Screening Vitis Genotypes for Responses to Botrytis cinerea and Evaluation of Antioxidant Enzymes, Reactive Oxygen Species and Jasmonic Acid in Resistant and Susceptible Hosts

 and
and
Abstract
:1. Introduction
2. Results
2.1. Grape Genotypes Exhibit Different Levels of Resistance to B. cinerea
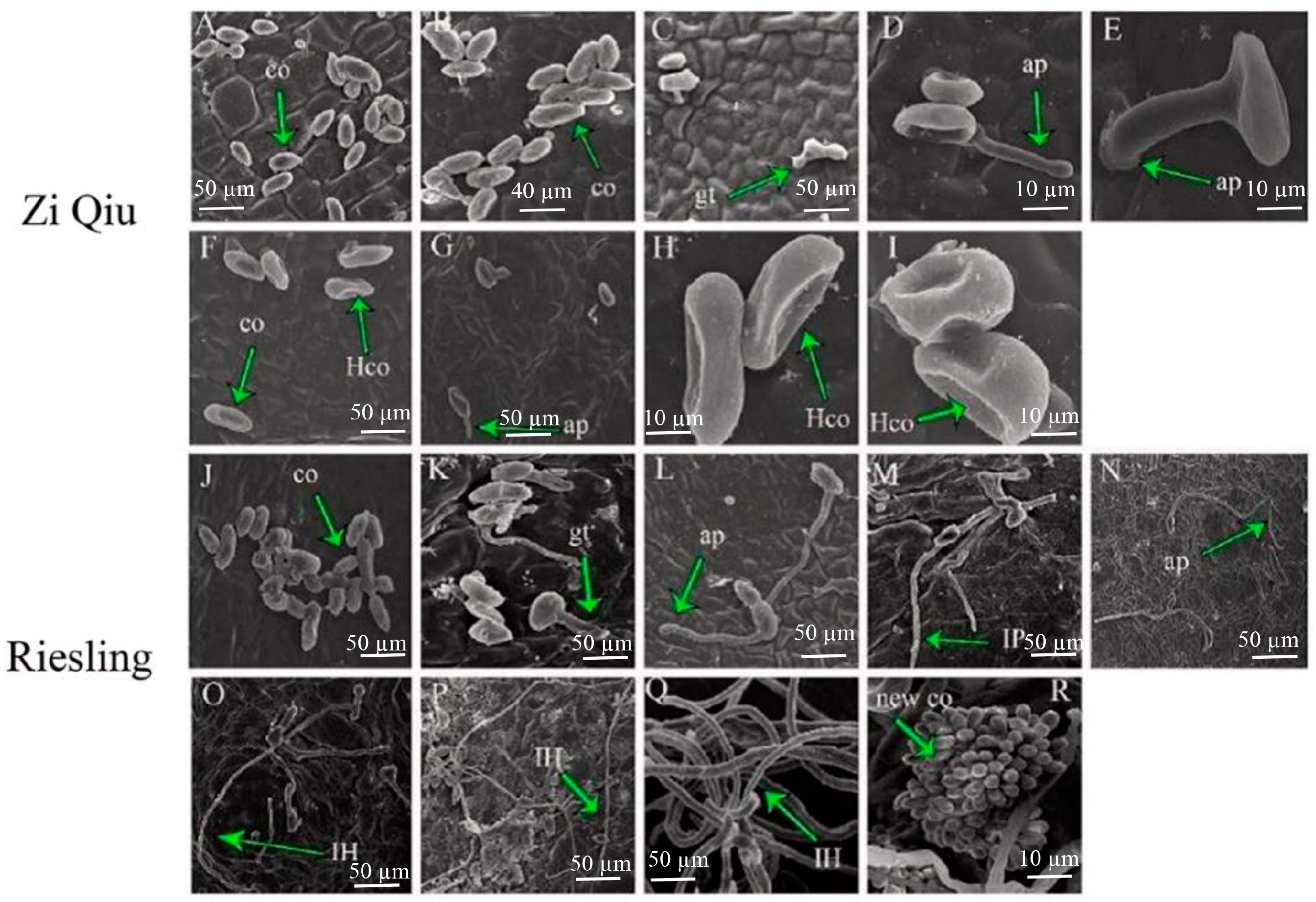
2.2. B. cinerea Growth on the HR Genotype ’Zi Qiu’ and the HS Genotype ‘Riesling’
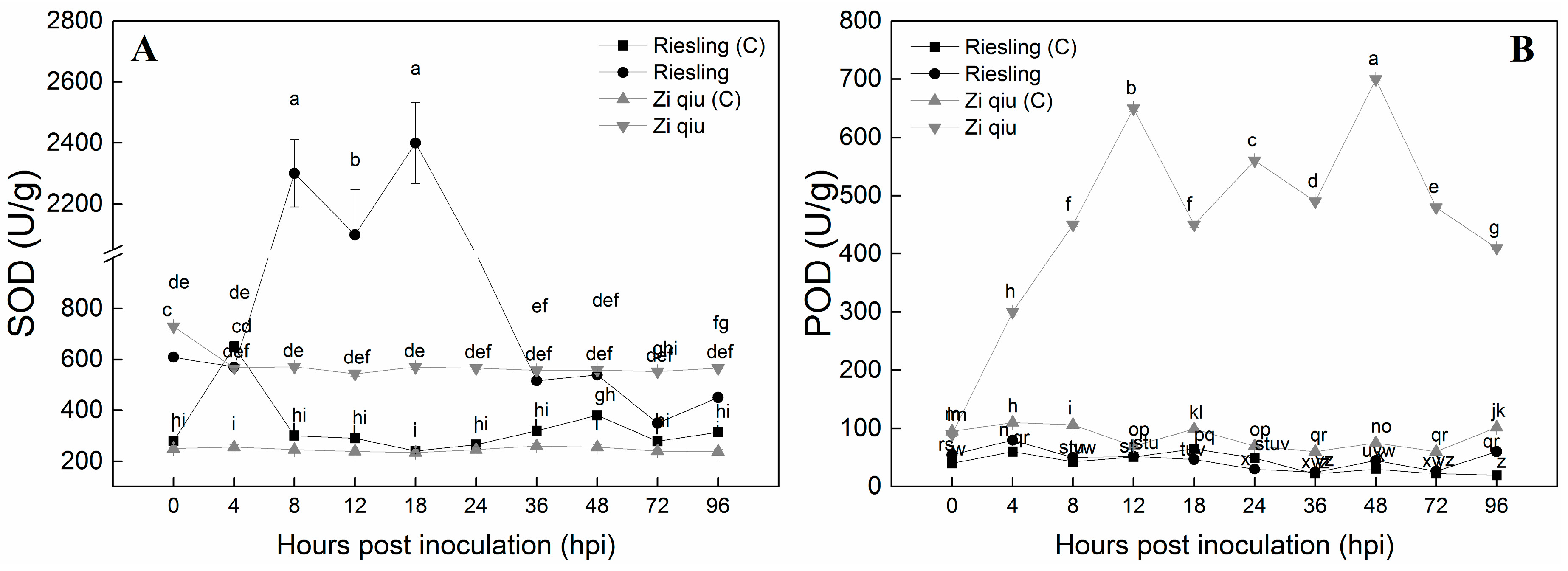
2.3. Activity of Peroxidase and Superoxide Dismutase in the HR Genotype ’Zi Qiu’ and the HS Genotype ‘Riesling’ Infected by B. cinerea
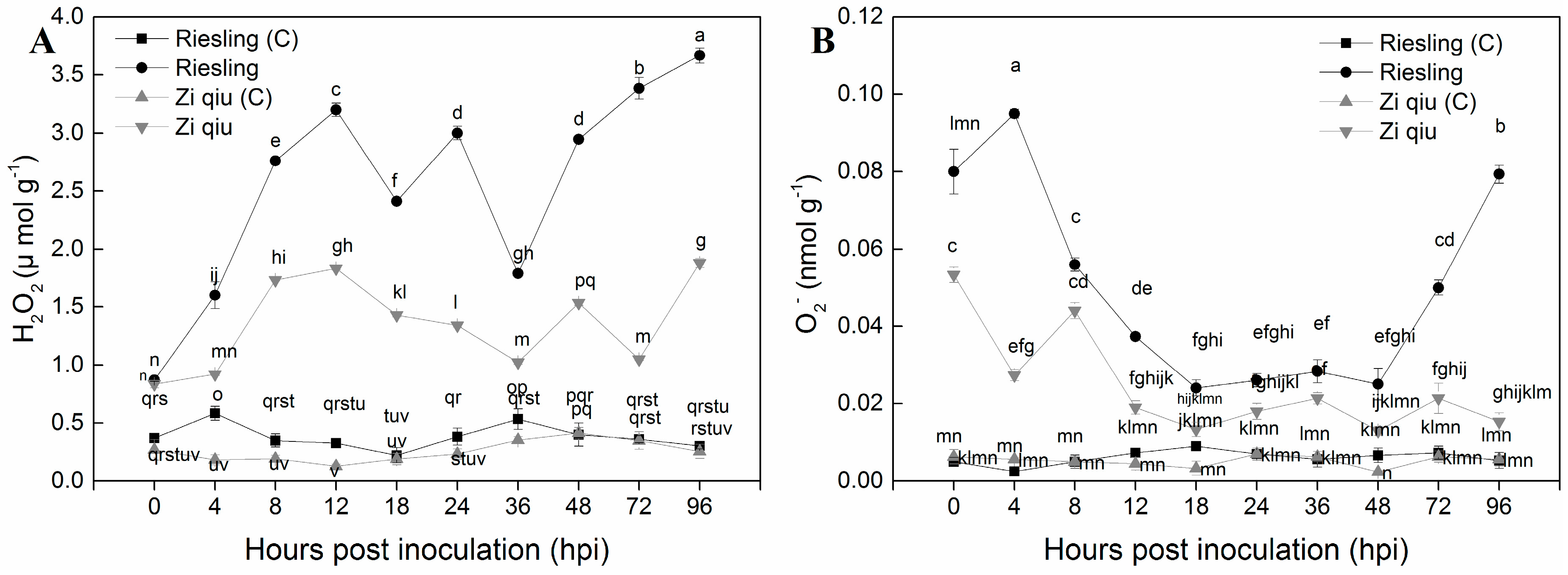
2.4. Hydrogen Peroxide (H2O2) Accumulation in HR ’Zi Qiu’ and HS ‘Riesling’ Leaves in Response to Infection with B. cinerea
2.5. Accumulation of Superoxide Radicals (O2−) in HR ’Zi Qiu’ and HS ‘Riesling’ Leaves in Response to Infection with B. cinerea
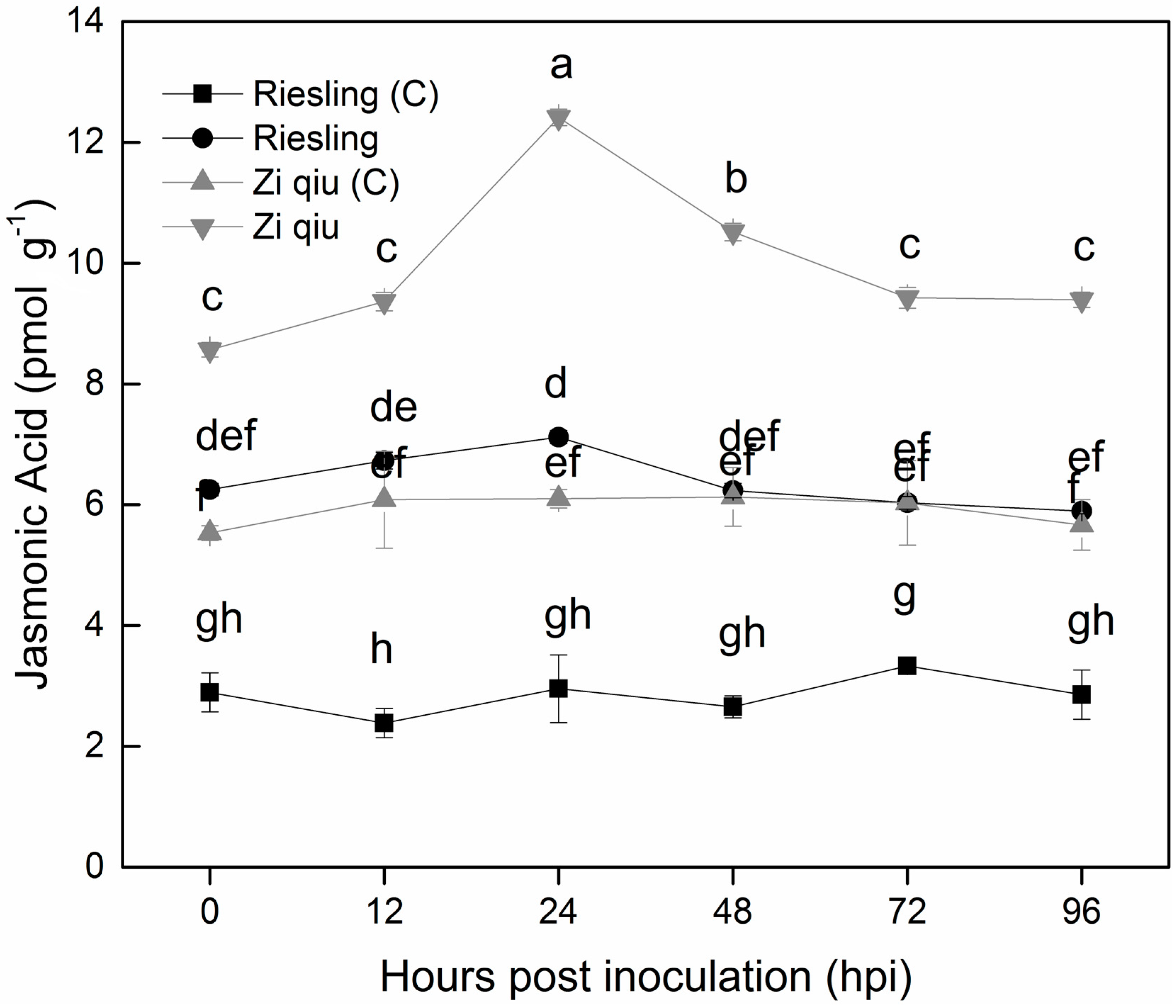
2.6. Jasmonic Acid Levels in Leaves of the HR Genotype ‘Zi Qiu’ and the HS Genotype ‘Riesling’ Following Inoculation with B. cinerea
2.7. Pearson’s Correlation Coefficients
3. Discussion
4. Materials and Methods
4.1. Plant and Fungal Materials
4.2. Detached Leaf Evaluation
4.3. Disease Severity Rating
4.4. Light Microscopy
4.5. SEM
4.6. ROS Measurement
4.6.1. H2O2 Measurement
4.6.2. O2− Measurement
4.7. Enzyme Extraction and Activity Assays
4.8. JA Quantification in HR and HS Genotypes
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ROS | reactive oxygen species |
| HR | highly resistant |
| HS | highly susceptible |
| SOD | superoxide dismutase |
| POD | peroxidase |
| JA | jasmonic acid |
| H2O2 | hydrogen peroxide |
| O2- | superoxide radicals |
| SI | severity index |
| HPI | hours post inoculation |
| SEM | scanning electron microscope |
References
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Cadle-davidson, L. Monitoring pathogenesis of natural. Vitis 2008, 4, 387–395. [Google Scholar]
- Percival, D.; Fisher, K.; Sullivan, J. Use of fruit zone leaf removal with Vitis vinifera L. cv. Riesling grapevines. II. Effect on fruit composition, yield and occurrence of bunch rot (Botrytis cinerea Pers.: Fr.). Am J. Enol. Vitic. 1994, 45, 133–140. [Google Scholar]
- Kulakiotu, E.K.; Thanassoulopoulos, C.C.; Sfakiotakis, E.M. Postharvest biological control of Botrytis cinerea on kiwifruit by volatiles of [Isabella] Grapes. Phytopathology 2004, 94, 1280–1285. [Google Scholar] [CrossRef] [PubMed]
- Gessler, C.; Pertot, I.; Perazzolli, M. Plasmoparaviticola: A review of knowledge on downy mildew of grapevine and effective disease management. Phytopathol. Mediterr. 2011, 50, 3–44. [Google Scholar]
- Angelini, D.M.R.M.; Masiello, M.; Rotolo, C.; Pollastro, S.; Faretra, F. Molecular characterisation and detection of resistance to succinate dehydrogenase inhibitor fungicides in botryotinia fuckeliana (Botrytis cinerea). Pest Manag. Sci. 2014, 70, 1884–1893. [Google Scholar] [CrossRef]
- Savocchia, S.; Stummer, B.E.; Wicks, T.J.; Vanheewijck, R.; Scorr, E.S. Reduced sensitivity of Uncinula necator to sterol demethylation inhibiting fungicides in southern Australian vineyards. Australas. Plant Pathol. 2004, 33, 465–473. [Google Scholar] [CrossRef]
- Kretschmer, M.; Leroch, M.; Mosbach, A.; Walker, A.S.; Fillinger, S.; Mernke, D.; Schoonbeek, H.; Pradier, J.M.; Leroux, P.W.; Hahn, M. Fungicide driven evolution and molecular basis of multidrug resistance in field population of grey mould fungus Botrytis cinerea. PLoS Pathog. 2009, 5, e1000696. [Google Scholar] [CrossRef]
- LI, H. Studies on the resistance of grapevine to powdery mildew. Plant Pathol. 1993, 42, 792–796. [Google Scholar] [CrossRef]
- Peros, J.P.; Nguyen, T.H.; Troulet, C.; Michel-Romitti, C.; Notteghem, J.L. Assessment of powdery mildew resistance of grape and Erysiphe necator pathogenicity using laboratory assay. Vitis 2006, 45, 29–36. [Google Scholar]
- Wan, R.; Hou, X.; Wang, X.; Qu, J.; Singer, S.D.; Wang, Y.; Wang, X. Resistance evaluation of Chinese wild Vitis genotypes against Botrytis cinerea and different responses of resistant and susceptible hosts to the infection. Front. Plant Sci. 2015, 6, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; He, C. Regulation of plant reactive oxygen species (ROS) in stress responses: Learning from AtRBOHD. Plant Cell Rep. 2016, 35, 995–1007. [Google Scholar] [CrossRef]
- Govrin, E.M.; Levine, A. The hypersensitive response facilitates plant infection by the necrotrophic pathogen Botrytis cinerea. Curr. Biol. 2000, 10, 751–757. [Google Scholar] [CrossRef]
- Temme, N.; Tudzynski, P. Does Botrytis cinerea ignore H2O2 -induced oxidative stress during infection? Characterization of botrytis activator protein 1. APS J. 2009, 22, 987–998. [Google Scholar]
- Durrant, W.E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef] [PubMed]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef]
- Poolsawat, O.; Tharapreuksapong, A.; Wongkaew, S.; Chaowiset, W.; Tantasawat, P. Laboratory and field evaluations of resistance to Sphaceloma ampelinum causing anthracnose in grapevine. Australas. Plant Pathol. 2012, 41, 263–269. [Google Scholar] [CrossRef]
- Karuppanapandian, T.; Moon, J.C.; Kim, C.; Manoharan, K.; Kim, W. Reactive oxygen species in plants: Their generation, signal transduction, and scavenging mechanisms. Aust. J. Crop Sci. 2011, 5, 709–725. [Google Scholar]
- Gabler, F.M.; Smilanick, J.L.; Mansour, M.; Ramming, D.W.; Mackey, B.E. Correlations of morphological, anatomical, and chemical features of grape berries with resistance to Botrytis cinerea. Phytopathology 2003, 93, 1263–1273. [Google Scholar] [CrossRef]
- Liu, L.; Li, H. Review: Research progress in amur grape, Vitis amurensis Rupr. Can. J. Plant Sci. 2013, 93, 565–575. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; He, P.; Chen, J.; Lamikanra, O.; Lu, J. Evaluation of foliar resistance to Uncinula necator in Chinese wild Vitis spp. species. Vitis 1995, 34, 159–164. [Google Scholar]
- Wang, Y.; Liu, Y.; He, P.; Lamikanra, O.; Lu, J. Resistance of Chinese Vitis species to Elsinoe ampelina (de Bary) Shear. HortScience 1998, 33, 123–126. [Google Scholar]
- Liu, R.; Wang, L.; Zhu, J.; Chen, T.; Wang, Y.; Xu, Y. Histological responses to downy mildew in resistant and susceptible grapevines. Protoplasma 2014, 252, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.A.; Jones, J.D.G.; Dangl, J.L. Reactive oxygen species signaling in response to pathogens. Plant Physiol. 2006, 141, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Redox signaling in plants. Antioxid. Redox Signal. 2013, 18, 2087–2090. [Google Scholar] [CrossRef] [PubMed]
- Asai, S.; Yoshioka, H. Nitric oxide as a partner of reactive oxygen species participates in disease resistance to necrotrophic pathogen Botrytis cinerea in nicotiana benthamiana. APS J. 2009, 22, 619–629. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.; Tognetti, V.B.; Vandepoele, K. ROS signaling: The new wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef]
- Smilanick, J.L.; Mansour, M.F.; Gabler, F.M.; Margosan, D.A.; Hashim-Buckey, J. Control of post-harvest gray mold of table grapes in the San Joaquin Valley of California by fungicides applied during the growing season. Plant Dis. 2010, 94, 250–257. [Google Scholar] [CrossRef]
- Asselbergh, B.; Curvers, K.; Franca, S.C.; Audenaert, K.; Vuylsteke, M.; Van Breusegem, F. Resistance to Botrytis cinerea in sitiens, an abscisic acid-deficient tomato mutant, involves timely production of hydrogen peroxide and cell wall modifications in the epidermis. Plant Physiol. 2007, 144, 1863–1877. [Google Scholar] [CrossRef] [PubMed]
- Patykowski, J. Role of hydrogen peroxide and apoplastic peroxidase in tomato-Botrytis cinerea interaction. Acta Physiol. Plant. 2006, 28, 589–598. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, B.; Li, X.; Ouyang, Z.; Huang, L.; Hong, Y. The denovo biosynthesis of vitamin B6 is required for disease resistance against Botrytis cinerea in tomato. Mol. Plant-Microbe Interact. 2014, 27, 688–699. [Google Scholar]
- Simon, U.K.; Polanschütz, L.M.; Koffler, B.E.; Zechmann, B. High resolution imaging of temporal and spatial changes of subcellular ascorbate, glutathione and H2O2 distribution during Botrytis cinerea infection in Arabidopsis. PLoS ONE 2013, 8, e65811. [Google Scholar] [CrossRef]
- Atkinson, N.J.; Urwin, P.E. In Posidonia oceanica cadmium induces changes in DNA methylation and chromatin patterning. J. Exp. Bot. 2012, 63, 695–709. [Google Scholar]
- Jia, H.; Zhang, C.; Pervaiz, T.; Zhao, P.; Liu, Z.; Wang, B. Jasmonic acid involves in grape fruit ripening and resistant against Botrytis cinerea. Funct. Integr. Genom. 2016, 16, 79–94. [Google Scholar] [CrossRef]
- Bruinsma, M.; Van Dam, N.M.; Van Loon, J.J.A.; Dicke, M. Jasmonic acid-induced changes in brassica oleracea affect oviposition preference of two specialist herbivores. J. Chem. Ecol. 2007, 33, 655–668. [Google Scholar] [CrossRef] [PubMed]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef]
- Li, C.; Schilmiller, A.L.; Liu, G.; Lee, G.I.; Jayanty, S.; Sageman, C. Role of beta-oxidation in jasmonate biosynthesis and systemic wound signaling in tomato. Plant Cell. 2005, 17, 971–986. [Google Scholar] [CrossRef]
- Kang, J.H.; Wang, L.; Giri, A.; Baldwin, I.T. Silencing threonine deaminase and JAR4 in nicotiana attenuata impairs jasmonic acid-isoleucine-mediated defenses against manduca sexta. Plant Cell. 2006, 18, 3303–3320. [Google Scholar] [CrossRef]
- Shan, X.; Zhang, Y.; Peng, W.; Wang, Z.; Xie, D. Molecular mechanism for jasmonate-induction of anthocyanin accumulation in arabidopsis. J. Exp. Bot. 2009, 60, 3849–3860. [Google Scholar] [CrossRef] [PubMed]
- Robert-Seilaniantz, A.; Navarro, L.; Bari, R.; Jones, J.D. Pathological hormone imbalances. Curr. Opin. Plant Biol. 2007, 10, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Liu, N.; Zhang, J.; Wang, Y. Field natural identification of resistance of grape resources from USA and Xinjiang of China to primary fungi diseases. J. Frui. Sci. 2011, 28, 849–856. [Google Scholar]
- Liu, C.; Ma, X.; Wu, G. Grape Varieties in China; Chinese Agriculture Press: Beijing, China, 2014. [Google Scholar]
- Li, D.; Wan, Y.; Wang, Y.; He, P. Relatedness of resistance to anthracnose and to white rot in Chinese wild grapes. Vit. J. Grapevine Res. 2008, 47, 213–215. [Google Scholar]
- Liu, S.M.; Sykes, S.R.; Clingeleffer, P.R. A method using leafed single-node cuttings to evaluate downy mildew resistance in grapevine. Vitis 2003, 42, 173–180. [Google Scholar]
- Cheng, Y.; Zhang, H.; Yao, J.; Wang, X.; Xu, J.; Han, Q. Characterization of non-host resistance in broad bean to the wheat stripe rust pathogen. BMC Plant Biol. 2012, 12, 96. [Google Scholar] [CrossRef]
- Moloi, M.J.; Westhuizen, A.J. The reactive oxygen species are involved in resistance responses of wheat to the Russian wheat aphid. J. Plant Physiol. 2006, 163, 1118–1125. [Google Scholar] [CrossRef]
- Elstner, E.F.; Heupel, A. Formation of hydrogen peroxide by isolated cell walls from horseradish (Armoracia lapathifolia Gilib). Planta 1976, 320, 317–320. [Google Scholar] [CrossRef]
- Maehly, A.C.; Chance, B. Theassayofcatalases andperoxidases. Meth. Biochem. Anal. 1954, 1, 357–424. [Google Scholar]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases. Biol. Roles Copp. 1977, 125–142. [Google Scholar]
- Royo, J.; Leon, J.; Vancanneyt, G.; Albar, J.P.; Rosahl, S.; Ortego, F. Antisense-mediated depletion of a potato lipoxygenase reduces wound induction of proteinase inhibitors and increases weight gain of insect pests. Proc. Natl. Acad. Sci. USA 1999, 96, 1146–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albrecht, T.; Kehlen, A.; Stahl, K.; Knöfel, H.D.; Sembdner, G.; Weiler, E.W. Quantification of rapid, transient increases in jasmonic acid in wounded plants using a monoclonal antibody. Planta 1993, 191, 86–94. [Google Scholar] [CrossRef]
Sample Availability: Not Available. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Name of Genotype | Disease Severity a (%) | Resistance Level b | Geographical Origin |
|---|---|---|---|---|
| V. vinifera L. | Beauty Seedless | 83.6 | HS | University of California, Davis, CA, USA |
| V. vinifera L. × V. amurensis Rupr | Beibinghong | 84.6 | HS | 1995, Institute of Special Animal and Plant Science of CAAS, Beijing, China |
| V. vinifera L. × V. amurensis Rupr | Beichun | 93.6 | HS | 1954, Beijing Botanical Garden Institute of Botany, Chinese Academy of Science, Beijing, China |
| V. vinifera L. × V. amurensis Rupr | Beihong | 42.4 | S | 1954, Beijing Botanical Garden Institute of Botany, Chinese Academy of Science, China |
| V. vinifera L. | Bixiang Seedless | 92.2 | HS | 1994, Jilin Academy of Agricultural Science, Changchun, China |
| V. vinifera L. | Black Rose | 76.5 | S | The Division of Horticultural Research of Commonwealth Scientific and Industrial Research Organization (CSIRO), Melbourne, Australia |
| V. vinifera L. | Blue French | 76.5 | S | Austria |
| V. vinifera L. | Blush Seedless | 71.8 | S | Professor Olmo, Davis Station, CA, USA. |
| V. vinifera L. | Cabernet Sauvignon | 22.7 | R | Aquitaine, Bordeaux, France |
| V. vinifera L. | Canner Seedless | 40.5 | S | University of California, Davis, CA, USA. |
| V. vinifera L. | Cannero | 80.5 | HS | Cannero Riviera, Italy |
| V. vinifera L. | Carignan | 93.7 | HS | Carinena, Aragon, Spain |
| V. vinifera L. | Chardonnay | 96.1 | HS | Burgundy, France |
| V. vinifera L. | Chenin Blanc | 27.4 | R | Anjou, Loire Valley, France |
| V. vinifera L. | Cinsault | 91.2 | HS | Languedoc, France |
| V. vinifera L. | Crimson Seedless | 94.2 | HS | Institute of Fruit Tree Research of Changli, Hebei Academy of Agriculture and Forestry Science, Shijiazhuang, China |
| V. vinifera L. | Dabai Grape | 73.2 | S | Unknown |
| V. vinifera L. × V. labrusca L. | Dong fang zhixing | 94.3 | HS | 2007, Hiroshima, Japan |
| V. vinifera L. | Early Muscat | 98.6 | HS | 1997, Shandong Province, Jinan, China |
| V. vinifera L. | Flame Seedless | 45.8 | S | 1973, Freson, CA, USA |
| V. vinifera L. | Fresno Seedless | 51.7 | S | Fresno, CA, USA |
| V. vinifera L. × V. labrusca L. | Gold Finger | 83.6 | HS | 1982, Japan |
| V. vinifera L. × V. labrusca L. | Hei bala duo | 39.2 | R | 2004, Kumamoto Prefecture, Japan |
| V. vinifera L. | Heise tiancai | 93.0 | HS | 2009, Kofu, Japan |
| V. vinifera L. | Hongmu nage | 58.7 | S | Atux, Xinjiang, China |
| V. vinifera L. × V. labrusca L. | Hutai NO.8 | 93.6 | HS | Grape Institute, Xian, China |
| V. vinifera L. | Italian Riesling | 59.9 | S | Italy |
| V. vinifera L. | Jing xiu | 92.0 | HS | 1994, Chinese Academy of Science, Beijing, China |
| V. vinifera L. | Jing ke jing | 81.2 | HS | 1984, Chinese Academy of Science, Beijing, China |
| V. vinifera L. | Jingzaojing | 91.8 | HS | 1984, Chinese Academy of Science, Beijing, China |
| V. vinifera L. | Jinxiangyu | 78.5 | HS | 1997, Chinese Academy of Science, Beijing, China |
| V. vinifera L. | Ju mei gui | 4.6 | HR | Dalian Academy of Agriculture Science, Dalian, China |
| V. riparia L. × V. labrusca L. | Kang san | 18.0 | R | Unknown |
| V. vinifera L.× V. labrusca L. | Kyoho | 25.7 | R | 1937, Japan |
| V. vinifera L. | Lady Finger | 87.6 | HS | 1984, Japan |
| V. vinifera L. | Long Emprer | 48.8 | S | Unknown |
| V. vinifera L. | Malta Seedless | 73.6 | S | Malta |
| V. vinifera L. | Manaizi | 95.2 | HS | Tulufan, Xinjiang, China |
| V. vinifera L. | Marselan | 24.6 | R | France |
| V. vinifera L. | May Purple | 38.0 | R | Unknown |
| V. vinifera L. | Merlot | 94.5 | HS | Bordeaux, France |
| V. vinifera L. | Moldova | 95.3 | HS | Moldova |
| V. vinifera L. | Mr Mori | 94.5 | HS | 1985, Japan |
| V. vinifera L. | Muscat Blanc | 63.4 | S | Eastern Mediterranean |
| V. vinifera L | Muscat of Alexandia | 42.0 | S | Egypt |
| V. vinifera L. | Muscat Rose | 78.1 | HS | Greece |
| V. vinifera L.× V. labrusca L. | Neptune Seedless | 47.2 | S | 1998, University of Arkansas, AR, USA. |
| V. vinifera L. | Otilia Seedless | 93.2 | HS | Romania |
| V. vinifera L. | Pearl of Csaba | 85.3 | HS | 1904, Hungary. |
| V. vinifera L. | Perlette | 59.5 | S | Fresno, CA, USA. |
| V. vinifera L. | Phoenix NO.51 | 88.5 | HS | Germany |
| V. vinifera L. | Pinot Noir | 94.0 | HS | Burgundy, France |
| V. vinifera L. | Prince Seedless | 23.4 | R | Hebei academy of agriculture and forestry science, Changli, China |
| V. vinifera L. | Qiu manai | 41.6 | S | Atushi, Xinjiang, China |
| V. vinifera L. | Queen | 24.6 | R | Unknown |
| V. vinifera L. | Red Globe | 85.0 | HS | University of California, CA, USA |
| V. vinifera L. | Red Hanepoot | 82.1 | HS | Unknown |
| V. vinifera L. | Riesling | 98.0 | HS | Germany |
| V. vinifera L. | Rizamat | 27.3 | R | The Soviet Union |
| V. vinifera L. | Rkatsiteli | 75.0 | S | Georgia |
| V. vinifera L. | Ruby Seedless | 86.3 | HS | University of California, CA, USA. |
| V. vinifera L. | Russian Seedless | 27.2 | R | Australia |
| V. vinifera L. | Sangiovese | 78.1 | HS | Italy |
| V. vinifera L. | Sauvignon Blanc | 71.4 | S | France |
| V. vinifera L. | Semillon | 93.2 | HS | France |
| V. vinifera L | Sultanina Rose | 54.6 | S | Unknown |
| V. vinifera L. × V. labrusca L. | Summer Black | 97.6 | HS | 1968, Yamanashi Prefecture, Japan |
| V. vinifera L. | Thompson Seedless | 76.2 | S | Tulufan, Xinjiang, China |
| V. vinifera L. × V. labrusca L. | Tian yuan qi | 90.0 | HS | 1994, Fruit Insitute, Liaoning, China |
| V. vinifera L. | Tokay | 93.9 | HS | Hungary |
| V. vinifera L. | Ugni Blanc | 97.6 | HS | Italy |
| V. vinifera L. | Victoria | 92.0 | HS | Romania |
| V. vinifera L. | Wink | 93.4 | HS | 1998, Japan |
| V. amurensis Rupr | Xue lanhong | 94.4 | HS | Institute of Special Animal and Plant Science of CAAS, Jilin, China |
| V. vinifera L. | Yangle | 55.8 | S | Russia |
| V. vinifera L. | Yatomi Rosa | 95.2 | HS | 1990, Japan |
| V. vinifera L. | Yili xiang | 69.8 | S | Yili, Xinjiang, China |
| V. vinifera L. × V. labrusca L. | Zao jin xiang | 28.2 | R | 1963, Grape institute, Liaoning, China |
| V. vinifera L. | Zhana | 63.8 | S | Albania |
| V. davidii Foex | Zi qiu | 3.3 | HR | 2004, Hunan Agricultural University, Hunan, China |
| V. vinifera L. × V. labrusca L. | Zui jin xiang | 67.8 | S | 1997, Liaoning Academy of Agriculture Science, China |
| JA | O2− | H2O2 | |
|---|---|---|---|
| SOD | −0.9959 * | 0.8182 * | 0.8974 * |
| POD | 0.0831 | 0.5278 * | 0.3975 * |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, M.U.; Hanif, M.; Wan, R.; Hou, X.; Ahmad, B.; Wang, X. Screening Vitis Genotypes for Responses to Botrytis cinerea and Evaluation of Antioxidant Enzymes, Reactive Oxygen Species and Jasmonic Acid in Resistant and Susceptible Hosts. Molecules 2019, 24, 5. https://doi.org/10.3390/molecules24010005
Rahman MU, Hanif M, Wan R, Hou X, Ahmad B, Wang X. Screening Vitis Genotypes for Responses to Botrytis cinerea and Evaluation of Antioxidant Enzymes, Reactive Oxygen Species and Jasmonic Acid in Resistant and Susceptible Hosts. Molecules. 2019; 24(1):5. https://doi.org/10.3390/molecules24010005
Chicago/Turabian StyleRahman, Mati Ur, Muhammad Hanif, Ran Wan, Xiaoqing Hou, Bilal Ahmad, and Xiping Wang. 2019. "Screening Vitis Genotypes for Responses to Botrytis cinerea and Evaluation of Antioxidant Enzymes, Reactive Oxygen Species and Jasmonic Acid in Resistant and Susceptible Hosts" Molecules 24, no. 1: 5. https://doi.org/10.3390/molecules24010005