Optimization of Light Intensity and NaNO3 Concentration in Amazon Cyanobacteria Cultivation to Produce Biodiesel


Abstract
:1. Introduction
2. Results and Discussion
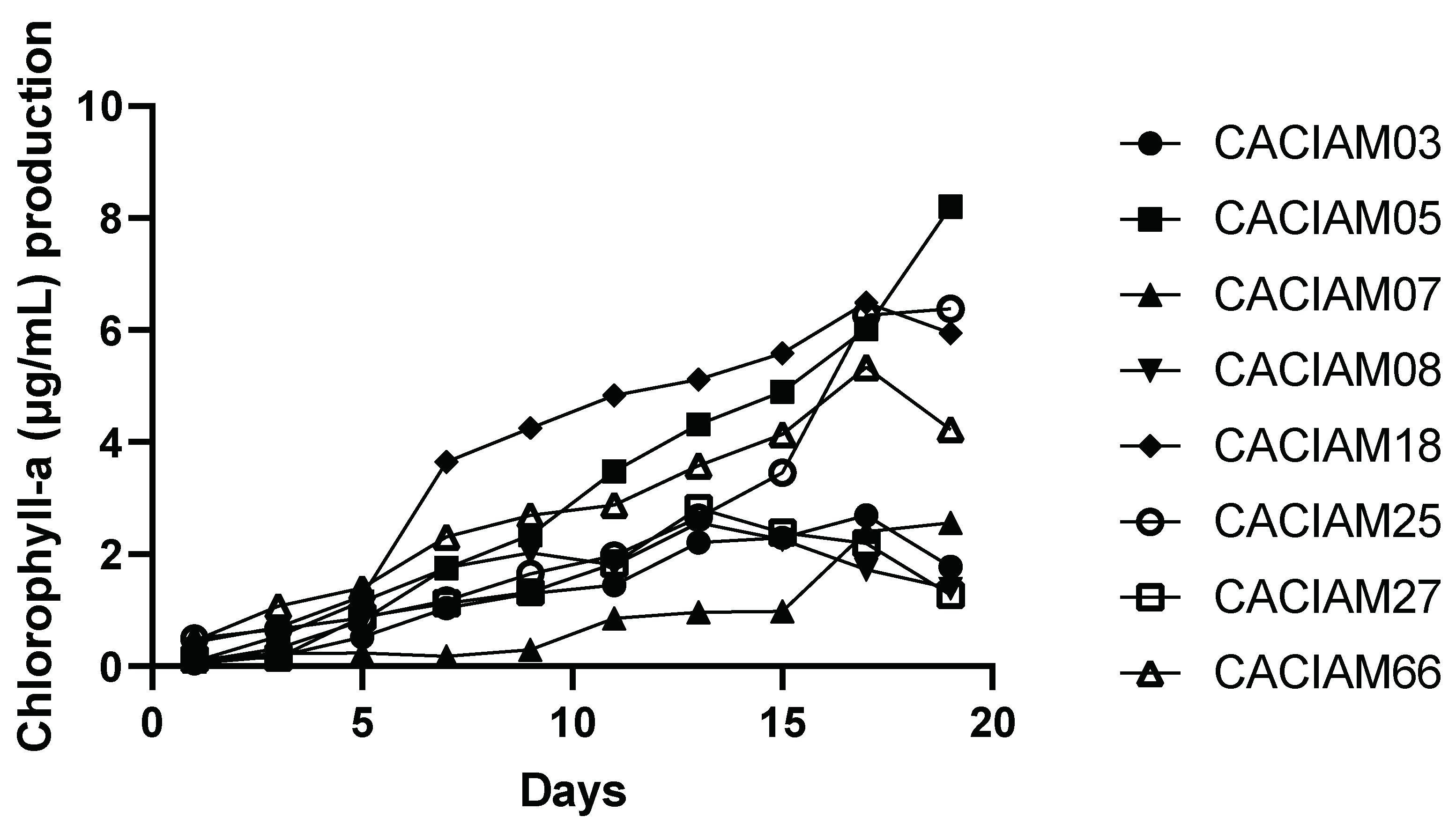
2.1. Selection of Strains by Determination of Chlorophyll a
2.2. Influence of Light Intensity and NaNO3 Concentration on Cyanobacteria Cultivation
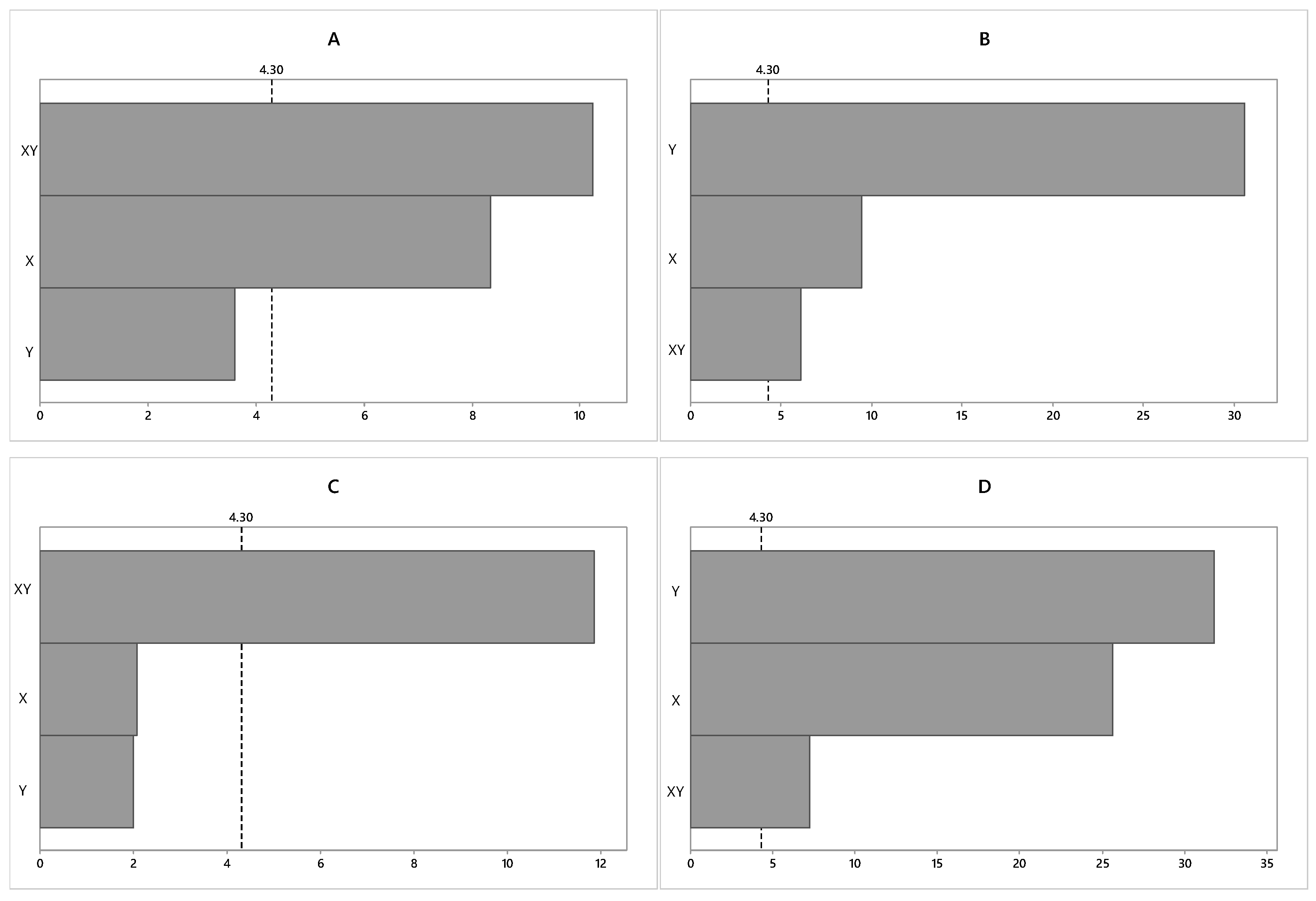
2.2.1. Statistical Analysis of the Factorial Planning of Strain Cultivation with Biomass Productivity as the Response Variable
2.2.2. Statistical Analysis of the Factorial Planning of Strain Cultivation with Lipid Content as the Response Variable
2.3. Empirical Parameters of the Biodiesel Quality from the Fatty Acid Profile
3. Materials and Methods
3.1. Obtaining the Strains and Cultivation
3.2. Measurement of Growth
3.2.1. Biomass Productivity (Determination of Chlorophyll a)
3.2.2. Biomass Productivity (Dry Biomass)
3.3. Optimization of Culture Conditions
3.4. Total Lipids Extraction and Fatty Acid Profile by Gas Chromatography (GC) Analysis
3.5. Biodiesel Properties Based on the Fatty Acid Profile
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Srinuanpan, S.; Cheirsilp, B.; Prasertsan, P.; Kato, Y.; Asano, Y. Strategies to increase the potential use of oleaginous microalgae as biodiesel feedstocks: Nutrient starvations and cost-effective harvesting process. Renew. Energy 2018, 122, 507–516. [Google Scholar] [CrossRef]
- Parmar, A.; Singh, N.K.; Pandey, A.; Gnansounou, E.; Madamwar, D. Cyanobacteria and microalgae: A positive prospect for biofuels. Bioresour. Technol. 2011, 102, 10163–10172. [Google Scholar] [CrossRef] [PubMed]
- Hankamer, B.; Lehr, F.; Rupprecht, J.; Mussgnug, J.H.; Posten, C.; Kruse, O. Photosynthetic biomass and H 2 production by green algae: from bioengineering to bioreactor scale-up. Physiol. Plant. 2007, 131, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Angermayr, S.A.; Hellingwerf, K.J.; Lindblad, P.; Teixeira de Mattos, M.J. Energy biotechnology with cyanobacteria. Curr. Opin. Biotechnol. 2009, 20, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Machado, I.M.P.; Atsumi, S. Cyanobacterial biofuel production. J. Biotechnol. 2012, 162, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Abed, R.M.M.; Dobretsov, S.; Sudesh, K. Applications of cyanobacteria in biotechnology. J. Appl. Microbiol. 2009, 106, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Da Rós, P.C.M.; Silva, C.S.P.; Silva-Stenico, M.E.; Fiore, M.F.; De Castro, H.F. Assessment of chemical and physico-chemical properties of cyanobacterial lipids for biodiesel production. Mar. Drugs 2013, 11, 2365–2381. [Google Scholar] [CrossRef]
- Lee, J.Y.; Yoo, C.; Jun, S.Y.; Ahn, C.Y.; Oh, H.M. Comparison of several methods for effective lipid extraction from microalgae. Bioresour. Technol. 2010, 101, 2009–2011. [Google Scholar] [CrossRef]
- Patel, A.; Arora, N.; Mehtani, J.; Pruthi, V.; Pruthi, P.A. Assessment of fuel properties on the basis of fatty acid profiles of oleaginous yeast for potential biodiesel production. Renew. Sustain. Energy Rev. 2017, 77, 604–616. [Google Scholar] [CrossRef] [Green Version]
- Bouaid, A.; Vázquez, R.; Martinez, M.; Aracil, J. Effect of free fatty acids contents on biodiesel quality. Pilot plant studies. Fuel 2016, 174, 54–62. [Google Scholar] [CrossRef]
- Giakoumis, E.G.; Sarakatsanis, C.K. Estimation of biodiesel cetane number, density, kinematic viscosity and heating values from its fatty acid weight composition. Fuel 2018, 222, 574–585. [Google Scholar] [CrossRef]
- Singh, V.; Chaudhary, D.K.; Mani, I.; Dhar, P.K. Recent advances and challenges of the use of cyanobacteria towards the production of biofuels. Renew. Sustain. Energy Rev. 2016, 60, 1–10. [Google Scholar] [CrossRef]
- Converti, A.; Casazza, A.A.; Ortiz, E.Y.; Perego, P.; Del Borghi, M. Effect of temperature and nitrogen concentration on the growth and lipid content of Nannochloropsis oculata and Chlorella vulgaris for biodiesel production. Chem. Eng. Process. Process Intensif. 2009, 48, 1146–1151. [Google Scholar] [CrossRef]
- He, Q.; Yang, H.; Hu, C. Optimizing light regimes on growth and lipid accumulation in Ankistrodesmus fusiformis H1 for biodiesel production. Bioresour. Technol. 2015, 198, 876–883. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.S.P.; Silva-Stenico, M.E.; Fiore, M.F.; de Castro, H.F.; Da Rós, P.C.M. Optimization of the cultivation conditions for Synechococcus sp. PCC7942(cyanobacterium) to be used as feedstock for biodiesel production. Algal Res. 2014, 3, 1–7. [Google Scholar] [CrossRef]
- Ruangsomboon, S. Effect of light, nutrient, cultivation time and salinity on lipid production of newly isolated strain of the green microalga, Botryococcus braunii KMITL 2. Bioresour. Technol. 2012, 109, 261–265. [Google Scholar] [CrossRef]
- Aboim, J.B.; Oliveira, D.; Ferreira, J.E.; Siqueira, A.S.; Dall’Agnol, L.T.; Rocha Filho, G.N.; Gonçalves, E.C.; Nascimento, L.A. Determination of biodiesel properties based on a fatty acid profile of eight Amazon cyanobacterial strains grown in two different culture media. RSC Adv. 2016, 6, 109751–109758. [Google Scholar] [CrossRef]
- Seyfabadi, J.; Ramezanpour, Z.; Khoeyi, Z.A. Protein, fatty acid, and pigment content of Chlorella vulgaris under different light regimes. J. Appl. Phycol. 2011, 23, 721–726. [Google Scholar] [CrossRef]
- Zili, F.; Mezhoud, N.; Trabelsi, L.; Chreif, I.; Ben Ouada, H. Fatty acid composition of the thermophilic Gloeocapsa gelatinosa under different combinations of temperature, light intensity, and NaNO3 concentration. J. Appl. Phycol. 2014, 27, 97–107. [Google Scholar] [CrossRef]
- Zulkifli, A.F.; Ramli, A.; Lim, J.W.; Lam, M.K. Effect of NaNO3 and NaCl concentration on Nannochloropsis oculata cell biomass and FAME composition for biodiesel production. J. Phys. Conf. Ser. 2018, 1123, 012071. [Google Scholar] [CrossRef]
- Sajjadi, B.; Chen, W.-Y.; Raman, A.A.A.; Ibrahim, S. Microalgae lipid and biomass for biofuel production: A comprehensive review on lipid enhancement strategies and their effects on fatty acid composition. Renew. Sustain. Energy Rev. 2019, 8, 55. [Google Scholar] [CrossRef]
- Sharma, J.; Kumar, S.S.; Bishnoi, N.R.; Pugazhendhi, A. Enhancement of lipid production from algal biomass through various growth parameters. J. Mol. Liq. 2018, 269, 712–720. [Google Scholar] [CrossRef]
- Vadivel, M.; Arvindnarayan, S.; Kumar, G.; Shobana, S.; Dharmaraja, J.; Nguyen, D.D.; Chang, S.W.; Prabhu, K.K.S. Biodiesel Potentiality of Microalgae Species: evaluation Using Various Nitrogen Sources. Waste Biomass Valor. 2019, 1–9. [Google Scholar] [CrossRef]
- Terra, D.; Oliveira, D.; Turbay, C.; Miguel, A.; Feitosa, T.; Barbosa, J.; De Nazaré, A.; Oliveira, D.; Pereira, L.; Silva, A.; et al. Lipid pro fi le analysis of three new Amazonian cyanobacteria as potential sources of biodiesel. Fuel 2018, 234, 785–788. [Google Scholar]
- Miao, X.; Wu, Q.; Yang, C. Fast pyrolysis of microalgae to produce renewable fuels. J. Anal. Appl. Pyrolysis 2004, 71, 855–863. [Google Scholar] [CrossRef]
- Da Rós, P.C.; Silva, C.S.; Silva-Stenico, M.E.; Fiore, M.F.; De Castro, H.F. Microcystis aeruginosa lipids as feedstock for biodiesel synthesis by enzymatic route. J. Mol. Catal. B Enzym. 2012, 84, 177–182. [Google Scholar] [CrossRef]
- Abirhire, O.; Kadiri, M. Growth and biodiesel production from the blue-green alga Microcystis aeruginosa Kutz. African J. E 2013, 33–39. [Google Scholar]
- Khan, M.I.; Lee, M.G.; Seo, H.J.; Shin, J.H.; Shin, T.S.; Yoon, Y.H.; Kim, M.Y.; Choi, J.I.; Kim, J.D. Enhancing the Feasibility of Microcystis aeruginosa as a Feedstock for Bioethanol Production under the Influence of Various Factors. Biomed Res. Int. 2016, 2016, 1–9. [Google Scholar]
- Khan, M.I.; Lee, M.G.; Shin, J.H.; Kim, J.D. Pretreatment optimization of the biomass of Microcystis aeruginosa for efficient bioethanol production. AMB Express 2017, 7. [Google Scholar] [CrossRef]
- Noreña-Caro, D.; Benton, M.G. Cyanobacteria as photoautotrophic biofactories of high-value chemicals. J. CO2 Util. 2018, 28, 335–366. [Google Scholar] [CrossRef]
- Mondal, M.; Ghosh, A.; Tiwari, O.N.; Gayen, K.; Das, P.; Mandal, M.K.; Halder, G. Influence of carbon sources and light intensity on biomass and lipid production of Chlorella sorokiniana BTA 9031 isolated from coalfield under various nutritional modes. Energy Convers. Manag. 2017, 145, 247–254. [Google Scholar] [CrossRef]
- Deng, X.; Chen, B.; Xue, C.; Li, D.; Hu, X.; Gao, K. Biomass production and biochemical profiles of a freshwater microalga Chlorella kessleri in mixotrophic culture: Effects of light intensity and photoperiodicity. Bioresour. Technol. 2019, 273, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Mandotra, S.K.; Kumar, P.; Suseela, M.R.; Nayaka, S.; Ramteke, P.W. Evaluation of fatty acid profile and biodiesel properties of microalga Scenedesmus abundans under the influence of phosphorus, pH and light intensities. Bioresour. Technol. 2016, 201, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Naira, V.R.; Das, D.; Maiti, S.K. Real time light intensity based carbon dioxide feeding for high cell-density microalgae cultivation and biodiesel production in a bubble column photobioreactor under outdoor natural sunlight. Bioresour. Technol. 2019, 284, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Yin, K.; Yuan, J.-P.; Cao, G.-X.; Xue, M.; Wu, C.-F.; Wang, J.-H. Characterization of a newly isolated green microalga Scenedesmus sp. as a potential source of biodiesel. Afr. J. Biotechnol. 2016, 11, 16083–16094. [Google Scholar]
- Cuellar-Bermudez, S.P.; Romero-Ogawa, M.A.; Vannela, R.; Lai, Y.J.S.; Rittmann, B.E.; Parra-Saldivar, R. Effects of light intensity and carbon dioxide on lipids and fatty acids produced by Synechocystis sp. PCC6803 during continuous flow. Algal Res. 2015, 12, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Renaud, S.M.; Van Thinh, L.; Lambrinidis, G.; Parry, D.L. Effect of temperature on growth, chemical composition and fatty acid composition of tropical Australian microalgae grown in batch cultures. Aquaculture 2002, 211, 195–214. [Google Scholar] [CrossRef]
- Economou, C.N.; Marinakis, N.; Moustaka-Gouni, M.; Kehayias, G.; Aggelis, G.; Vayenas, D.V. Lipid production by the filamentous cyanobacterium Limnothrix sp. growing in synthetic wastewater in suspended- and attached-growth photobioreactor systems. Ann. Microbiol. 2015, 65, 1941–1948. [Google Scholar] [CrossRef]
- Colla, L.M.; Bertolin, T.E.; Costa, J.A.V. Fatty Acids Profile of Spirulina platensis Grown under Different Temperatures and Nitrogen Concentrations. Zeitschrift fur Naturforsch. Sect. C J. Biosci. 2004, 59, 55–59. [Google Scholar] [CrossRef]
- Yaakob, Z.; Narayanan, B.N.; Padikkaparambil, S.; Unni K., S.; Akbar P., M. A review on the oxidation stability of biodiesel. Renew. Sustain. Energy Rev. 2014, 35, 136–153. [Google Scholar] [CrossRef]
- Ramos, M.J.; Fernández, C.M.; Casas, A.; Rodríguez, L.; Pérez, Á. Influence of fatty acid composition of raw materials on biodiesel properties. Bioresour. Technol. 2009, 100, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Srinuanpan, S.; Cheirsilp, B.; Prasertsan, P. Effective biogas upgrading and production of biodiesel feedstocks by strategic cultivation of oleaginous microalgae. Energy 2018, 148, 766–774. [Google Scholar] [CrossRef]
- Martínez, G.; Sánchez, N.; Encinar, J.M.; González, J.F. Fuel properties of biodiesel from vegetable oils and oil mixtures. Influence of methyl esters distribution. Biomass Bioenergy 2014, 63, 22–32. [Google Scholar] [CrossRef]
- Allen, M.M. Simple Conditions for Growth of Unicellular Blue-Green Algae on plates†. J. Phycol. 1967, 4, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Gorham, P.R.; McLachlan, J.; Hammer, U.T.; Kim, W.K. Isolation and culture of toxic strains of Anabaena flos-aquae (Lyngb.) de Bréb. Internationale Vereinigung für theoretische und angewandte Limnologie Verhandlungen 1964, 15, 796–804. [Google Scholar] [CrossRef]
- Meeks, J.C.; Castenholz, R.W. Growth and Photosynthesis in an Extreme Thermophile. Arch. Mikrobiol. 1971, 41, 25–41. [Google Scholar] [CrossRef]
- Fiore, M.F.; Moon, D.H.; Tsai, S.M.; Lee, H.; Trevors, J.T. Miniprep DNA isolation from unicellular and filamentous cyanobacteria. J. Microbiol. Methods 2000, 39, 159–169. [Google Scholar]
- Chatsungnoen, T.; Chisti, Y. Optimization of oil extraction from Nannochloropsis salina biomass paste. Algal Res. 2016, 15, 100–109. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. Canadian Journal of Biochemistry and Physiology. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay | Variable | Biomass Productivity (mg/L/day) | Lipid Content (%) | Lipid Productivity (mg/L/day) | ||||||||||||
| CACIAM 2 | CACIAM | CACIAM | ||||||||||||||
| Coded | Real 1 | 05 | 08 | 18 | 25 | 05 | 08 | 18 | 25 | 05 | 08 | 18 | 25 | |||
| X | Y | X | Y | |||||||||||||
| 1 | − | − | 15 | 1 | 26.8 | 12.1 | 23.2 | 24.2 | 15.3 | 12.4 | 20.6 | 33.1 | 4.1 | 3.8 | 4.8 | 3.4 |
| 2 | − | + | 15 | 2 | 37.4 | 15.3 | 33.2 | 42.1 | 18.9 | 31.7 | 32.9 | 14.4 | 7.1 | 2.3 | 10.9 | 24.6 |
| 3 | + | − | 100 | 1 | 39.5 | 16.3 | 34.7 | 39.0 | 25.6 | 15.0 | 37.9 | 58.3 | 10.1 | 7.2 | 13.2 | 11.1 |
| 4 | + | + | 100 | 2 | 48.4 | 18.4 | 45.8 | 53.7 | 17.9 | 44.0 | 20.7 | 28.4 | 8.7 | 3.3 | 9.5 | 7.4 |
| 5 | 0 | 0 | 57.5 | 1.5 | 34.6 | 9.5 | 34.2 | 30.5 | 18.1 | 18.1 | 13.7 | 7.8 | 6.3 | 1.7 | 4.7 | 2.4 |
| 6 | 0 | 0 | 57.5 | 1.5 | 36.1 | 10.2 | 34.0 | 29.7 | 18.7 | 18.9 | 14.9 | 8.5 | 6.7 | 1.9 | 5.1 | 2.5 |
| 7 | 0 | 0 | 57.5 | 1.5 | 35.8 | 9.2 | 34.6 | 31.6 | 17.6 | 17.3 | 12.4 | 7.0 | 6.3 | 1.6 | 4.3 | 2.2 |
| Strains | Fatty Acid | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C12:0 | C14:0 | C16:0 | C16:1 | C17:0 | C18:0 | C18:1 | C18:2 | C18:3 | C20:0 | SFA | MUFA | PUFA | |
| Synechocystis sp. CACIAM05 NaNO3 +/Light + | 9.16 | 7.08 | 54.96 | - | - | 9.62 | - | - | - | - | 80.82 | 0.00 | 0.00 |
| Synechocystis sp. CACIAM05 NaNO3 +/Light − | - | - | 29.90 | 2.13 | 2.13 | - | 42.65 | - | - | - | 32.03 | 44.78 | 0.00 |
| Synechocystis sp. CACIAM05 NaNO3 −/Light + | - | - | 37.65 | 2.25 | 2.27 | - | 28.15 | - | - | - | 39.92 | 30.41 | 0.00 |
| Synechocystis sp. CACIAM05 NaNO3 −/Light − | - | - | 37.77 | 2.01 | 2.06 | - | 20.55 | - | - | - | 39.83 | 22.56 | 0.00 |
| Synechocystis sp. CACIAM05 NaNO3 0/Light 0 | - | - | 60.52 | - | - | 2.62 | - | 14.65 | 2.96 | - | 63.14 | 0.00 | 17.61 |
| M. aeruginosa CACIAM08 NaNO3 +/Light + | - | - | 29.97 | - | - | 15.32 | 49.50 | - | - | 1.33 | 46.62 | 49.50 | 0.00 |
| M. aeruginosa CACIAM08 NaNO3 +/Light − | - | - | 25.71 | - | 28.36 | - | 43.90 | - | - | - | 54.07 | 43.90 | 0.00 |
| M. aeruginosa CACIAM08 NaNO3 −/Light + | - | - | - | - | 23.77 | 5.69 | 5.81 | - | - | - | 29.47 | 5.81 | 0.00 |
| M. aeruginosa CACIAM08 NaNO3 −/Light − | - | 1.62 | 33.14 | - | 11.25 | 11.20 | 15.42 | - | - | 1.39 | 58.61 | 15.42 | 0.00 |
| M. aeruginosa CACIAM08 NaNO3 0/Light 0 | - | - | 31.18 | - | 20.67 | 3.95 | 29.87 | - | - | - | 55.80 | 29.87 | 0.00 |
| Pantanalinema rosaneae CACIAM18 NaNO3 +/Light + | - | 9.92 | 32.99 | 34.13 | - | - | 4.59 | - | - | - | 42.92 | 38.72 | 0.00 |
| Pantanalinema rosaneae CACIAM18 NaNO3 +/Light − | - | 9.91 | 25.22 | 52.55 | - | - | - | - | - | - | 35.13 | 52.55 | 0.00 |
| Pantanalinema rosanea CACIAM18 NaNO3 −/Light + | - | 18.58 | 29.66 | 23.79 | - | - | - | - | - | - | 48.23 | 23.79 | 0.00 |
| Pantanalinema rosaneae CACIAM18 NaNO3 −/Light − | - | 14.91 | 32.74 | 42.96 | - | 2.42 | - | - | - | - | 50.06 | 42.957 | 0.00 |
| Pantanalinema rosaneae CACIAM18 NaNO3 0/Light 0 | - | 18.10 | 29.86 | 25.51 | - | - | 7.79 | - | - | - | 47.97 | 33.30 | 0.00 |
| Limnothrix sp. CACIAM25 NaNO3 +/Light + | - | 3.59 | 22.12 | 5.11 | 20.54 | 9.44 | 33.33 | - | - | 1.15 | 56.83 | 38.44 | 0.00 |
| Limnothrix sp. CACIAM25 NaNO3 +/Light − | - | - | 39.83 | - | 12.72 | 8.97 | 38.49 | - | - | - | 61.51 | 38.49 | 0.00 |
| Limnothrix sp. CACIAM25 NaNO3 −/Light + | - | 10.62 | 43.29 | 20.93 | - | 3.47 | - | 13.05 | - | - | 57.38 | 20.93 | 13.05 |
| Limnothrix sp. CACIAM25 NaNO3 −/Light − | - | 1.62 | 33.14 | - | 11.25 | 11.20 | 15.42 | - | - | 1.39 | 48.15 | 33.08 | 0.00 |
| Limnothrix sp. CACIAM25 NaNO3 0/Light 0 | - | - | 23.39 | 3.43 | 29.45 | - | 32.88 | - | - | - | 52.84 | 36.31 | 0.00 |
| Strains | DU | SV (mg/g) | IV | CN | LCSF | CFPP (°C) | CP (°C) | PP (°C) | APE | BAPE | HHV | ν (mm²/s) | ρ (g/cm³) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CACIAM05 NaNO3 +/Light + | 0.0 | 181.9 | 0.0 | 76.3 | 10.3 | 15.9 | 23.9 | 19.1 | 0.0 | 0.0 | 31.5 | 2.8 | 0.7 |
| CACIAM05 NaNO3 +/Light − | 44.8 | 159.0 | 40.5 | 71.5 | 3.0 | −7.1 | 10.7 | 4.8 | 42.7 | 0.0 | 30.3 | 2.9 | 0.7 |
| CACIAM05 NaNO3 −/Light + | 30.4 | 147.7 | 27.6 | 77.1 | 3.8 | −4.7 | 14.8 | 9.3 | 28.2 | 0.0 | 27.7 | 2.6 | 0.6 |
| CACIAM05 NaNO3 −/Light − | 22.6 | 131.9 | 20.5 | 83.1 | 3.8 | −4.6 | 14.9 | 9.3 | 20.6 | 0.0 | 24.5 | 2.3 | 0.5 |
| CACIAM05 NaNO3 0/Light 0 | 35.2 | 172.5 | 34.6 | 70.1 | 7.4 | 6.7 | 26.8 | 22.3 | 35.2 | 20.6 | 31.7 | 2.9 | 0.7 |
| CACIAM08 NaNO3 +/Light + | 49.5 | 196.2 | 44.5 | 64.1 | 12.0 | 21.2 | 10.8 | 4.9 | 49.5 | 2.7 | 38.0 | 3.9 | 0.8 |
| CACIAM08 NaNO3 +/Light − | 43.9 | 201.9 | 39.5 | 64.5 | 2.6 | −8.4 | 8.5 | 2.4 | 43.9 | 0.0 | 38.7 | 4.0 | 0.9 |
| CACIAM08 NaNO3 −/Light + | 5.8 | 72.0 | 5.2 | 121.0 | 2.9 | −7.5 | −5.0 | −12.2 | 5.8 | 0.0 | 14.0 | 1.7 | 0.3 |
| CACIAM08 NaNO3 −/Light − | 15.4 | 154.9 | 13.9 | 78.4 | 10.3 | 15.9 | 12.4 | 6.7 | 15.4 | 2.8 | 29.2 | 2.9 | 0.6 |
| CACIAM08 NaNO3 0/Light 0 | 29.9 | 177.9 | 26.9 | 70.9 | 5.1 | −0.5 | 11.4 | 5.6 | 29.9 | 0.0 | 33.8 | 3.3 | 0.7 |
| CACIAM18 NaNO3 +/Light + | 38.7 | 180.6 | 38.2 | 67.9 | 3.3 | −6.1 | 12.4 | 6.6 | 4.6 | 0.0 | 31.8 | 2.7 | 0.7 |
| CACIAM18 NaNO3 +/Light − | 52.6 | 195.0 | 52.5 | 62.5 | 2.5 | −8.6 | 8.3 | 2.2 | 0.0 | 0.0 | 34.1 | 2.8 | 0.8 |
| CACIAM18 NaNO3 −/Light + | 23.8 | 162.7 | 23.8 | 74.5 | 3.0 | −7.2 | 10.6 | 4.7 | 0.0 | 0.0 | 28.0 | 2.4 | 0.6 |
| CACIAM18 NaNO3 −/Light − | 43.0 | 207.4 | 42.9 | 63.0 | 4.5 | −2.4 | 12.2 | 6.5 | 0.0 | 0.0 | 36.2 | 3.1 | 0.8 |
| CACIAM18 NaNO3 0/Light 0 | 33.3 | 181.2 | 32.5 | 69.1 | 3.0 | −7.1 | 10.7 | 4.8 | 7.8 | 0.0 | 31.7 | 2.7 | 0.7 |
| CACIAM25 NaNO3 +/Light + | 38.4 | 197.7 | 35.1 | 66.0 | 8.1 | 8.9 | 6.6 | 0.4 | 33.3 | 2.3 | 37.6 | 3.8 | 0.8 |
| CACIAM25 NaNO3 +/Light − | 38.5 | 207.3 | 34.6 | 64.9 | 8.5 | 10.1 | 16.0 | 10.5 | 38.5 | 0.0 | 39.5 | 4.1 | 0.9 |
| CACIAM25 NaNO3 −/Light + | 47.0 | 199.5 | 44.5 | 63.6 | 6.1 | 2.6 | 17.8 | 12.5 | 26.1 | 13.1 | 35.7 | 3.1 | 0.8 |
| CACIAM25 NaNO3 −/Light − | 33.1 | 167.7 | 30.1 | 72.1 | 6.3 | 3.4 | 6.8 | 0.5 | 30.0 | 0.0 | 32.1 | 3.2 | 0.7 |
| CACIAM25 NaNO3 0/Light 0 | 36.3 | 184.8 | 33.0 | 68.4 | 2.3 | −9.1 | 7.3 | 1.1 | 32.9 | 0.0 | 35.2 | 3.5 | 0.8 |
| Biodiesel Quality Specifications | |||||||||||||
| ABNT NBR | Report | Report | Country specific | - | - | 3.0 (min.)/6.0 (max.) | - | ||||||
| EU EN 14214 | 120 (max.) | 51 (min.) | Country specific | - | Country specific | 3.5 (min.)/5.0 (max.) | 0.86 (min.)/0.90 (max.) | ||||||
| USA ASTM D6751 | - | 47 (min.) | - | Report | - | 1.9 (min.)/6.0 (max.) | - | ||||||
| Variable | Level | |||
|---|---|---|---|---|
| Independent Variables | Symbol | - | 0 | + |
| Light Intensity (µmol photons m−2 s−1) | X | 15 | 57.5 | 100 |
| NaNO3 (g/L) | Y | 1.0 | 1.5 | 2.0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aboim, J.B.; Oliveira, D.T.d.; Mescouto, V.A.d.; dos Reis, A.S.; da Rocha Filho, G.N.; Santos, A.V.; Xavier, L.P.; Santos, A.S.; Gonçalves, E.C.; do Nascimento, L.A.S. Optimization of Light Intensity and NaNO3 Concentration in Amazon Cyanobacteria Cultivation to Produce Biodiesel. Molecules 2019, 24, 2326. https://doi.org/10.3390/molecules24122326
Aboim JB, Oliveira DTd, Mescouto VAd, dos Reis AS, da Rocha Filho GN, Santos AV, Xavier LP, Santos AS, Gonçalves EC, do Nascimento LAS. Optimization of Light Intensity and NaNO3 Concentration in Amazon Cyanobacteria Cultivation to Produce Biodiesel. Molecules. 2019; 24(12):2326. https://doi.org/10.3390/molecules24122326
Chicago/Turabian StyleAboim, Joseline Barbosa, Deborah Terra de Oliveira, Vanessa Albuquerque de Mescouto, André Silva dos Reis, Geraldo Narciso da Rocha Filho, Agenor Valadares Santos, Luciana Pereira Xavier, Alberdan Silva Santos, Evonnildo Costa Gonçalves, and Luis Adriano Santos do Nascimento. 2019. "Optimization of Light Intensity and NaNO3 Concentration in Amazon Cyanobacteria Cultivation to Produce Biodiesel" Molecules 24, no. 12: 2326. https://doi.org/10.3390/molecules24122326