2.2.1. Effect of Heavy Metals (Silver and Cadmium)
Elicitation of C. officinalis hairy roots with Ag+ and Cd2+ ions (supplied as silver nitrate AgNO3, and cadmium chloride CdCl2) was performed according to the procedure described in chapter 3.2.1.
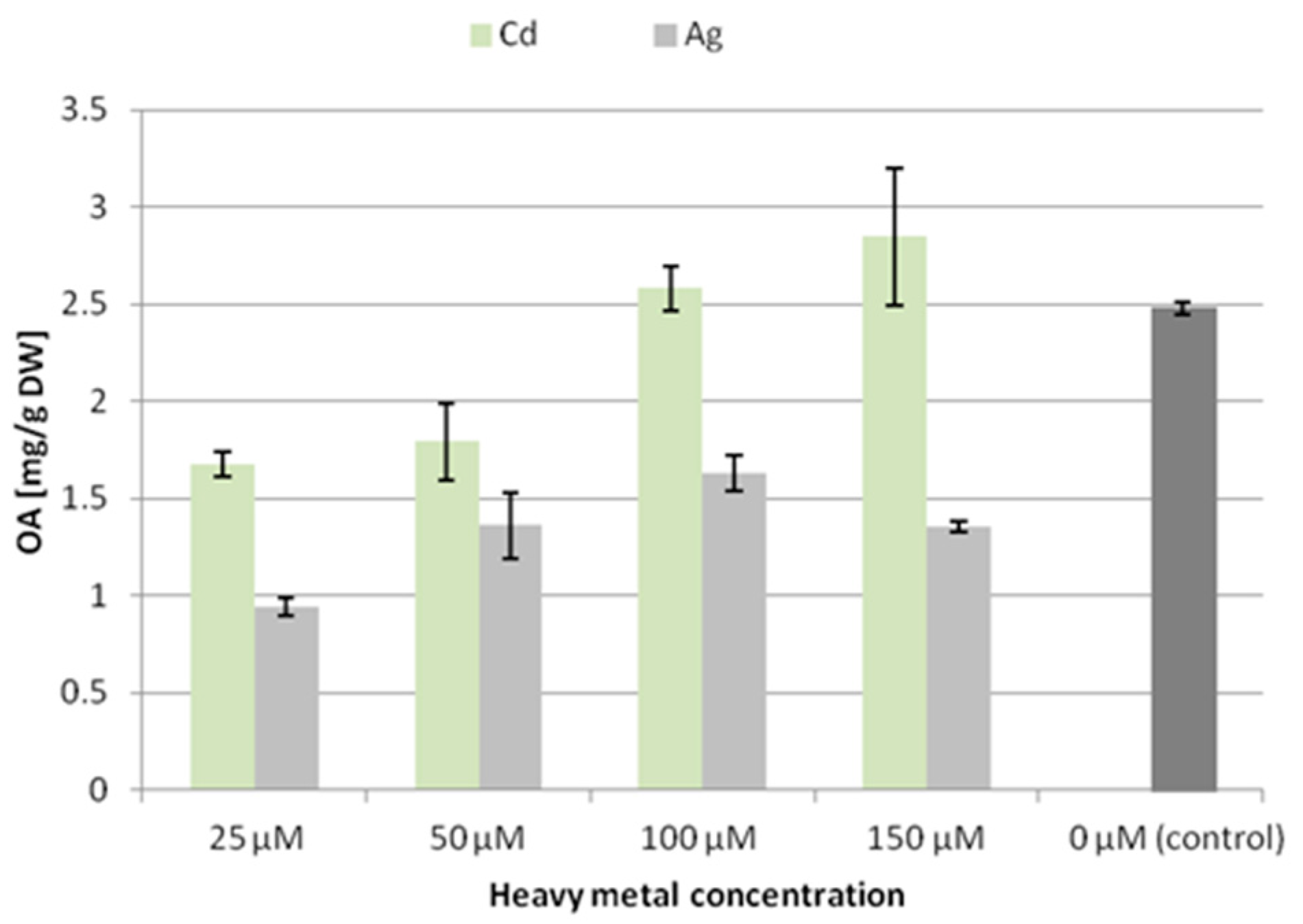
The CC16 line was treated by four concentrations of silver nitrate and cadmium chloride: 25, 50, 100, and 150 μM. All concentrations significantly reduced both the fresh mass and dry weight of the roots (by 47–77% in accordance with increasing concentrations, see
Figures S1 and S2). The highest concentration (150 μM) of both salts exerted a visibly harmful effect on the hairy roots, which became very dark and stopped growing. Therefore, the CH2 line was treated only by concentrations of 25, 50, and 100 μM. The CH2 line seemed to be less sensitive to the influence of Cd
2+ ions, at least at the lowest applied concentration (25 µM), where no decrease in the fresh or dry mass was noted. However, at higher concentrations, particularly 100 µM, the decrease in both fresh and dry mass reached 50–66%. It should be emphasized that even low concentrations of heavy metals led to significant changes in the root morphology. The roots became darker, thicker, and significantly less branched, particularly after Ag
+ treatment (
Figure S3).
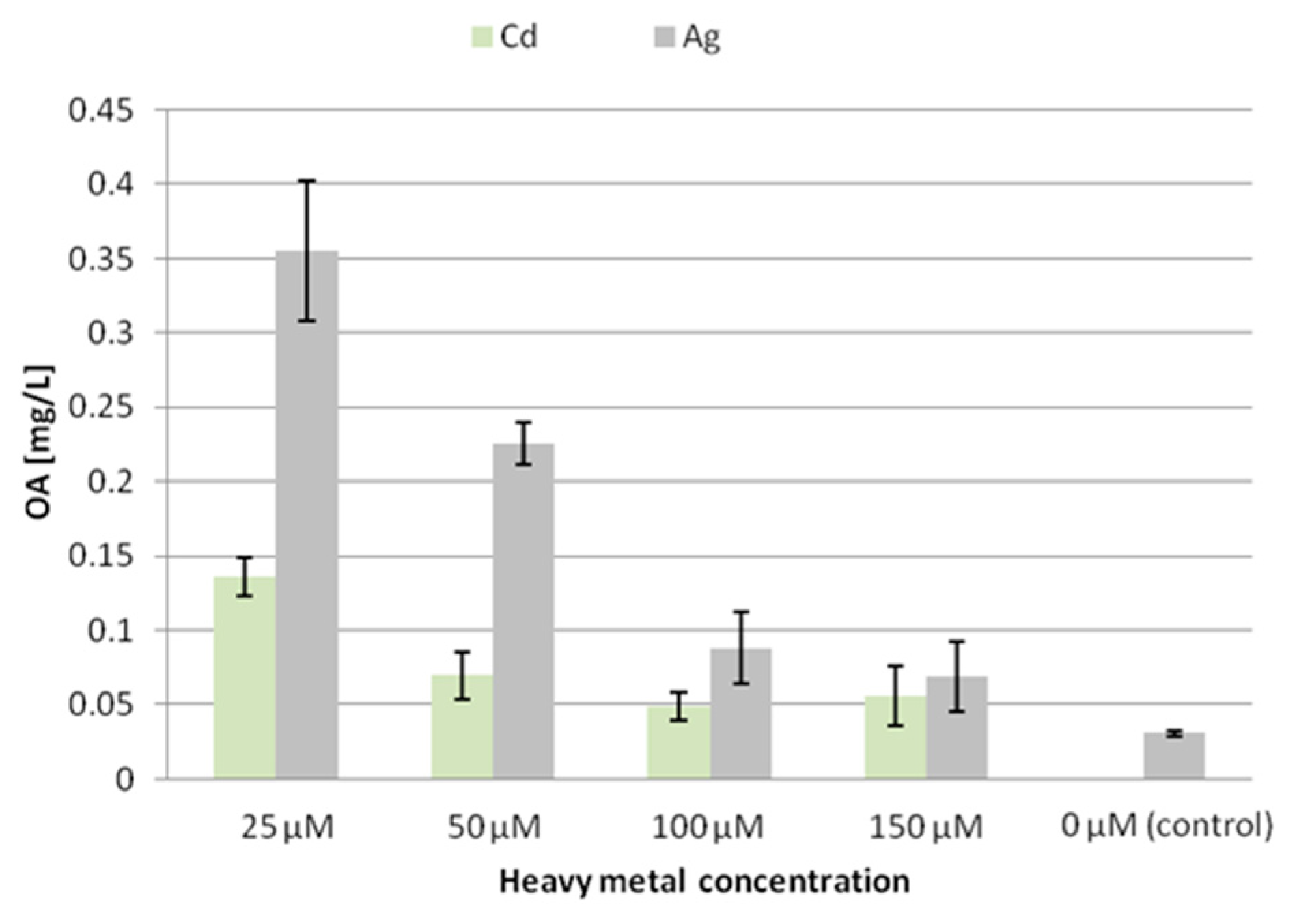
Elicitation with the tested heavy metal ions exerted diverse effects on the accumulation of oleanolic acid glycosides in the tissue of the two hairy root lines.
In the CC16 line, the content of oleanolic acid (released from hydrolyzed saponins) decreased significantly at all the applied concentrations of silver ions, with the lowest level (decreased by 62% compared to the control) after treatment with 25 µM Ag
+. In contrast, in samples treated with cadmium ions, the content of oleanolic acid decreased only at the two low concentrations, whereas it increased by 15% after treatment with 150 µM Cd
2+ (
Figure 2).
The secretion of oleanolic acid glycosides from the CC16 line hairy roots increased after treatment with both tested heavy metals, although much more significantly after elicitation with silver ions (
Figure 3). The strongest effect was observed at a concentration of 25 μM, where the amount of oleanolic acid secreted to the culture medium increased 5-fold and 12-fold after treatment with Cd
2+ or Ag
+, respectively. In turn, Cd
2+ ions at concentrations of 100 and 150 μM did not show statistically significant increases in oleanolic acid in the culture medium. The lowest secretion of oleanolic acid saponins after elicitation with Ag
+ ions (however, still more than 2-fold) was observed at a concentration 150 μM. Thus, the influence of silver ions on the secretion of oleanolic acid glycosides in the line CC16 was found to be inversely concentration-dependent.
The strong stimulation of the secretion of oleanolic acid glycosides exerted by the lower concentrations of both applied heavy metal ions is correlated with the lowest levels of these compounds accumulated in hairy root tissue. Therefore, the decrease in the content of oleanolic acid in the tissue can be explained by their increased secretion to the surrounding medium. At higher concentrations of heavy metal ions, the secretion of oleanolic acid glycosides was not as intensive, and thus a greater amount of these compounds was accumulated in the tissue. This correlation is particularly remarkable in the case of cadmium ions.
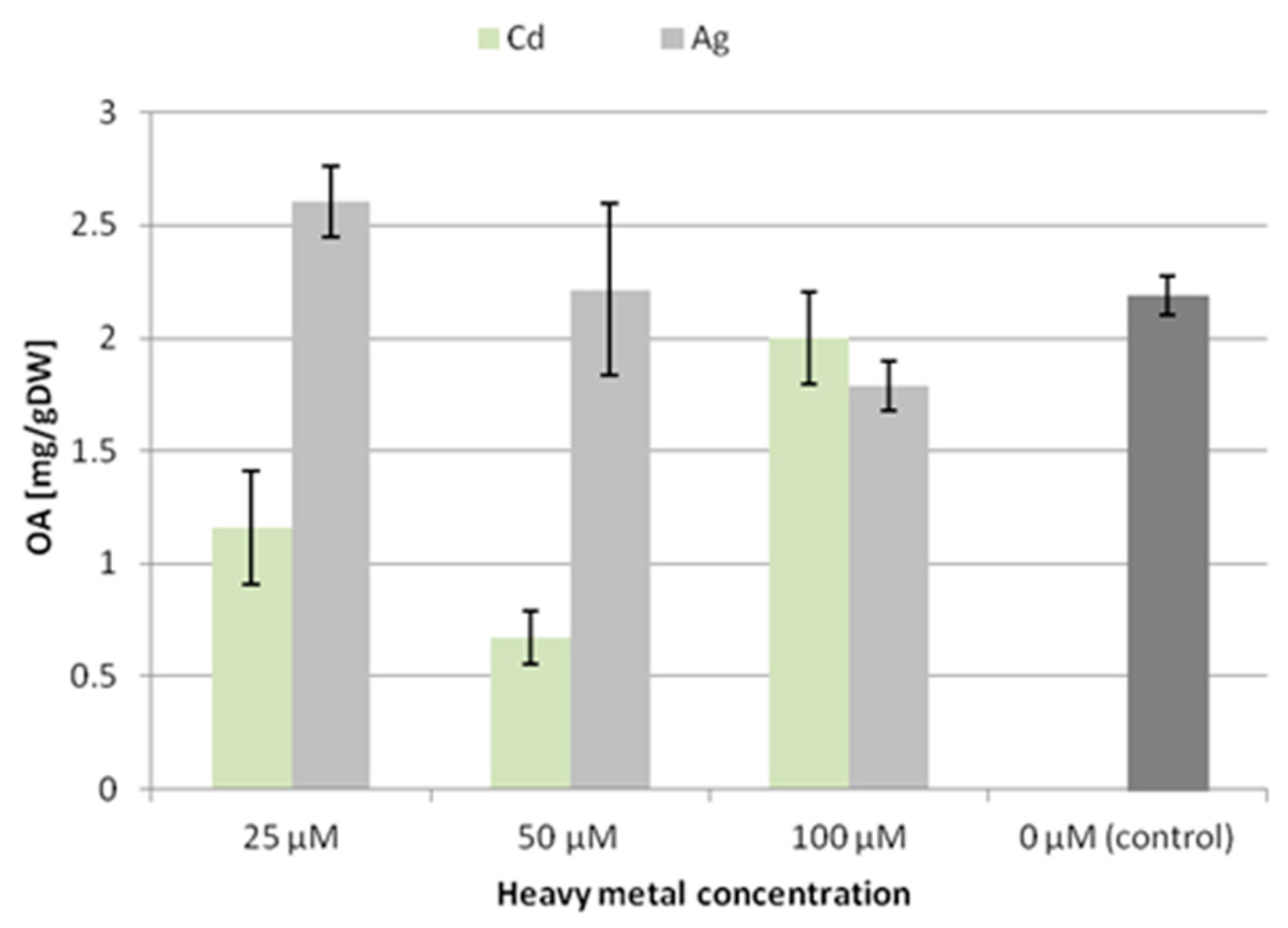
The trends of accumulation and secretion of oleanolic acid glycosides after treatment with heavy metals noted in the CH2 line (
Figure 4 and
Figure 5) differ in some aspects from those observed for the CC16 line.
After treatment with silver ions, the content of saponins accumulated in the CH2 line root tissue increased at the lowest applied concentration (25 μM Ag+) by 14% and decreased by 18% at 100 μM Ag+. In turn, after treatment with Cd2+ ions, the content of saponins accumulated in the root tissue decreased, particularly at the two lower concentrations, i.e., almost by 50% at both 25 and 50 μM Cd2+. Thus, the accumulation of saponins in the CH2 line hairy roots was affected more strongly by Cd2+ ions than Ag+ ions, and these effects were opposite to those observed for the CC16 line. It can be concluded that the two lines of hairy roots, CC16 and CH2, significantly differ in their sensitivity to the two applied metal ions regarding the effect exerted on their accumulation of oleanolic acid glycosides.
Elicitation with Ag+ or Cd2+ ions increased the secretion of oleanolic acid glycosides in the CH2 line at all the applied concentrations, but with opposite tendencies. The saponin secretion was increased 4.5-fold by comparison to the control at 25 μM Cd2+, and almost 8-fold at 100 μM Cd2+. Conversely, the secretion increased almost 5.5-fold after treatment with 25 μM Ag+, and only 4-fold at 100 μM Ag+. Thus, the enhancement of secretion of oleanolic acid glycosides in the CH2 line was directly proportional to growing cadmium ion concentration, whereas it was inversely proportional to silver ion concentration.
A similar inverse relationship between saponin secretion and silver ion concentration was also observed for the CC16 line. Thus, the influence of the tested heavy metal ions on saponin secretion in the two hairy root lines differed, particularly with respect to the cadmium ion concentration.
The content of sterols in the CC16 line after elicitation with silver ions decreased by almost 50% compared to the control at a concentration of 25 µM Ag
+ (
Table 1). According to the results described above, this decline in sterol biosynthesis and accumulation was correlated with the reduced accumulation of oleanolic acid glycosides compensated by a higher level of saponin secretion. With growing concentration of supplied silver ions, the content of sterols progressively increased, exceeding the control level by 22% at 150 µM Ag
+ (however, this concentration was highly detrimental to hairy root growth). Simultaneously, the ratio of the two most abundant sterols, stigmasterol and sitosterol, was remarkably changed, equaling 2.5:1 for the control and 0.9:1 after treatment with 50 μM Ag
+, where sitosterol was the most abundant sterol instead of stigmasterol.
In contrast to the effect exerted by Ag
+, all the applied concentrations of Cd
2+ increased the sterol accumulation in the CC16 line hairy roots (
Table 2). This effect was directly concentration-dependent, from a 19% increase at 25 μM Cd
2+ up to a level of 150% of the control at 150 μM Cd
2+. The amount of predominant stigmasterol was similar at all the tested Cd
2+ concentrations (on average 30% higher than the control level), whereas the amount of sitosterol decreased significantly (to 50% of the control) at the lowest applied concentration of Cd
2+. Thus, the ratio of the stigmasterol and sitosterol was again substantially changed, equaling 6.6:1 at 25 μM Cd
2+ and 2.2:1 at 150 μM Cd
2+.
After elicitation with silver ions, the content of sterols in the CH2 line decreased by approximately 30% compared to the control at all the applied concentrations of Ag
+ (
Table 3). The ratio of stigmasterol to sitosterol was changed, equaling 1.4:1 in the control and approximately 0.9:1 in all the Ag-treated hairy root samples, so sitosterol became the predominant sterol instead of stigmasterol.
In contrast to the influence of cadmium exerted on the CC16 line, where the content of sterols was elevated in comparison to the control, with the CH2 line a decline in sterol accumulation was observed, with the lowest level (a 33% decrease compared to the control) at the concentration of 25 μM Cd
2+ (
Table 4). Similarly as described above, sitosterol became the prevailing compound, with stigmasterol: sitosterol ratios of 0.8:1 at 25 and 50 μM Cd
2+, and 0.6:1 at 100 μM Cd
2+.
Various applications of heavy metals as abiotic elicitors enhancing the productivity of in vitro cultures have been reported so far. For example, silver and cadmium ions at a concentration of 25 μM increased considerably (more than 11-fold) the content of diterpene quinines (referred to as tashinones) in
Salvia miltiorrhiza cell suspension cultures [
20]. Silver nitrate at a concentration of 1mM increased by 5- and 8-fold the accumulation of scopolamine and hyoscyamine, respectively, in hairy root cultures of
Brugmansia candida; moreover, it stimulated a 3-fold increase in the release of scopolamine into the medium [
21]. However, the toxic influence of high concentrations of heavy metal ions cannot be neglected, since sometimes the inhibition of hairy root growth was observed, and it was associated with a decrease in the accumulation of metabolites and an increase in their release. The higher release of compounds to the medium could be due to damage to the cell membrane or to cellular lysis, appearing as a consequence of heavy metal toxicity [
21].
Summarizing the results obtained in the present study, it has to be admitted that the influence of silver and cadmium ions exerted various concentration-dependent effects that were different in the two tested hairy root lines. Generally, the accumulation of oleanolic acid glycosides in the hairy root tissue of both lines was reduced, whereas the main phenomenon observed as a response to heavy metal treatment was the strong stimulation (up to 12-fold) of the secretion of oleanolic acid saponins. The influence of both supplied heavy metal ions on sterol biosynthesis was particularly specific. In the CC16 line, the total content of sterols increased (up to 22%) for all the applied concentrations of Cd2+ and the two highest (100 and 150 μM) concentrations of Ag+. Simultaneously, the ratio of stigmasterol to sitosterol was significantly changed, with the growing amount of sitosterol finally exceeding the predominant stigmasterol. In the CH2 line, the total content of sterols declined for all applied Ag+ and Cd2+ concentrations; however, the phenomenon of the increased biosynthesis of sitosterol and its prevalence over stigmasterol was also observed. Thus, heavy metals seem to deeply disturb the triterpenoid biosynthesis pathway, since their influence includes modifications concerning not only the switch between sterols and pentacyclic compounds, but also significant changes in the sterol profile. Nevertheless, the advantage of the use of heavy metals as elicitors is debatable due to their harmful influence on hairy root growth and the visible changes in the roots’ morphology and branching.
2.2.2. Effect of Ultrasound
Elicitation of hairy roots with ultrasound was performed according to the procedure described in chapter 3.2.2. After exposure to ultrasound, the hairy root samples were cultured during the subsequent 7 days. The fresh weight of both lines of hairy roots exposed to a 10 min treatment with ultrasound increased by 11% and 8% for the CC16 line and CH2 line, respectively. After 20 min of exposure, the fresh weight of the CC16 line roots remained the same as the control, whereas the fresh weight of the CH2 line roots decreased by 40% (
Figure S4).
The dry weight increased by approximately 7% for the CH2 root line exposed to 10 min of ultrasound treatment, and it remained practically unchanged for the CC16 line. The dry weight of hairy roots exposed to 20 min ultrasound elicitation decreased, particularly remarkably (by 47%) for the CH2 line (
Figure S5). Ultrasound treatment changed the color of the roots to a darker shade for both the CC16 and CH2 lines, particularly in samples after 20 min exposure (
Figure S6).
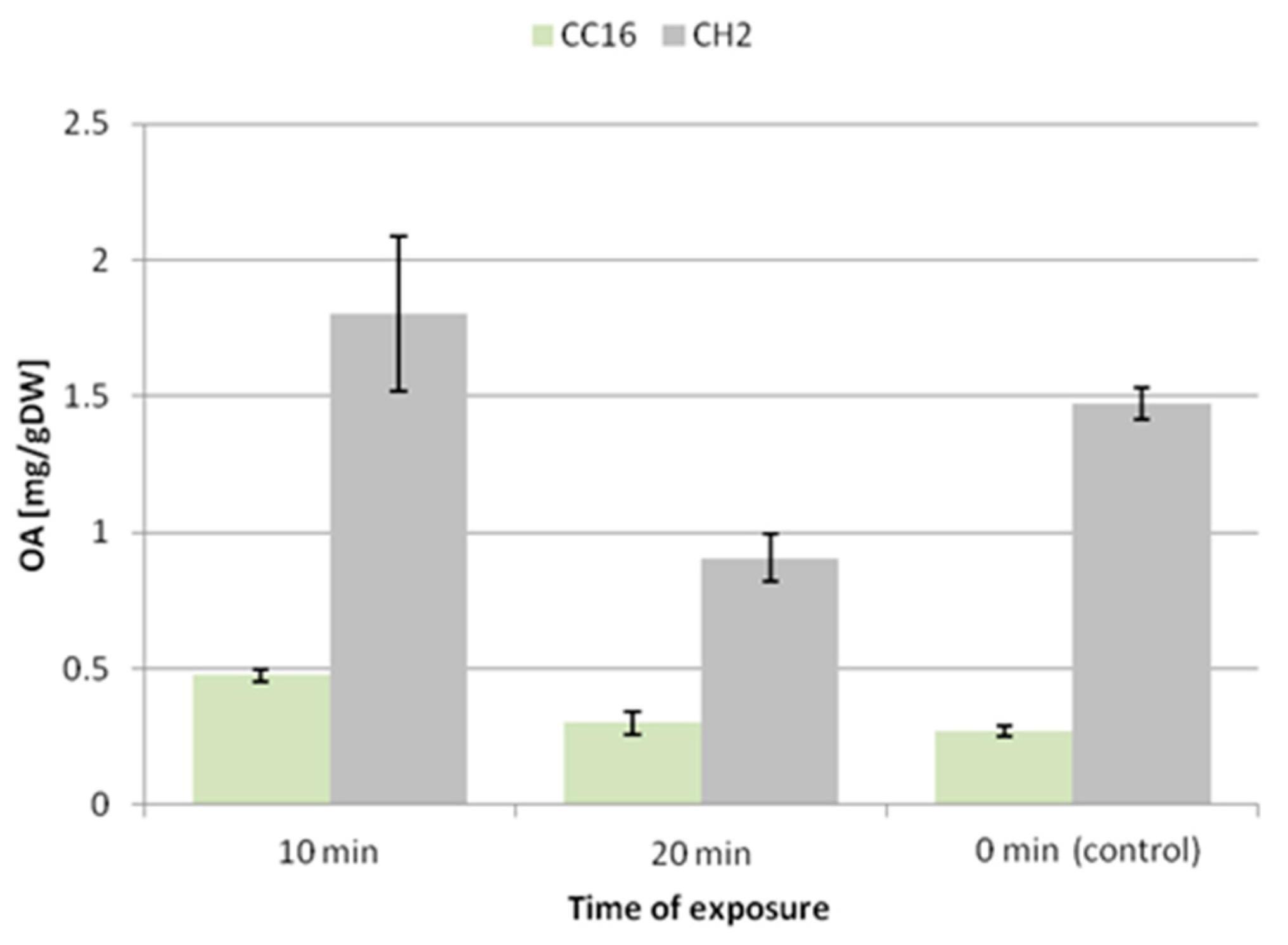
The 10 min exposure to ultrasound caused an increase in the accumulation of oleanolic acid glycosides by 50% and 64% in the hairy root tissue of the CC16 and CH2 lines, respectively. In turn, the 20 min exposure did not exert a significant effect on the CC16 line, whereas it decreased the level of saponins by 26% in the CH2 line (
Figure 6).
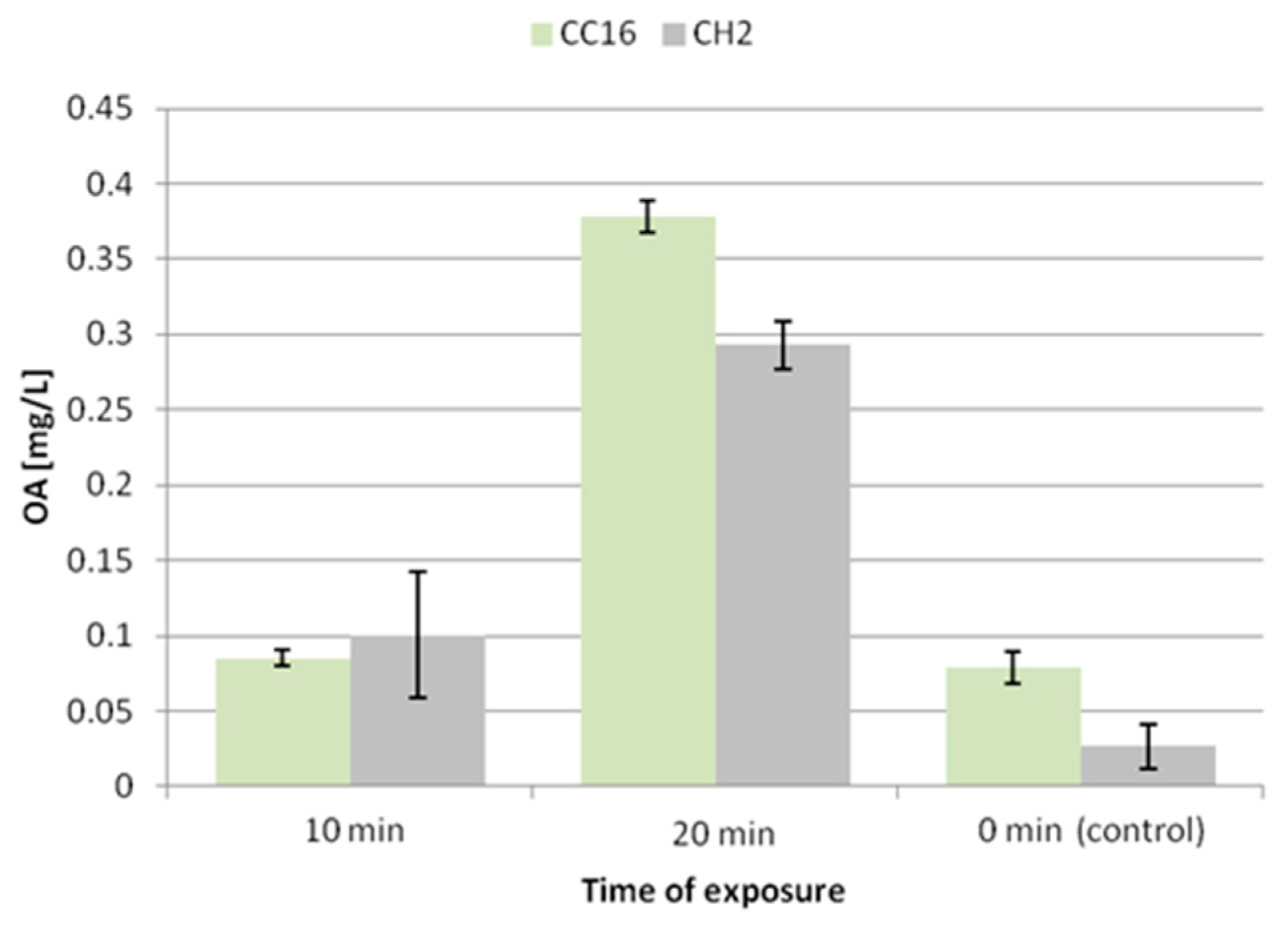
After 10 min exposure to ultrasound, the secretion of oleanolic acid glycosides did not remarkably change in the hairy root CC16 line, while it increased 3.8-fold in the CH2 line (
Figure 7). A significant enhancement of the secretion was noted after 20 min exposure, 4-fold in the CC16 line and 11-fold in CH2 line. Thus, the elevated secretion of saponins after 20 min exposure to ultrasound is correlated with the reduction of the accumulation of these compounds in hairy root tissue.
The content of sterols increased in the hairy roots of both lines exposed to 10 min of ultrasound treatment, by 7% and 11% in the CC16 and CH2 lines, respectively, and it significantly decreased by 38% and 47% after 20 min of ultrasound treatment, without changing the ratio among individual compounds (
Table 5 and
Table 6).
Ultrasound has been often applied as a physical elicitor for stimulation and manipulation of cells, tissues, and organs in various plant in vitro techniques. It was reported that low-energy ultrasound enhanced the synthesis and secretion of shikonin to the culture media from a suspension culture of
Lithospermum erythrorhizon [
22]. Therefore, ultrasound was regarded as working in two ways: it increased the membrane permeability—shikonin was excreted to the medium 3.5-times more effectively, and it stimulated the activity of key enzymes responsible for shikonin synthesis, i.e., phenylalanine ammonia lyase and
p-hydroxybenzoic acid geranyltransferase. The selection of appropriate parameters is crucial for reducing the level of mechanical damage and for permitting the reversible solubilization of various cellular membranes.
Nevertheless, ultrasound does not always improve the exudation of metabolites. Safari et al. (2012) reported a remarkable enhancement in the production of taxanes in hazel (
Corylus avellana) suspension culture elicited by ultrasound, with no parallel increase in their release through the cell membrane [
23]. Likewise, the production of cichoric, caftaric, chlorogenic, and caffeic acid was enhanced in ultrasound-elicitated
Echinacea purpurea hairy root culture; however, these compounds were not detected in the liquid medium [
24].
The present study revealed that, in the case of C. officinalis hairy root cultures, the effect of ultrasound of 50 kHz frequency markedly depended on the time of exposure. A short-time exposure caused an enhancement in the accumulation of oleanolic acid glycosides in the root tissues (50%–64%), accompanied by a slight increase in sterol content (7%–12%), whereas the secretion of saponins did not change in the CC16 line and increased almost 4-fold in the CH2 line. A longer exposure caused a decline in the level of saponins only in the tissue of the hairy root CH2 line, while in the less sensitive CC16 line it remained comparable to the control. 20 min of ultrasound treatment significantly stimulated the secretion of oleanolic acid saponins (4-fold in the CC16 line and 11-fold in the CH2 line), simultaneously decreasing the content of sterols (by 38%–46%). No changes in the ratio among individual sterols were noted.
2.2.3. Effect of UV radiation
Elicitation of hairy roots with UV-C radiation was performed according to the procedure described in chapter 3.2.3. After every exposure to UV radiation, the hairy root samples were cultured during the subsequent 7 days. Thirty minutes of UV exposure resulted in a slight decrease in the fresh weight of both hairy root lines, by 13 % for CC16 and 15 % for CH2. After 60 min UV exposure, a decline in fresh weight (by 14%) was noted only for the CH2 line (
Figure S7). The effect of UV radiation on the hairy root dry weight was also not very prominent. After 30 min exposure it decreased by 13% and 8% in the CC16 and CH2 hairy root lines, respectively. After 60 min UV exposure, again a decline in dry weight (by 15%) was noted only for the CH2 line (
Figure S8). Neither the root color nor their morphology and branching were significantly changed.
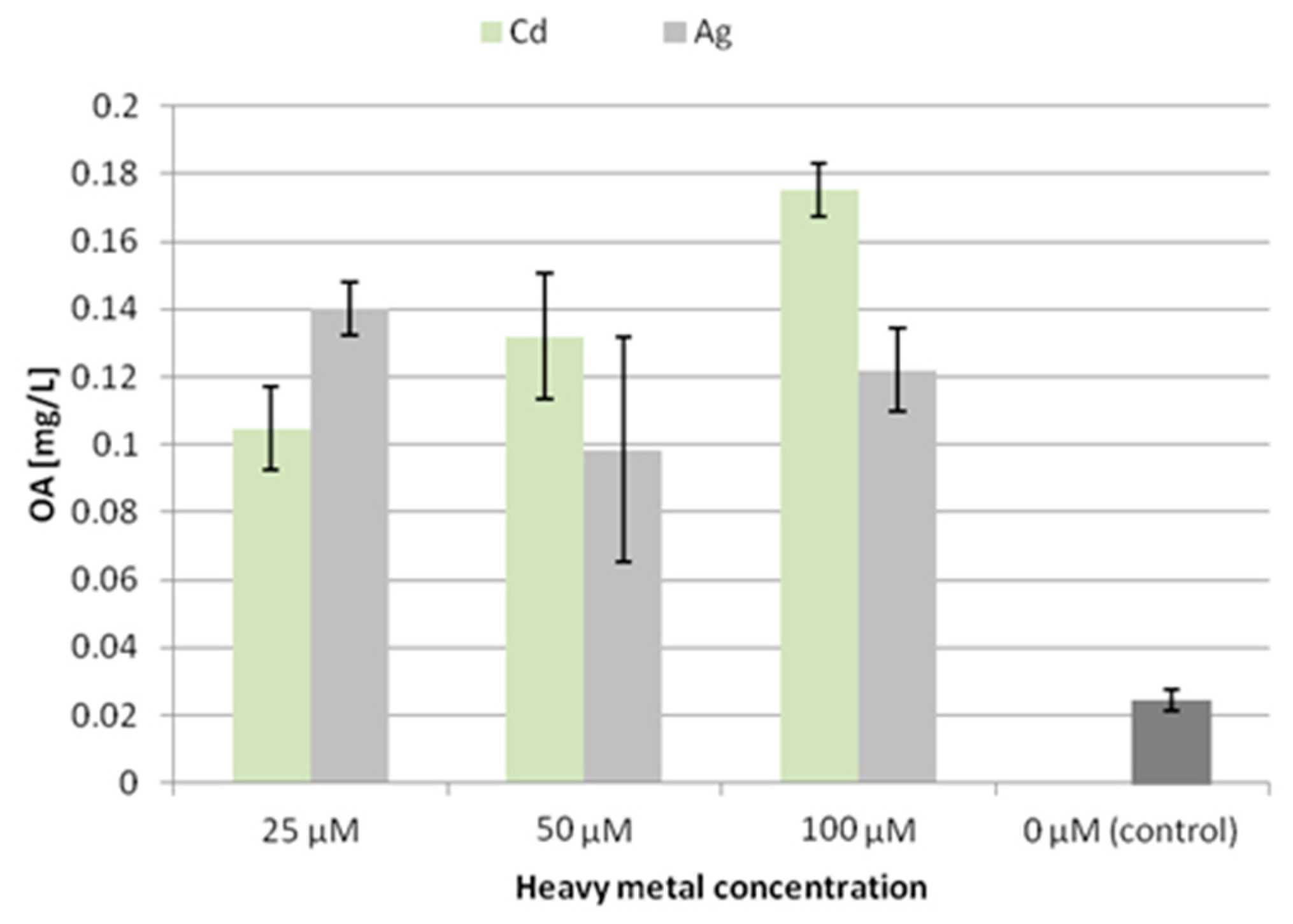
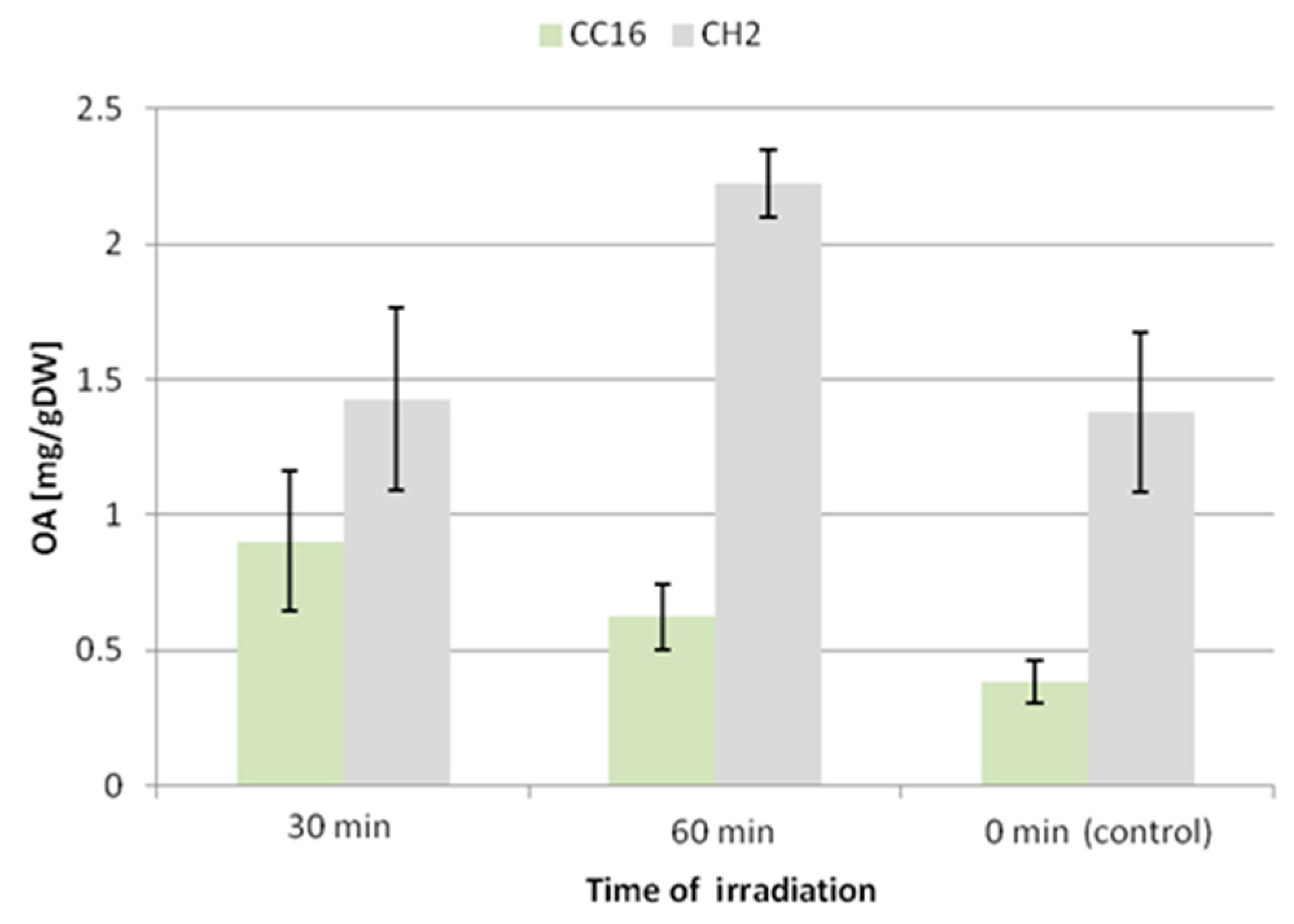
The accumulation of oleanolic acid glycosides was elevated in both hairy root lines; however, the CC16 hairy roots seemed to be more susceptible to this elicitation. After 30 and 60 min of UV exposure the accumulation of saponins in this line increased 2.4-fold and 1.6-fold, respectively. In turn, the content of saponins in CH2 hairy roots practically did not change in samples subjected to 30 min UV exposure (increased by 3%, statistically not significant), and it increased by 38% after 60 min exposure (
Figure 8).
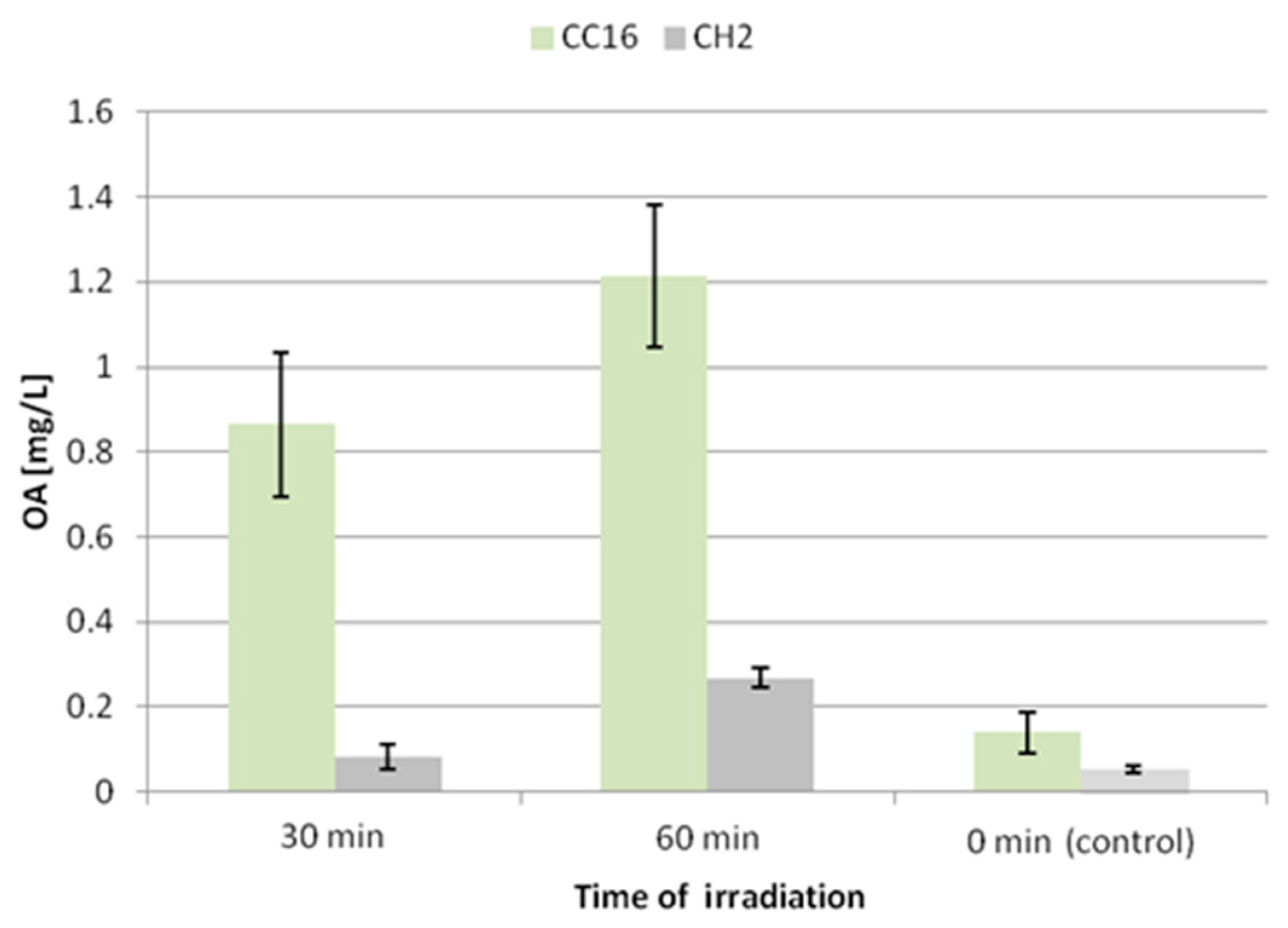
A similar effect was noted for the secretion of oleanolic acid glycosides (
Figure 9). The concentration of oleanolic acid in the medium for the CC16 line increased 6-fold and 8.5-fold after 30 and 60 min of UV exposure, respectively. This enhancement of the secretion of saponins was less significant (1.5-fold and 5-fold after 30 and 60 min of UV exposure, respectively) in the CH2 line.
In both hairy root lines the content of sterols increased, in samples exposed to 30 min UV radiation by 6% and 13% in the CC16 and CH2 lines, respectively, and by 15% and 27% in samples after 60 min of exposure (
Table 7 and
Table 8). However, the increase noted for the CC16 line was not statistically significant. Again, as after ultrasound treatment, no remarkable changes in the ratio among individual compounds were noted.
Application of UV light has often been reported to influence the production of compounds protective against this potentially harmful irradiation, such as anthocyanins [
25]. However, both UV-B (280–315 nm) and UV-C (below 280 nm) were reported to exert stimulatory effects on the production of other metabolites, e.g., UV-B stimulated the synthesis of indole alkaloids in
Catharanthus roseus hairy root cultures, while UV-C promoted the synthesis of resveratrol and other stilbenes in callus and cell suspension cultures of various
Vitis vinifera genotypes [
9]. Nowadays, there is also an increasing interest in applying UV-B or UV-C radiation to enhance the concentration of health-promoting secondary plant metabolites in plant-based foods, especially during postharvest and storage. Such treatment can lead to enhanced accumulation of flavonoids or carotenoids [
26]; however, according to the results obtained in our study, its influence on the triterpenoid content of plant material also cannot be ruled out.
Thus, the two C. officinalis hairy root lines displayed different susceptibilities to UV-C radiation. In the CC16 line, the content of oleanolic acid saponins accumulated in hairy root tissue was enhanced 2.4-fold and 1.6-fold after 30 and 60 min of UV exposure, respectively, whereas the saponin content increased by only 38% after 60 min exposure in the CH2 line. Saponin secretion increased 6-fold and 8.5-fold after 30 and 60 min of UV treatment in the CC16 line, and 1.5-fold and 5-fold in the CH2 line. In turn, the content of sterols increased more significantly in the CH2 line (by 13% and 27% after 30 and 60 min of treatment, respectively). As in the case of ultrasound, again no changes in the ratio among individual sterols were observed.














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}