersicum L.) cultivar Micro-Tom. J. Pineal Res. 2009, 46, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Liu, G.; Chang, Y.; Lin, D.; Reiter, R.J.; He, C.; Shi, H. Melatonin biosynthesis enzymes recruit WRKY transcription factors to regulate melatonin accumulation and transcriptional activity on W-box in cassava. J. Pineal Res. 2018, 65, E12487. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Wang, M.; Zhao, Y.; Han, P.; Dai, Y. Melatonin from different fruit sources, functional roles, and analytical methods. Trends Food Sci. Technol. 2014, 37, 21–31. [Google Scholar] [CrossRef]
- Luboshitzky, R.; Yanai, D.; Shen-Orr, Z.; Israeli, E.; Herer, P.; Lavie, P. Daily and seasonal variations in the concentration of melatonin in the human pineal gland. Brain Res. Bull. 1998, 47, 271–276. [Google Scholar] [CrossRef]
- Underwood, H. The pineal and melatonin: Regulators of circadian function in lower vertebrates. Experientia 1989, 45, 914–922. [Google Scholar] [CrossRef]
- Badria, F.A. Melatonin, serotonin, and tryptamine in some Egyptian food and medicinal plants. J. Med. Food 2002, 5, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Hattori, A.; Migitaka, H.; Iigo, M.; Itoh, M.; Yamamoto, K.; Ohtani-Kaneko, R.; Hara, M.; Suzuki, T.; Reiter, R.J. Identification of melatonin in plants and its effects on plasma melatonin levels and binding to melatonin receptors in vertebrates. Biochem. Mol. Biol. Int. 1995, 35, 627–634. [Google Scholar] [PubMed]
- Hernández-Ruiz, J.; Cano, A.; Arnao, M.B. Melatonin acts as a growth-stimulating compound in some monocot species. J. Pineal Res. 2005, 39, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Chemical stress by different agents affects the melatonin content of barley roots. J. Pineal Res. 2009, 46, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Posmyk, M.M.; Bałabusta, M.; Wieczorek, M.; Sliwinska, E.; Janas, K. Melatonin applied to cucumber (Cucumis sativus L.) seeds improves germination during chilling stress. J. Pineal Res. 2009, 46, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Riga, P.; Medina, S.; García-Flores, L.A.; Gil-Izquierdo, Á. Melatonin content of pepper and tomato fruits: Effects of cultivar and solar radiation. Food Chem. 2014, 156, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, Y.; Rebollo-Hernanz, M.; Herrera, T.; Cayuelas, L.T.; Rodríguez-Rodríguez, P.; de Pablo, Á.L.L.; Arribas, S.M.; Martin-Cabrejas, M.A. Intake of bean sprouts influences melatonin and antioxidant capacity biomarker levels in rats. Food Funct. 2016, 7, 1438–1445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Back, K. Melatonin promotes seminal root elongation and root growth in transgenic rice after germination. J. Pineal Res. 2012, 53, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Le, T.N.N.; Byeon, Y.; Kim, Y.S.; Back, K. Transient induction of melatonin biosynthesis in rice (Oryza sativa L.) during the reproductive stage. J. Pineal Res. 2013, 55, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Huo, Y.; Tan, D.-X.; Liang, Z.; Zhang, W.; Zhang, Y. Melatonin in Chinese medicinal herbs. Life Sci. 2003, 73, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Arnao, M.B.; Hernández-Ruiz, J. Growth conditions influence the melatonin content of tomato plants. Food Chem. 2013, 138, 1212–1214. [Google Scholar] [CrossRef] [PubMed]
- Menendez-Pelaez, A.; Poeggeler, B.; Reiter, R.J.; Barlow-Walden, L.; Pablos, M.I.; Tan, D.X. Nuclear localization of melatonin in different mammalian tissues: Immunocytochemical and radioimmunoassay evidence. J. Cell. Biochem. 1993, 53, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; Campbell, S.S.; Saxena, P.K. The role of serotonin and melatonin in plant morphogenesis: Regulation of auxin-induced root organogenesis in in vitro-cultured explants of St. John’s wort (Hypericum perforatum L.). In Vitro Cell. Dev. Biol.-Plant 2001, 37, 786–793. [Google Scholar] [CrossRef]
- Kołodziejczyk, I.; Dzitko, K.; Szewczyk, R.; Posmyk, M.M. Exogenous melatonin expediently modifies proteome of maize (Zea mays L.) embryo during seed germination. Acta Physiol. Plant. 2016, 38, 146. [Google Scholar] [CrossRef]
- Wei, W.; Li, Q.-T.; Chu, Y.-N.; Reiter, R.J.; Yu, X.-M.; Zhu, D.-H.; Zhang, W.-K.; Ma, B.; Lin, Q.; Zhang, J.-S. Melatonin enhances plant growth and abiotic stress tolerance in soybean plants. J. Exp. Bot. 2014, 66, 695–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez-Ruiz, J.; Cano, A.; Arnao, M.B. Melatonin: A growth-stimulating compound present in lupin tissues. Planta 2004, 220, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin promotes adventitious-and lateral root regeneration in etiolated hypocotyls of Lupinus albus L. J. Pineal Res. 2007, 42, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Sun, Y.; Liu, Z.; Jin, W.; Sun, Y. Effects of melatonin on seedling growth, mineral nutrition, and nitrogen metabolism in cucumber under nitrate stress. J. Pineal Res. 2017, 62, E12403. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Li, A.; Yu, H.; Li, W.; Liang, C.; Guo, S.; Zhang, R.; Chu, C. Melatonin regulates root architecture by modulating auxin response in rice. Front. Plant Sci. 2017, 8, 134. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Z.; Sun, L.; Wang, T.; Miao, P.; Zhu, X.; Liu, S.; Song, F.; Mao, H.; Li, X. Melatonin Improves the Photosynthetic Carbon Assimilation and Antioxidant Capacity in Wheat Exposed to Nano-ZnO Stress. Molecules 2017, 22, 1727. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Tan, D.X.; Jiang, D.; Liu, F. Melatonin enhances cold tolerance in drought-primed wild-type and abscisic acid-deficient mutant barley. J. Pineal Res. 2016, 61, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Sarropoulou, V.N.; Therios, I.N.; Dimassi-Theriou, K.N. Melatonin promotes adventitious root regeneration in in vitro shoot tip explants of the commercial sweet cherry rootstocks CAB-6P (Prunus cerasus L.), Gisela 6 (P. cerasus× P. canescens), and MxM 60 (P. avium× P. mahaleb). J. Pineal Res. 2012, 52, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wei, J.-P.; Scott, E.R.; Liu, J.-W.; Guo, S.; Li, Y.; Zhang, L.; Han, W.-Y. Exogenous Melatonin Alleviates Cold Stress by Promoting Antioxidant Defense and Redox Homeostasis in Camellia sinensis L. Molecules 2018, 23, 165. [Google Scholar] [CrossRef] [PubMed]
- Denancé, N.; Sánchez-Vallet, A.; Goffner, D.; Molina, A. Disease resistance or growth: The role of plant hormones in balancing immune responses and fitness costs. Front. Plant Sci. 2013, 4, 155. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin and its relationship to plant hormones. Ann. Bot. 2017, 121, 195–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelagio-Flores, R.; Muñoz-Parra, E.; Ortiz-Castro, R.; López-Bucio, J. Melatonin regulates Arabidopsis root system architecture likely acting independently of auxin signaling. J. Pineal Res. 2012, 53, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; An, B.; Wei, Y.; Reiter, R.J.; Shi, H.; Luo, H.; He, C. Melatonin regulates root meristem by repressing auxin synthesis and polar auxin transport in Arabidopsis. Front. Plant Sci. 2016, 7, 1882. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Qi, W.B.; Reiter, R.J.; Wei, W.; Wang, B.M. Exogenously applied melatonin stimulates root growth and raises endogenous indoleacetic acid in roots of etiolated seedlings of Brassica juncea. J. Plant Physiol. 2009, 166, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Giridhar, A.R.P.; Ravishankar, G. Indoleamines and calcium channels influence morphogenesis in in vitro cultures of Mimosa pudica L. Plant Signal. Behav. 2009, 4, 1136–1141. [Google Scholar] [CrossRef]
- Sharif, R.; Mujtaba, M.; Ur Rahman, M.; Shalmani, A.; Ahmad, H.; Anwar, T.; Tianchan, D.; Wang, X. The Multifunctional Role of Chitosan in Horticultural Crops; A Review. Molecules 2018, 23, 872. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Shao, J.; Shi, L.; Xu, L.; Shen, Z.; Chen, W.; Yang, Z. Melatonin increases chilling tolerance in postharvest peach fruit by alleviating oxidative damage. Sci. Rep. 2018, 8, 806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazari, F.; Saba, M.K. Combination Effect of 1-Methylcyclopropene (1-MCP) with Ajowan Essential Oil and Silver Nanoparticles on Postharvest Life of Gerbera (Gerbera jamesonii) Cut Flowers. HortScience 2017, 52, 1550–1555. [Google Scholar] [CrossRef]
- Wu, B.; Guo, Q.; Wang, G.X.; Peng, X.Y.; Che, F.B. Effects of different postharvest treatments on the physiology and quality of ‘Xiaobai’apricots at room temperature. J. Food Sci. Technol. 2015, 52, 2247–2255. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Wang, Z.; Cao, J.; Cui, H.; Ma, Z. Cold stress increases reactive oxygen species formation via TRPA1 activation in A549 cells. Cell Stress Chaperones 2016, 21, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Tie, W.; Ou, W.; Yan, Y.; Kong, H.; Zuo, J.; Ding, X.; Ding, Z.; Liu, Y.; Wu, C. Crosstalk between calcium and melatonin affects postharvest physiological deterioration and quality loss in cassava. Postharvest Biol. Technol. 2018, 140, 42–49. [Google Scholar] [CrossRef]
- Arah, I.K.; Amaglo, H.; Kumah, E.K.; Ofori, H. Preharvest and postharvest factors affecting the quality and shelf life of harvested tomatoes: A mini review. Int. J. Agron. 2015, 2015. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, R.; Sun, Y.; Liu, Z.; Jin, W.; Sun, Y. The beneficial effects of exogenous melatonin on tomato fruit properties. Sci. Hortic. 2016, 207, 14–20. [Google Scholar] [CrossRef]
- Zhu, Z.; Lee, B. Friends or foes: New insights in jasmonate and ethylene co-actions. Plant Cell Physiol. 2015, 56, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wei, J.; Zhang, M.; Zhang, L.; Li, C.; Wang, Q. Ethylene independent induction of lycopene biosynthesis in tomato fruits by jasmonates. J. Exp. Bot. 2012, 63, 5751–5761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Pirrello, J.; Chervin, C.; Roustan, J.-P.; Bouzayen, M. Ethylene control of fruit ripening: Revisiting the complex network of transcriptional regulation. Plant Physiol. 2015, 169, 2380–2390. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, V.; Prabha, T.; Tharanathan, R. Fruit ripening phenomena–an overview. Crit. Rev. Food Sci. Nutr. 2007, 47, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Zhang, N.; Wang, J.; Zhang, H.; Li, D.; Shi, J.; Li, R.; Weeda, S.; Zhao, B.; Ren, S. Melatonin promotes ripening and improves quality of tomato fruit during postharvest life. J. Exp. Bot. 2014, 66, 657–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Zhu, M.; Ren, L.; Li, A.; Chen, G.; Hu, Z. The SlFSR Gene Controls Fruit Shelf-Life in Tomato. J. Exp. Bot. 2018, 2897–2909. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, X.; Chen, T.; Ji, Y.; Shi, K.; Wang, L.; Zheng, X.; Kong, J. Melatonin in Apples and Juice: Inhibition of Browning and Microorganism Growth in Apple Juice. Molecules 2018, 23, 521. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Yang, H.; Tie, W.; Yan, Y.; Ding, Z.; Liu, Y.; Wu, C.; Wang, J.; Reiter, R.J.; Tan, D.-X. Natural Variation in Banana Varieties Highlights the Role of Melatonin in Postharvest Ripening and Quality. J. Agric. Food Chem. 2017, 65, 9987–9994. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Hu, H.; Luo, S.; Wu, Z.; Li, P. Melatonin delaying senescence of postharvest broccoli by regulating respiratory metabolism and antioxidant activity. Trans. Chin. Soc. Agric. Eng. 2018, 34, 300–308. [Google Scholar]
- Ma, Q.; Zhang, T.; Zhang, P.; Wang, Z.Y. Melatonin attenuates postharvest physiological deterioration of cassava storage roots. J. Pineal Res. 2016, 60, 424–434. [Google Scholar] [CrossRef] [PubMed]
- Marta, B.; Szafrańska, K.; Posmyk, M.M. Exogenous melatonin improves antioxidant defense in cucumber seeds (Cucumis sativus L.) germinated under chilling stress. Front. Plant Sci. 2016, 7, 575. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Zhang, Z.K.; Chai, H.K.; Cheng, N.; Yang, Y.; Wang, D.N.; Yang, T.; Cao, W. Melatonin treatment delays postharvest senescence and regulates reactive oxygen species metabolism in peach fruit. Postharvest Biol. Technol. 2016, 118, 103–110. [Google Scholar] [CrossRef]
- Zhai, R.; Liu, J.; Liu, F.; Zhao, Y.; Liu, L.; Fang, C.; Wang, H.; Li, X.; Wang, Z.; Ma, F. Melatonin limited ethylene production, softening and reduced physiology disorder in pear (Pyrus communis L.) fruit during senescence. Postharvest Biol. Technol. 2018, 139, 38–46. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Fard, J.R. Melatonin treatment attenuates postharvest decay and maintains nutritional quality of strawberry fruits (Fragaria × anannasa cv. Selva) by enhancing GABA shunt activity. Food Chem. 2017, 221, 1650–1657. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zheng, H.; Sheng, K.; Liu, W.; Zheng, L. Effects of melatonin treatment on the postharvest quality of strawberry fruit. Postharvest Biol. Technol. 2018, 139, 47–55. [Google Scholar] [CrossRef]
- Xu, W.; Cai, S.Y.; Zhang, Y.; Wang, Y.; Ahammed, G.J.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Yu, J.Q.; Reiter, R.J. Melatonin enhances thermotolerance by promoting cellular protein protection in tomato plants. J. Pineal Res. 2016, 61, 457–469. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Gao, F.; Ni, Z.; Lin, L.; Deng, Q.; Tang, Y.; Wang, X.; Luo, X.; Xia, H. Melatonin improves heat tolerance in kiwifruit seedlings through promoting antioxidant enzymatic activity and glutathione S-transferase transcription. Molecules 2018, 23, 584. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.F.; Xu, T.F.; Wang, Z.Z.; Fang, Y.L.; Xi, Z.M.; Zhang, Z.W. The ameliorative effects of exogenous melatonin on grape cuttings under water-deficient stress: Antioxidant metabolites, leaf anatomy, and chloroplast morphology. J. Pineal Res. 2014, 57, 200–212. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, J.; Wang, W.; Sun, Y. Exogenous melatonin improves growth and photosynthetic capacity of cucumber under salinity-induced stress. Photosynthetica 2016, 54, 19–27. [Google Scholar] [CrossRef]
- Zhang, H.J.; Zhang, N.; Yang, R.C.; Wang, L.; Sun, Q.Q.; Li, D.B.; Cao, Y.Y.; Weeda, S.; Zhao, B.; Ren, S. Melatonin promotes seed germination under high salinity by regulating antioxidant systems, ABA and GA4 interaction in cucumber (Cucumis sativus L.). J. Pineal Res. 2014, 57, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shi, Y.; Zhang, X.; Du, H.; Xu, B.; Huang, B. Melatonin suppression of heat-induced leaf senescence involves changes in abscisic acid and cytokinin biosynthesis and signaling pathways in perennial ryegrass (Lolium perenne L.). Environ. Exp. Bot. 2017, 138, 36–45. [Google Scholar] [CrossRef]
- Fan, J.; Hu, Z.; Xie, Y.; Chan, Z.; Chen, K.; Amombo, E.; Chen, L.; Fu, J. Alleviation of cold damage to photosystem II and metabolisms by melatonin in Bermudagrass. Front. Plant Sci. 2015, 6, 925. [Google Scholar] [CrossRef] [PubMed]
- Tiryaki, I.; Keles, H. Reversal of the inhibitory effect of light and high temperature on germination of Phacelia tanacetifolia seeds by melatonin. J. Pineal Res. 2012, 52, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Hernández, I.G.; Gomez, F.J.V.; Cerutti, S.; Arana, M.V.; Silva, M.F. Melatonin in Arabidopsis thaliana acts as plant growth regulator at low concentrations and preserves seed viability at high concentrations. Plant Physiol. Biochem. 2015, 94, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Tan, D.-X.; Liang, D.; Chang, C.; Jia, D.; Ma, F. Melatonin mediates the regulation of ABA metabolism, free-radical scavenging, and stomatal behaviour in two Malus species under drought stress. J. Exp. Bot. 2014, 66, 669–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Zhou, J.; Tan, D.-X.; Wang, N.; Wang, L.; Shan, D.; Kong, J. Melatonin improves waterlogging tolerance of Malus baccata (Linn.) Borkh. seedlings by maintaining aerobic respiration, photosynthesis and ROS migration. Front. Plant Sci. 2017, 8, 483. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.; Hernández-Ruiz, J. Protective effect of melatonin against chlorophyll degradation during the senescence of barley leaves. J. Pineal Res. 2009, 46, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zeng, L.; Cheng, Y.; Lu, G.; Fu, G.; Ma, H.; Liu, Q.; Zhang, X.; Zou, X.; Li, C. Exogenous melatonin alleviates damage from drought stress in Brassica napus L.(rapeseed) seedlings. Acta Physiol. Plant. 2018, 40, 43. [Google Scholar] [CrossRef]
- Hu, Z.; Fan, J.; Xie, Y.; Amombo, E.; Liu, A.; Gitau, M.M.; Khaldun, A.; Chen, L.; Fu, J. Comparative photosynthetic and metabolic analyses reveal mechanism of improved cold stress tolerance in bermudagrass by exogenous melatonin. Plant Physiol. Biochem. 2016, 100, 94–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Li, Y.; Tian, Y.; Qu, M.; Zhang, W.; Gao, L. Melatonin Has the Potential to Alleviate Cinnamic Acid Stress in Cucumber Seedlings. Front. Plant Sci. 2017, 8, 1193. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; Li, J.; Li, H. Effects of Exogenous Melatonin on Photosynthetic Characteristics of Eggplant (Solanum melongena L.) under Cadmium Stress. In Advances in Engineering Research, Proceedings of the 2015 6th International Conference on Manufacturing Science and Engineering, Guangzhou, China, 28–29 December 2015; Atlantis Press: Hong Kong, China, 2015; Volume 32, pp. 353–356. [Google Scholar]
- Dawood, M.G.; El-Awadi, M.E. Alleviation of salinity stress on Vicia faba L. plants via seed priming with melatonin. Acta Biológica Colomb. 2015, 20, 223–235. [Google Scholar] [CrossRef]
- Ye, J.; Wang, S.; Deng, X.; Yin, L.; Xiong, B.; Wang, X. Melatonin increased maize (Zea mays L.) seedling drought tolerance by alleviating drought-induced photosynthetic inhibition and oxidative damage. Acta Physiol. Plant. 2016, 38, 48. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, S.; Yang, S.; Chen, Y. Melatonin alleviates cold-induced oxidative damage by regulation of ascorbate–glutathione and proline metabolism in melon seedlings (Cucumis melo L.). J. Hortic. Sci. Biotechnol. 2017, 92, 313–324. [Google Scholar] [CrossRef]
- Antoniou, C.; Chatzimichail, G.; Xenofontos, R.; Pavlou, J.J.; Panagiotou, E.; Christou, A.; Fotopoulos, V. Melatonin systemically ameliorates drought stress-induced damage in Medicago sativa plants by modulating nitro-oxidative homeostasis and proline metabolism. J. Pineal Res. 2017, 62, e12401. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, P.; Wei, Z.; Liang, D.; Liu, C.; Yin, L.; Jia, D.; Fu, M.; Ma, F. The mitigation effects of exogenous melatonin on salinity-induced stress in Malus hupehensis. J. Pineal Res. 2012, 53, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Shi, S.; Dou, F.; Song, Y.; Ma, F. Exogenous Melatonin Alleviates Alkaline Stress in Malus hupehensis Rehd. by Regulating the Biosynthesis of Polyamines. Molecules 2017, 22, 1542. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wang, A.; Li, X.; Kou, M.; Wang, W.; Chen, X.; Xu, T.; Zhu, M.; Ma, D.; Li, Z. Melatonin-Stimulated Triacylglycerol Breakdown and Energy Turnover under Salinity Stress Contributes to the Maintenance of Plasma Membrane H+–ATPase Activity and K+/Na+ Homeostasis in Sweet Potato. Front. Plant Sci. 2018, 9, 256. [Google Scholar] [CrossRef] [PubMed]
- Szafrańska, K.; Reiter, R.J.; Posmyk, M.M. Melatonin application to Pisum sativum L. seeds positively influences the function of the photosynthetic apparatus in growing seedlings during paraquat-induced oxidative stress. Front. Plant Sci. 2016, 7, 1663. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Zheng, G.; Li, W.; Wang, Y.; Hu, B.; Wang, H.; Wu, H.; Qian, Y.; Zhu, X.G.; Tan, D.X. Melatonin delays leaf senescence and enhances salt stress tolerance in rice. J. Pineal Res. 2015, 59, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Posmyk, M.M.; Kuran, H.; Marciniak, K.; Janas, K.M. Presowing seed treatment with melatonin protects red cabbage seedlings against toxic copper ion concentrations. J. Pineal Res. 2008, 45, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zeng, B.; Mao, Y.; Kong, X.; Wang, X.; Yang, Y.; Zhang, J.; Xu, J.; Rengel, Z.; Chen, Q. Melatonin alleviates aluminium toxicity through modulating antioxidative enzymes and enhancing organic acid anion exudation in soybean. Funct. Plant Biol. 2017, 44, 961–968. [Google Scholar] [CrossRef]
- Mukherjee, S.; David, A.; Yadav, S.; Baluška, F.; Bhatla, S.C. Salt stress-induced seedling growth inhibition coincides with differential distribution of serotonin and melatonin in sunflower seedling roots and cotyledons. Physiol. Plant. 2014, 152, 714–728. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.L.; Xu, H.; Li, D.; Gao, X.; Li, T.; Wang, R. Effect of melatonin priming on photosynthetic capacity of tomato leaves under low-temperature stress. Photosynthetica 2018, 56, 884–892. [Google Scholar] [CrossRef]
- Zhou, X.; Zhao, H.; Cao, K.; Hu, L.; Du, T.; Baluška, F.; Zou, Z. Beneficial roles of melatonin on redox regulation of photosynthetic electron transport and synthesis of D1 protein in tomato seedlings under salt stress. Front. Plant Sci. 2016, 7, 1823. [Google Scholar] [CrossRef] [PubMed]
- Martinez, V.; Nieves-Cordones, M.; Lopez-Delacalle, M.; Rodenas, R.; Mestre, T.C.; Garcia-Sanchez, F.; Rubio, F.; Nortes, P.A.; Mittler, R.; Rivero, R.M. Tolerance to Stress Combination in Tomato Plants: New Insights in the Protective Role of Melatonin. Molecules 2018, 23, 535. [Google Scholar] [CrossRef] [PubMed]
- Debnath, B.; Hussain, M.; Irshad, M.; Mitra, S.; Li, M.; Liu, S.; Qiu, D. Exogenous melatonin mitigates acid rain stress to tomato plants through modulation of leaf ultrastructure, photosynthesis and antioxidant potential. Molecules 2018, 23, 388. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chang, J.; Chen, H.; Wang, Z.; Gu, X.; Wei, C.; Zhang, Y.; Ma, J.; Yang, J.; Zhang, X. Exogenous melatonin confers salt stress tolerance to watermelon by improving photosynthesis and redox homeostasis. Front. Plant Sci. 2017, 8, 295. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.A.; Jiao, Y.; Chen, C.; Shireen, F.; Zheng, Z.; Imtiaz, M.; Bie, Z.; Huang, Y. Melatonin pretreatment improves vanadium stress tolerance of watermelon seedlings by reducing vanadium concentration in the leaves and regulating melatonin biosynthesis and antioxidant-related gene expression. J. Plant Physiol. 2018, 220, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chang, J.; Zheng, J.; Dong, Y.; Liu, Q.; Yang, X.; Wei, C.; Zhang, Y.; Ma, J.; Zhang, X. Local melatonin application induces cold tolerance in distant organs of Citrullus lanatus L. via long distance transport. Sci. Rep. 2017, 7, 40858. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.; Zhao, X.; Liu, S.; Sun, F.; Zhang, C.; Xi, Y. Beneficial effects of melatonin in overcoming drought stress in wheat seedlings. Plant Physiol. Biochem. 2017, 118, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Wang, Q.; Shah, F.A.; Liu, W.; Wang, D.; Huang, S.; Fu, S.; Wu, L. Exogenous Melatonin Confers Cadmium Tolerance by Counterbalancing the Hydrogen Peroxide Homeostasis in Wheat Seedlings. Molecules 2018, 23, 799. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Wang, P.; Li, M.; Ke, X.; Li, C.; Liang, D.; Wu, S.; Ma, X.; Li, C.; Zou, Y. Exogenous melatonin improves Malus resistance to Marssonina apple blotch. J. Pineal Res. 2013, 54, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.-W.; Ma, W. Phytohormone pathways as targets of pathogens to facilitate infection. Plant Mol. Biol. 2016, 91, 713–725. [Google Scholar] [CrossRef] [PubMed]
- Marco, S.; Levy, D. Involvement of ethylene in the development of cucumber mosaic virus-induced chlorotic lesions in cucumber cotyledonst. Physiol. Plant Pathol. 1979, 14, 235–244. [Google Scholar] [CrossRef]
- Alazem, M.; Lin, N.S. Roles of plant hormones in the regulation of host–virus interactions. Mol. Plant Pathol. 2015, 16, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Jameson, P.E.; Clarke, S.F. Hormone-virus interactions in plants. Crit. Rev. Plant Sci. 2002, 21, 205–228. [Google Scholar] [CrossRef]
- Dehkordi, A.N.; Rubio, M.; Babaeian, N.; Albacete, A.; Martínez-Gómez, P. Phytohormone Signaling of the Resistance to Plum pox virus (PPV, Sharka Disease) Induced by Almond (Prunus dulcis (Miller) Webb) Grafting to Peach (P. persica L. Batsch). Viruses 2018, 10, 238. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Hu, W.; Wang, Q.; Zeng, H.; Li, X.; Yan, Y.; Reiter, R.J.; He, C.; Shi, H. Identification, transcriptional and functional analysis of heat-shock protein 90s in banana (Musa acuminata L.) highlight their novel role in melatonin-mediated plant response to Fusarium wilt. J. Pineal Res. 2017, 62, e12367. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zheng, X.; Reiter, R.J.; Feng, S.; Liu, S.; Wang, Y.; Jin, L.; Li, Z.; Datla, R.; Ren, M. Melatonin Attenuates Potato late Blight by Disrupting cell Growth, Stress Tolerance, Fungicide Susceptibility and Homeostasis of gene Expression in Phytophthora infestans. Front. Plant Sci. 2017, 8, 1993. [Google Scholar] [CrossRef] [PubMed]
- Jannin, L.; Arkoun, M.; Ourry, A.; Laîné, P.; Goux, D.; Garnica, M.; Fuentes, M.; San Francisco, S.; Baigorri, R.; Cruz, F. Microarray analysis of humic acid effects on Brassica napus growth: Involvement of N, C and S metabolisms. Plant Soil 2012, 359, 297–319. [Google Scholar] [CrossRef]
- Varela-Valencia, R.; Gómez-Ortiz, N.; Oskam, G.; de Coss, R.; Rubio-Piña, J.; del Río-García, M.; Albores-Medina, A.; Zapata-Perez, O. The effect of titanium dioxide nanoparticles on antioxidant gene expression in tilapia (Oreochromis niloticus). J. Nanoparticle Res. 2014, 16, 2369. [Google Scholar] [CrossRef]
- Zhang, N.; Zhang, H.J.; Zhao, B.; Sun, Q.Q.; Cao, Y.Y.; Li, R.; Wu, X.X.; Weeda, S.; Li, L.; Ren, S.; et al. The RNA-seq approach to discriminate gene expression profiles in response to melatonin on cucumber lateral root formation. J. Pineal Res. 2014, 56, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Sheshadri, S.; Nishanth, M.; Yamine, V.; Simon, B. Effect of Melatonin on the stability and expression of reference genes in Catharanthus roseus. Sci. Rep. 2018, 8, 2222. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Liu, Z.; Zhu, L.; Ma, Z.; Wang, J.; Zhu, J. Exogenous melatonin improves plant iron deficiency tolerance via increased accumulation of polyamine-mediated nitric oxide. Int. J. Mol. Sci. 2016, 17, 1777. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, S.; Chen, Y. Effects of melatonin on photosynthetic performance and antioxidants in melon during cold and recovery. Biol. Plant. 2017, 61, 571–578. [Google Scholar] [CrossRef]
- Berg, J.A.; Appiano, M.; Bijsterbosch, G.; Visser, R.G.; Schouten, H.J.; Bai, Y. Functional characterization of cucumber (Cucumis sativus L.) Clade V MLO genes. BMC Plant Biol. 2017, 17, 80. [Google Scholar] [CrossRef] [PubMed]
- Schouten, H.J.; Krauskopf, J.; Visser, R.G.; Bai, Y. Identification of candidate genes required for susceptibility to powdery or downy mildew in cucumber. Euphytica 2014, 200, 475–486. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Sun, X.; Wang, N.; Tan, D.X.; Ma, F. Melatonin enhances the occurrence of autophagy induced by oxidative stress in Arabidopsis seedlings. J. Pineal Res. 2015, 58, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Gao, T.; Liang, B.; Zhao, Q.; Ma, F.; Li, C. Effects of Exogenous Melatonin on Methyl Viologen-Mediated Oxidative Stress in Apple Leaf. Int. J. Mol. Sci. 2018, 19, 316. [Google Scholar] [Green Version]
- Keunen, E.; Peshev, D.; Vangronsveld, J.; Van Den Ende, W.; Cuypers, A. Plant sugars are crucial players in the oxidative challenge during abiotic stress: Extending the traditional concept. Plant Cell Environ. 2013, 36, 1242–1255. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Xu, L.; Su, T.; Jiang, Y.; Hu, L.; Ma, F. Melatonin regulates carbohydrate metabolism and defenses against Pseudomonas syringae pv. tomato DC3000 infection in Arabidopsis thaliana. J. Pineal Res. 2015, 59, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Robles, P.; Quesada, V. Emerging Roles of Mitochondrial Ribosomal Proteins in Plant Development. Int. J. Mol. Sci. 2017, 18, 2595. [Google Scholar] [CrossRef] [PubMed]
- Jagadis Gupta, K.; Kumari, A.; Florez-Sarasa, I.; Fernie, A.R.; Igamberdiev, A.U. Interaction of nitric oxide with the components of plant mitochondrial electron transport chain. J. Exp. Bot. 2018, 69, 3413–3424. [Google Scholar] [CrossRef] [PubMed]
- Igamberdiev, A.U.; Hill, R.D. Elevation of cytosolic Ca2+ in response to energy deficiency in plants: The general mechanism of adaptation to low oxygen stress. Biochem. J. 2018, 475, 1411–1425. [Google Scholar] [CrossRef] [PubMed]
- Mur, L.A.; Mandon, J.; Persijn, S.; Cristescu, S.M.; Moshkov, I.E.; Novikova, G.V.; Hall, M.A.; Harren, F.J.; Hebelstrup, K.H.; Gupta, K.J. Nitric oxide in plants: An assessment of the current state of knowledge. AoB Plants 2013, 5, pls052. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Zhu, Y.; Cheng, H.; Yan, H.; Zhao, L.; Tang, J.; Ma, X.; Mao, P. Nitric Oxide Regulates Seedling Growth and Mitochondrial Responses in Aged Oat Seeds. Int. J. Mol. Sci. 2018, 19, 1052. [Google Scholar] [CrossRef] [PubMed]
- Franco, D.G.; Moretti, I.F.; Marie, S.K.N. Mitochondria Transcription Factor A: A Putative Target for the Effect of Melatonin on U87MG Malignant Glioma Cell Line. Molecules 2018, 23, 1129. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Kang, K.; Lee, K.; Back, K. Characterization of tryptamine 5-hydroxylase and serotonin synthesis in rice plants. Plant Cell Rep. 2007, 26, 2009–2015. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Kang, K.; Lee, K.; Back, K. Characterization of rice tryptophan decarboxylases and their direct involvement in serotonin biosynthesis in transgenic rice. Planta 2007, 227, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, M.; Higuchi, K.; Hanawa, Y.; Shiraiwa, Y.; Ezura, H. Cloning and characterization of a Chlamydomonas reinhardtii cDNA arylalkylamine N-acetyltransferase and its use in the genetic engineering of melatonin content in the Micro-Tom tomato. J. Pineal Res. 2009, 46, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, T.; Maisonneuve, S.; Isshiki, M.; Mizutani, M.; Chen, L.; Wong, H.L.; Kawasaki, T.; Shimamoto, K. Sekiguchi lesion gene encodes a cytochrome P450 monooxygenase that catalyzes conversion of tryptamine to serotonin in rice. J. Biol. Chem. 2010, 285, 11308–11313. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.; Kong, K.; Park, S.; Natsagdorj, U.; Kim, Y.S.; Back, K. Molecular cloning of a plant N-acetylserotonin methyltransferase and its expression characteristics in rice. J. Pineal Res. 2011, 50, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.; Lee, K.; Park, S.; Byeon, Y.; Back, K. Molecular cloning of rice serotonin N-acetyltransferase, the penultimate gene in plant melatonin biosynthesis. J. Pineal Res. 2013, 55, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Zuo, B.; Zheng, X.; He, P.; Wang, L.; Lei, Q.; Feng, C.; Zhou, J.; Li, Q.; Han, Z.; Kong, J. Overexpression of MzASMT improves melatonin production and enhances drought tolerance in transgenic Arabidopsis thaliana plants. J. Pineal Res. 2014, 57, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Choi, G.H.; Lee, H.Y.; Back, K. Chloroplast overexpression of rice caffeic acid O-methyltransferase increases melatonin production in chloroplasts via the 5-methoxytryptamine pathway in transgenic rice plants. J. Pineal Res. 2017, 63, e12412. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.Y.; Zhang, Y.; Xu, Y.P.; Qi, Z.Y.; Li, M.Q.; Ahammed, G.J.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Reiter, R.J.; et al. HsfA1a upregulates melatonin biosynthesis to confer cadmium tolerance in tomato plants. J. Pineal Res. 2017, 62, e12387. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Jiao, J.; Fan, X.; Sun, H.; Zhang, Y.; Jiang, J.; Liu, C. Endophytic bacterium Pseudomonas fluorescens RG11 may transform tryptophan to melatonin and promote endogenous melatonin levels in the roots of four grape cultivars. Front. Plant Sci. 2017, 7, 2068. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; Ma, Y.; Chen, S.; Liu, C.; Song, Y.; Qin, Y.; Yuan, C.; Liu, Y. Melatonin-producing endophytic bacteria from grapevine roots promote the abiotic stress-induced production of endogenous melatonin in their hosts. Front. Plant Sci. 2016, 7, 1387. [Google Scholar] [CrossRef] [PubMed]
- Janas, K.M.; Posmyk, M.M. Melatonin, an underestimated natural substance with great potential for agricultural application. Acta Physiol. Plant. 2013, 35, 3285–3292. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Currently there is no sample availaible. |
Figure 1.
Schematic representation of melatonin defense mechanism pathway.


{kind=link}
Table 1.
Reported concentration of melatonin in some edible plants.
| Crop | Used Methodology | Melatonin Content (pg/g FW(DW) Tissue) | Reference |
|---|---|---|---|
| Apple | GC-MS | 0.16 | [40] |
| Asparagus | RIA | 9.5 | [41] |
| Barley | LC | 500–12,000 R; 82,300 S | [42,43] |
| Cucumber fruit seeds | RIA | 24.6, 11,000 | [41,44] |
| Chilies | UHPLC-MS/MS | 31–93 | [45] |
| Kiwi | RIA | 0.02 | [41] |
| Kidney bean | ELISA | 529 DW | [46] |
| Rice | HPLC | 100 L; 500 S; 200 R; 400 Fl | [47,48] |
| Sunflower | HPLC | 29,000 DW | [16] |
| Tea (Shiya green tea) | HPLC | 2.12 μg g−1 | [49] |
| Tomato | LC | 15,000–142,000 L | [50] |
| Wheat | LC | 124,700 S | [42] |
Abbreviation: L = leaf, R = roots, FL = flower, GC-MS = gas chromatographic-mass spectroscopy, RIA = radioimmunoassay, LC = liquid chromatography, UHPLC-MS/MS = Ultra-high performance liquid chromatography coupled to mass spectrometry in tandem mode, ELISA = Enzyme-linked immunosorbent assay, HPLC = High performance liquid chromatography.
Table 2.
Effect of melatonin on postharvest produce.
| Crop | Stress/Condition | Concentration | Functional Improvement | Reference |
|---|---|---|---|---|
| Apple | Browning | 250 mg/L | Prevented apple juice from browning | [83] |
| Banana | Quality improvement | 50–500 μM | Slowed down ripening, low ethylene production, accelerate endogenous melatonin. | [84] |
| Broccoli | senescence | 100 μM/L | Maintained postharvest freshness | [85] |
| Cabbage | Cold | 100 μM/L | Enhanced anthocyanin activity and antioxidant capacities | [27] |
| Cassava | Hydrogen peroxide | 500 mg/L | Delayed postharvest physiological and root deterioration in cassava | [74,86] |
| Cucumber | Cold | 500 μM | Increased protection against cold-induced oxidative stress in seeds | [87] |
| Peach | Oxidative | 0.1 mmol/L | Slow down the senescence, increased antioxidant enzymatic activities and ascorbic acid content | [70,88] |
| Pear | Quality improvement | 100 μM | Slowed senescence process, increase antioxidants, less fruit firmness losses, exhibited to be a strong scavenger of ROS | [89] |
| Strawberry | Fungal, quality improvement | 1000 μmol/L or 100 μmol/L | triggered H2O2 accumulation, higher SOD activity, delayed senescence, decay, weight losses, maintained fruit firmness, titratable acidity, increased total phenol, flavonoids and antioxidant activity | [90,91] |
| Tomato | Quality improvement trial | 50 μM | Promotes ripening, upregulated the expression level of fruit color development genes and altered the ethylene production. | [81] |
Table 3.
Protective role of melatonin in various crops against different abiotic stresses.
| Crop | Stress Condition | Concentration | Functions | Reference |
|---|---|---|---|---|
| Arabidopsis | Heat | 1000 μM | Improved seed germination under heat stress | [100] |
| Apple | Drought | 100 µM | Reduced ABA activity and radical scavenging | [101] |
| Apple | Waterlogging | 200 μM | Reduced chlorosis and wilting of the seedlings | [102] |
| Barley | Senescence | 1 mM | Boosted chlorophyll content | [103] |
| Brassica napus L. | Drought | 0.05 mmol/L | Increased the overall growth indices of brassica seedlings | [104] |
| Bermuda grass | Cold | 100 μM | Induced photosynthetic activity under cold stress | [105] |
| Cucumber | Salinity | 100 μM | Overall growth | [95] |
| Cucumber | Cinnamic acid | 10 μM | Rescued cucumber seedlings from Cinnamic acid stress and increased the allocation of dry weight in roots. | [106] |
| Eggplant | Cadmium stress | 150 μmol/L | Enriched photosynthetic activity | [107] |
| Faba bean | Salinity | 500 μM | Enriched photosynthetic activity and mineral accumulation | [108] |
| Grapes | Water deficient | 200 μmol/L | Amended antioxidative enzymes activity | [94] |
| Maize | Drought | 100 μmol/L | Photosynthesis and growth | [109] |
| Melon | Cold | 200 μM | Improved proline and ascorbic acid content | [110] |
| Medicago sativa | Drought | 10 μM | regulation of nitro-oxidative and osmoprotective homeostasis | [111] |
| Malus hupehensis | Salinity | 0.1 mM | Improved photosynthetic activity and better plant growth | [112] |
| Malus hupehensis | Alkaline | 5 µM | Significantly induced the tolerance against alkaline stress by increasing the antioxidant activity and biosynthesis of polyamines | [113] |
| Perennial ryegrass | High temperature | 20 μM | Regulate abscisic acid and cytokinin biosynthesis | [97] |
| Potato | Salinity | 100 µM | Better chlorophyll content, antioxidant activities and water content | [114] |
| Pisum sativum L. | Oxidative stress | 50 μM | Reduced O2•− accumulation in leaf tissues and preservation of photosynthetic pigments | [115] |
| Rice | Salinity | 20 μM | Delay leaf senescence and cell death in rice | [116] |
| Red cabbage | Heavy metal | 10 μM | Improved seed germination and reduced the toxic effect of metal on the seedling. | [117] |
| Soybean | Multiple stress | 100 µM | Boost and maintain the overall plant growth | [54] |
| Soybean | Aluminum stress | 50 μM | Enhanced root growth and reduced aluminum toxicity | [118] |
| Sunflower | Salt | 15 μM | Regulate root growth and hypocotyl elongation under salt stress | [119] |
| Tomato | Cold and salinity | 100 μM | Improved photosynthesis and regulation of photosynthetic electron transport | [120,121] |
| Tomato | Heat and salinity | 100 μM | Induced antioxidant enzymes activity and better photosynthetic performance | [122] |
| Tomato | Acid rain | 100 μM | Enhanced tolerance against simulated acid rain and increased the photosynthetic activity | [123] |
| Tea | Cold | 100 μM | Triggered photosynthetic and antioxidant enzymes activities | [62] |
| Watermelon | Salinity | 150 μM | Redox homeostasis and improved photosynthetic activity | [124] |
| Watermelon | Vanadium stress | 0.1 μM | Lower the concentration of vanadium in leaf, stem and better photosynthetic and antioxidants activity | [125] |
| Watermelon | Cold | 150 μM and 1.5 μM | Alleviate cold stress by inducing long-distance signaling in the untreated tissue. | [126] |
| Wheat | Drought and nano-ZnO | 500 μM and 1 mM | Augmented seedling percentage, growth, and antioxidant enzymes activities. | [59,127] |
| Wheat | Cadmium stress | 50 mM | Reduce the level of hydrogen peroxide which increases the wheat plants growth | [128] |
Table 4.
Defense mechanism induced by melatonin against biotic stresses in different plants.
| Crop | Pathogen | Concentration | Beneficial Functions | Reference |
|---|---|---|---|---|
| Apple | Diplocarpon mali | 0.1 mM | Improved resistance to apple blotch disease | [129] |
| Arabidopsis | Pseudomonas syringae | 10 μM | Increased the resistance by suppressing the bacterium about 10-fold | [19] |
| Arabidopsis | Pseudomonas syringae | 10 μM | Alternatively, increased the resistance by triggering the level of endogenous salicylic acid. | [20] |
| Banana | Fusarium oxysporum | 100 μM | Induce resistance in banana against the pathogen attack | [135] |
| Lupinus albus | Penicillium spp. | 70 μM | Enhanced resistance against the fungal pathogen | [14] |
| Potato | Phytophthora infestans | 5 mM | Inhibited the potato late blight disease by arresting the mycelial growth | [136] |
| Strawberry | Botrytis cinerea and Rhizopus stolonifer | 1000 μmol/L | Attenuating fungal decay and maintaining nutritional quality of strawberry fruits | [90] |
| Tobacco | Pseudomonas syringae | 10 μM | Increased the resistance by suppressing the bacterium about 10-fold | [19] |
Table 5.
Role of exogenous melatonin in regulating gene expression.
| Crop | Stress/Conditions | Genes | Expression | Functions | Reference |
|---|---|---|---|---|---|
| Arabidopsis thaliana | Iron deficiency | FIT1, FRO2, IRT1 | ↑ | Increased plants tolerance to Fe deficiency | [141] |
| Arabidopsis thaliana | Oxidative | AtAPX1, AtCATs | ↑ | Removed damaged protein via the activation of autophagy | [145] |
| Arabidopsis thaliana | Heat | HSFA2, HSA32 | ↑ | Activated thermotolerance related genes in quadruple knockout mutant | [23] |
| Apple | Oxidative stress | MdTDC1, MdT5H4, MdAANAT2, and MdASMT1 | ↓ | Slowing the decline in chlorophyll concentrations, restraining membrane damage and lipid peroxidation | [146] |
| Cabbage | Oxidative | MYB, bHLH, WD40 | ↑ | Enhanced anthocyanins accumulation and increased antioxidant activities. | [27] |
| Melon | Cold | CmSOD, CmPOD, and CmCAT | ↑ | Recovered melon from cold stress through regulation of antioxidant activities | [142] |
| Potato | Salinity | SDP1, LACS6, LACS7, and ACX4 | ↑ | Maintenance of PM HC–ATPase activity and KC/NaC homeostasis and increase tolerance to salinity stress | [114] |
| Peach | Cold | PpAPX1, PpAPX3, PpAPX4, and PpAPX7 | ↑ | Activated the expression of genes involved in ASA-GSH cycle which improved resistance to cold stress | [70] |
| Peony | Fluctuating light | TDC | ↑↓ | Controls the production of melatonin biosynthesis under changing light spectrum | [30] |
| Rice | OsARF, OsSAUR | ↑ | Regulate rice root architecture on auxin dependent signalling manner | [58] | |
| Tomato | Salinity and Heat | SlcAPX, SlGR1, SlGST, and SlPh-GPX | ↑ | Enhanced tolerance to multiple stress by activating antioxidant enzymes system | [122] |
| Watermelon | Vanadium | Cla018095, Cla009820, Cla012125 | ↑ | Improved chlorophyll content and antioxidant activities | [125] |
| Wheat | Drought | APX and MDHAR4 | ↑ | Increase tolerance against drought stress | [127] |
↑ showing up-regulation and ↓ down-regulation of the respective genes presented in the table.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sharif, R.; Xie, C.; Zhang, H.; Arnao, M.B.; Ali, M.; Ali, Q.; Muhammad, I.; Shalmani, A.; Nawaz, M.A.; Chen, P.; et al. Melatonin and Its Effects on Plant Systems. Molecules 2018, 23, 2352. https://doi.org/10.3390/molecules23092352
AMA Style
Sharif R, Xie C, Zhang H, Arnao MB, Ali M, Ali Q, Muhammad I, Shalmani A, Nawaz MA, Chen P, et al. Melatonin and Its Effects on Plant Systems. Molecules. 2018; 23(9):2352. https://doi.org/10.3390/molecules23092352
Chicago/Turabian StyleSharif, Rahat, Chen Xie, Haiqiang Zhang, Marino B. Arnao, Muhammad Ali, Qasid Ali, Izhar Muhammad, Abdullah Shalmani, Muhammad Azher Nawaz, Peng Chen, and et al. 2018. "Melatonin and Its Effects on Plant Systems" Molecules 23, no. 9: 2352. https://doi.org/10.3390/molecules23092352