
Structural, Thermal, and Storage Stability of Rapana Thomasiana Hemocyanin in the Presence of Cholinium-Amino Acid-Based Ionic Liquids

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Antibacterial and Antifouling Activity of the Cholinium Amino Acids
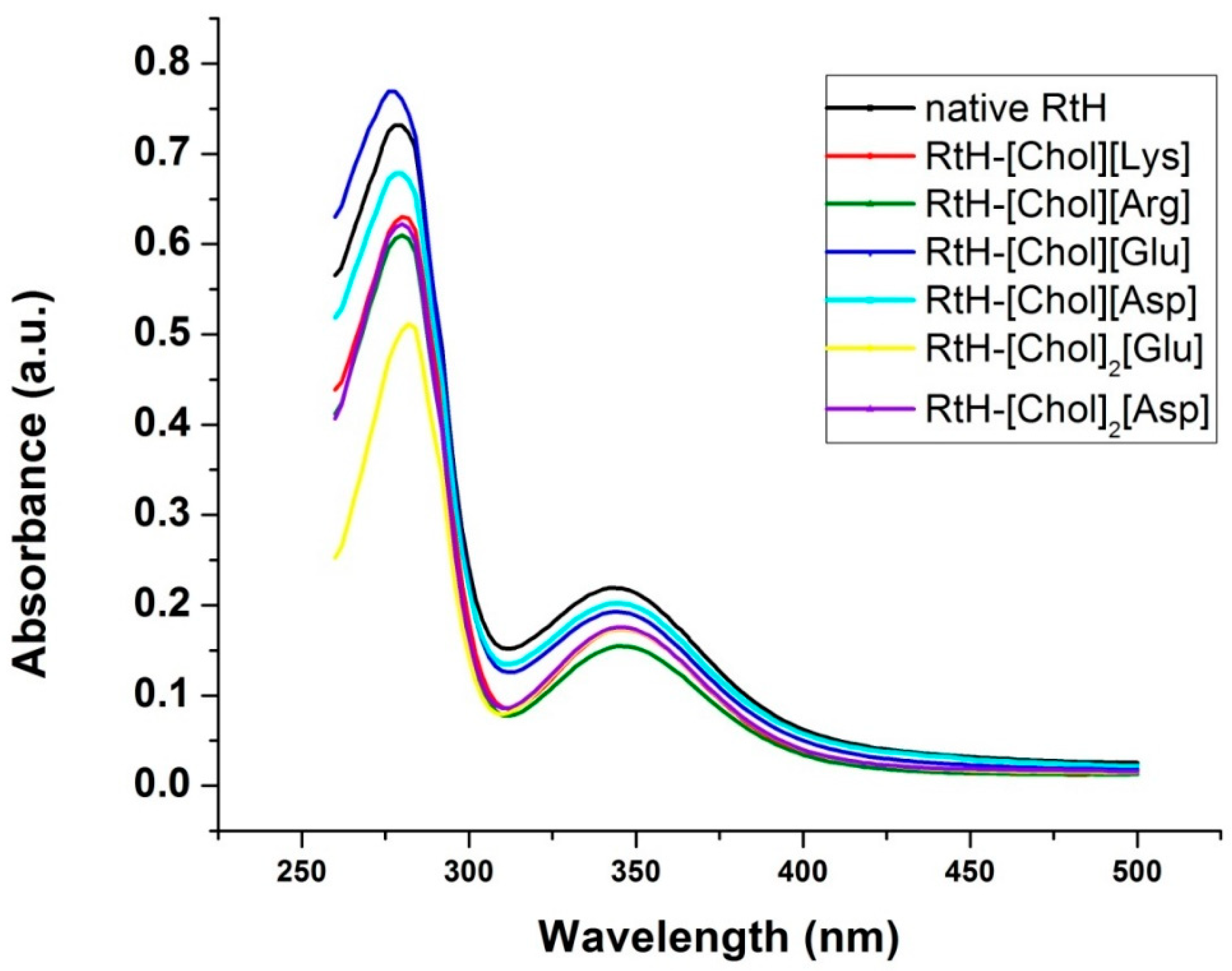
2.2. Effect of the Cholinium Amino Acids on the Conformation of Rapana Thomasiana Hemocyanin
2.3. Effect of Cholinium Amino Acids on the Thermal Stability of Rapana Thomasiana Hemocyanin
3. Materials and Methods
3.1. Materials
3.2. Antibacterial and Antibiofilm Activity of the Cholinium Amino Acids
3.3. UV-Vis Spectroscopy
3.4. Fourier Transformed Infrared Spectroscopy (FTIR)
3.5. Differential Scanning Calorimetry (DSC)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Welton, T. Ionic liquids: A brief history. Biophys. Rev. 2018, 10, 691–706. [Google Scholar] [CrossRef] [Green Version]
- Jessop, P.G.; Jessop, D.A.; Fu, D.; Phan, L. Solvatochromic parameters for solvents of interest in green chemistry. Green Chem. 2012, 14, 1245–1259. [Google Scholar] [CrossRef]
- Pollet, P.; Davey, E.A.; Ureña-Benavides, E.E.; Eckerta, C.A.; Liotta, C.L. Solvents for sustainable chemical processes. Green Chem. 2014, 16, 1034–1055. [Google Scholar] [CrossRef]
- Guglielmero, L.; Mezzetta, A.; Pomelli, C.S.; Chiappe, C.; Guazzelli, L. Evaluation of the effect of the dicationic ionic liquid structure on the cycloaddition of CO2 to epoxides. J. CO2 Util. 2019, 34, 437–445. [Google Scholar] [CrossRef]
- Keaveney, S.; Haines, R.; Harper, J. Ionic liquid solvents: The importance of microscopic interactions in predicting organic reaction outcomes. Pure Appl. Chem. 2017, 89, 745–757. [Google Scholar] [CrossRef] [Green Version]
- Shukla, S.K.; Khokarale, S.; Bui, T.; Mikkola, J.-P. Ionic liquids: Potential materials for carbon dioxide capture and utilization. Front. Mater. 2019. [Google Scholar] [CrossRef] [Green Version]
- Jonsson, E. Ionic liquids as electrolytes for energy storage applications—A modelling perspective. Energy Storage Mater. 2020, 827–835. [Google Scholar] [CrossRef]
- Tampucci, S.; Guazzelli, L.; Burgalassi, S.; Carpi, S.; Chetoni, P.; Mezzetta, A.; Nieri, P.; Polini, B.; Pomelli, C.S.; Terreni, E.; et al. pH-responsive nanostructures based on surface active fatty acid-protic ionic liquids for imiquimod delivery in skin cancer topical therapy. Pharmaceutics 2020, 12, 1078. [Google Scholar] [CrossRef]
- Santos, M.M.; Branco, L.C. Ionic liquids and deep eutectic solvents for application in pharmaceutics. Pharmaceutics 2020, 12, 909. [Google Scholar] [CrossRef]
- Hough, W.L.; Smiglak, M.; Rodrıguez, H.; Swatloski, R.P.; Spear, S.K.; Daly, D.T.; Pernak, J.; Grisel, J.E.; Carliss, R.D.; Soutullo, M.D.; et al. The third evolution of ionic liquids: Active pharmaceutical ingredients. New J. Chem. 2007, 31, 1429–1436. [Google Scholar] [CrossRef]
- Belchior, D.; Quental, M.; Pereira, M.; Mendonça, C.; Duarte, I.; Freire, M. Performance of tetraalkylammonium-based ionic liquids as constituents of aqueous biphasic systems in the extraction of ovalbumin and lysozyme. Sep. Purif. Technol. 2020, 233, 116019. [Google Scholar] [CrossRef]
- Vicente, F.; Bairos, J.; Roque, M.; Coutinho, J.; Ventura, S.; Freire, M. Use of ionic liquids as cosurfactants in mixed aqueous micellar two-phase systems to improve the simultaneous separation of immunoglobulin G and human serum albumin from expired human plasma. ACS Sustain. Chem. Eng. 2019, 7, 15102–15113. [Google Scholar] [CrossRef]
- Lee, S.Y.; Khoiroh, I.; Ooi, C.W.; Ling, T.C.; Show, P.L. Recent advances in protein extraction using ionic liquid-based aqueous two-phase systems. Sep. Purif. Rev. 2017, 46, 291–304. [Google Scholar] [CrossRef]
- Schindl, A.; Hagen, M.; Muzammal, S.; Gunasekera, H.; Croft, A. Proteins in ionic liquids: Reactions, applications, and futures. Front. Chem. 2019, 7, 347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbari, N.; Daneshjoo, S.; Akbari, J.; Khajeh, K. Isolation, characterization, and catalytic properties of a novel lipase which is activated in ionic liquids and organic solvents. Appl. Biochem. Biotechnol. 2011, 165, 785–794. [Google Scholar] [CrossRef]
- Ha, S.H.; Anh, T.V.; Lee, S.H.; Koo, Y.-M. Effect of ionic liquids on enzymatic synthesis of caffeic acid phenethyl ester. Bioprocess. Biosyst. Eng. 2012, 35, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Mei, L. Lyophilization of lipase with cyclodextrins for efficient catalysis in ionic liquids. J. Biosci. Bioeng. 2007, 103, 345–349. [Google Scholar] [CrossRef]
- Guncheva, M.; Dimitrov, M.; Napoly, F.; Draye, M.; Andrioletti, B. Novel hybrid materials on the basis of nanostructured tin dioxide and a lipase from Rhizopus delemar with improved enantioselectivity. J. Mol. Cat. B: Enzym. 2014, 102, 72–80. [Google Scholar] [CrossRef]
- Fujita, K. Ionic liquids as stabilization and refolding additives and solvents for proteins. Adv. Biochem. Eng. Biotechnol. 2019, 168, 215–226. [Google Scholar] [CrossRef]
- Todinova, S.; Guncheva, M.; Yancheva, D. Thermal and conformational stability of insulin in the presence of imidazolium-based ionic liquids. J. Therm. Anal. Calorim. 2016, 123, 2591–2598. [Google Scholar] [CrossRef]
- Guncheva, M.; Ossowicz, P.; Janus, E.; Todinova, S.; Yancheva, D. Elucidation of the effect of some cholinium amino acid ionic liquids on the thermal and the conformational stability of insulin. J. Mol. Liq. 2019, 283, 257–262. [Google Scholar] [CrossRef]
- van Holde, K.E.; Miller, K.I. Hemocyanins. Adv. Protein. Chem. 1995, 47, 1–81. [Google Scholar] [CrossRef]
- Croates, C.J.; Narin, J. Diverse immune functions of hemocyanins. Dev. Comp. Immunol. 2014, 45, 43–55. [Google Scholar] [CrossRef]
- Kato, S.; Matsui, T.; Gatsogiannis, C.; Tanaka, Y. Molluscan hemocyanin: Structure, evolution, and physiology. Biophys. Rev. 2018, 10, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Salama, W.; Mona, M. In vitro anti-tumor effects of hemocyanin isolated from Atergatis roseus and Eriphia verrucosa crabs. J. Cancer Biomed. Res. (JCBR) 2018, 1, 28–36. [Google Scholar] [CrossRef] [Green Version]
- McFadden, D.; Riggs, D.; Jackson, B.; Vona-Davis, L. Keyhole limpet hemocyanin, a novel immune stimulant with promising anticancer activity in Barrett’s esophageal adenocarcinoma. Am. J. Surg. 2003, 186, 552–555. [Google Scholar] [CrossRef] [PubMed]
- Gesheva, V.; Chausheva, S.; Mihaylova, N.; Manoylov, I.; Doumanova, L.; Idakieva, K.; Tchorbanov, A. Anti-cancer properties of gastropodan hemocyanins in murine model of coloncarcinoma. BMC Immunol. 2014, 15, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonova, O.; Dolashka, P.; Toncheva, D.; Rammensee, H.-G.; Floetenmeyer, M.; Stevanovic, S. In vitro antiproliferative effect of Helix aspersa hemocyanin on multiple malignant cell lines. Z. Naturforsch. C. J. Biosci. 2014, 69, 325–334. [Google Scholar] [CrossRef]
- Guncheva, M.; Paunova, K.; Ossowicz, P.; Rozwadowski, Z.; Janus, E.; Idakieva, K.; Todinova, S.; Raynova, Y.; Uzunova, V.; Apostolova, S.; et al. Modification of Rapana thomasiana hemocyanin with choline amino acid salts significantly enhances its antiproliferative activity against MCF-7 human breast cancer cells. RSC Adv. 2015, 5, 63345–63354. [Google Scholar] [CrossRef]
- Guncheva, M.; Paunova, K.; Ossowicz, P.; Rozwadowski, Z.; Janus, E.; Idakieva, K.; Todinova, S.; Raynova, Y.; Uzunova, V.; Apostolova, S.; et al. Rapana thomasiana hemocyanin modified with ionic liquids with enhanced anti breast cancer activity. Int. J. Biol. Macromol. 2016, 82, 798–805. [Google Scholar] [CrossRef] [PubMed]
- Guncheva, M.; Todinova, S.; Uzunova, V.; Idakieva, K.; Raynova, Y.; Ossowicz, P.; Janus, E.; Tzoneva, R. Destabilization of β-hemocyanin from Helix pomatia in presence of choline amino acids results in improved cell specificity and cytotoxicity against human breast cancer. ChemistrySelect 2019, 4, 11460–11466. [Google Scholar] [CrossRef]
- Hemocyanin from Megathura Crenulata (Keyhole Limpet). Available online: https://www.sigmaaldrich.com/content/dam/sigma-aldrich/docs/Sigma/Datasheet/2/h7017dat.pdf (accessed on 27 October 2020).
- Zanjani, N.T.; Sairi, F.; Marshall, G.; Saksena, M.M.; Valtchev, P.; Gomes, V.G.; Cunningham, A.L.; Dehghani, F. Formulation of abalone hemocyanin with high antiviral activity and stability. Eur. J. Pharm. Sci. 2014, 53, 77–85. [Google Scholar] [CrossRef]
- Modi, K.D.; Gadge, P.V.; Jain, P.; Pawar, S.; Shah, R.D.; Ingole, S.A.; Jain, R. Clinical challenges with excipients ininsulin formulation sand role of concentrated insulin. Int. J. Basic Clin. Pharmacol. 2019, 8, 821–826. [Google Scholar] [CrossRef]
- Wang, W. Advanced protein formulations. Prot. Sci. 2015, 24, 1031–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Challener, C.A. Excipient Selection for protein stabilization. Pharm. Technol. 2015, 3, S35–S39. [Google Scholar]
- Florio, W.; Becherini, S.; D’Andrea, F.; Lupetti, A.; Chiappe, C.; Guazzelli, L. Comparative evaluation of antimicrobial activity of different types of ionic liquids. Mater. Sci. Eng. C 2019, 104, 109907. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhang, S.; Dai, Y.; Lu, X.; Lei, Q.; Fang, W. Antimicrobial activity and cytotoxicity of piperazinium- and guanidinium-based ionic liquids. J. Hazard. Mater. 2016, 307, 73–81. [Google Scholar] [CrossRef]
- Yazdani, A.; Sivapragasam, M.; Levêque, J.M.; Moniruzzaman, M. Microbial biocompatibility and biodegradability of choline-amino acid based ionic liquids. J. Microb. Biochem. Technol. 2016, 8, 415–421. [Google Scholar] [CrossRef]
- Beltramini, M.; Bubacco, L.; Casella, L.; Alzuet, G.; Gullott, M.; Salvato, B. The oxidation of hernocyanin. Kinetics, reaction mechanism and characterization of Met-hemocyanin product. Eur. J. Biochem. 1995, 232, 98–105. [Google Scholar] [CrossRef]
- Idakieva, K.; Parvanova, K.; Todinova, S. Differential scanning calorimetry of the irreversible denaturation of Rapana thomasiana (marine snail, Gastropod) hemocyanin. Biochim. Biophys. Acta 2005, 1748, 50–56. [Google Scholar] [CrossRef]
- De Soyza, A.; Hall, E.; Mahenthiralingam, E.; Drevinek, P.; Kaca, W.; Drulis-Kawa, Z.; Stoitsova, S.R.; Toth, V.; Coenye, T.; Zlosnik, J.E.A.; et al. Developing an international Pseudomonas aeruginosa reference panel. Microbiol. Open 2013, 2, 1010–1023. [Google Scholar] [CrossRef]
- Barth, A. Infrared spectroscopy of proteins. Biochim. Biophys. Acta Bioenerg. 2007, 1767, 1073–1101. [Google Scholar] [CrossRef] [Green Version]
- Guncheva, M.; Idakieva, K.; Todinova, S.; Stoyanova, E.; Yancheva, D. Folate-conjugated Helix lucorum hemocyanin–preparation, stability, and cytotoxicity. Z. Naturforsch. C 2020, 75, 23–30. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | IC 50 # (mmol/L) | |||||
|---|---|---|---|---|---|---|
| [Chol][Glu] | [Chol][Asp] | [Chol]2[Glu] | [Chol]2[Asp] | [Chol][Lys] | [Chol][Arg] | |
| Gram-negative | ||||||
| E. coli 420 | no effect | no effect | 10.27 ± 0.82 | 7.29 ± 0.20 | 3.54 ± 0.16 | 6.14 ± 0.30 |
| E. coli 25 922 | no effect | no effect | 5.73 ± 0.26 | 34.9 ± 1.75 | 2.37 ± 0.04 | 2.9 ± 0.11 |
| P. aeruginosa PAO1 | no effect | no effect | no effect | promote bacterial growth ~ 10% | 6.40 ± 0.13 | 20.12 ± 0.37 |
| Gram-positive | ||||||
| S. aureus 29 213 | 5.91 ± 0.17 | 41.8 ± 1.19 | 13.8 ± 0.36 | 25.6 ± 0.61 | 5.23 ± 0.05 | 18.75 ± 0.56 |
| S. saprophyticus 15 305 | no effect | promote bacterial growth ~ 10% | 4.36 ± 0.16 | 12.5 ± 0.28 | 1.67 ± 0.13 | 2.39 ± 0.09 |
| B. subtilis 168 | promote bacterial growth ~12% | promote bacterial growth ~ 7% | promote bacterial growth ~ 5% | 14.7 ± 0.49 | 4.37 ± 0.22 | 8.48 ± 0.51 |
| Bacterial Strain | Biofilm Formation Inhibition, % # | |||||
|---|---|---|---|---|---|---|
| [Chol][Glu] | [Chol][Asp] | [Chol]2[Glu] | [Chol]2[Asp] | [Chol][Lys] | [Chol][Arg] | |
| Gram-negative | ||||||
| E. coli 420 | NA | NA | 53.1 ± 2.7 | 34.1 ± 1.9 47.1 ±2.5 (a) | 46.6 ± 2.3 57.1 ±2.1 (b) | 57.2 ± 2.5 |
| E. coli 25 922 | 76.3 ± 3.8 | 86.5 ± 2.5 | 91.2 ± 3.4 | 90.2 ± 4.1 | 89.2 ± 3.6 | 89.9 ± 3.4 |
| P. aeruginosa PAO1 | 43.5 ± 2.2 | 38.2 ± 3.1 | 49.5 ± 2.8 | 26.8 ± 1.8 62.6± 2.8 (b) | no effect 32.8± 2.0 (b) | 7.8 ± 0.5 60.8± 3.1 (b) |
| Gram-positive | ||||||
| S. aureus 29 213 | 40.1 ± 4.3 | 3.6 ± 2.1 29.4 ± 1.6 (b) | 23.9 ± 1.7 | 5.3 ± 0.9 56.6 ± 3.9 (b) | 7.8 ± 0.7 60.9 ± 2.5 | |
| S. saprophyticus 15 305 | 27.0 ± 3.1 36.8 ± 2.8 (a) | 24.8 ± 1.5 66.7 ± 2.6 (a) | 48.2 ± 1.8 87.2 ± 3.6 (a) | 40.2 ± 2.9 51.7 ± 2.9 (b) | 64.5 ± 2.1 94.5 ± 3.4 (b) | 60.7 ± 2.9 95.4 ± 3.1 (b) |
| B. subtilis 168 | no effect 38.7 ± 2.0 (b) | 44.3 ± 1.9 | 48.5 ± 1.7 | 54.9 ± 2.5 | 42.5 ± 2.8 | no effect 35.11 ± 1.7 (b) |
| Parameters Sample | Tm1 (°C) | ∆H1 (kJ mol−1) | Tm2 (°C) | ∆H2 (kJ mol−1) | Tm3 (°C) | ∆H3 (kJ mol−1) | Tm4 (°C) | ∆H4 (kJ mol−1) | ∆Htotal (kJ mol−1) |
|---|---|---|---|---|---|---|---|---|---|
| native RtH | 60.0 | 63,901 | 70.2 | 59,132 | 75.8 | 47,597 | 83.3 | 38696 | 194,637 |
| RtH-[Chol][Glu] | 62.0 | 74,040 | 70.4 | 100,553 | 77.7 | 46,898 | 85.0 | 23,272 | 244,962 |
| RtH-[Chol][Asp] | 71.4 | 61,384 | 77.3 | 45114 | 86.4 | 24,055 | 128,482 | ||
| RtH-[Chol]2[Glu] | 55.6 | 28,715 | 64.3 | 46,586 | 71.1 | 72,300 | 80.6 | 35,821 | 183,342 |
| RtH-[Chol]2[Asp] | 69.0 | 62,104 | 75.3 | 21,030 | 82.5 | 24,223 | 95.1 | 15,435 | 127,910 |
| RtH-[Chol][Arg] | 64.1 | 29,106 | 69.6 | 34,310 | 75.6 | 64,704 | 85.8 | 17310 | 136,582 |
| RtH-[Chol][Lys] | 62.3 | 13,980 | 68.5 | 16,213 | 73.6 | 13,483 | 78.2 | 12,113 | 56,067 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guncheva, M.; Idakieva, K.; Todinova, S.; Yancheva, D.; Paunova-Krasteva, T.; Ossowicz, P.; Janus, E. Structural, Thermal, and Storage Stability of Rapana Thomasiana Hemocyanin in the Presence of Cholinium-Amino Acid-Based Ionic Liquids. Molecules 2021, 26, 1714. https://doi.org/10.3390/molecules26061714
Guncheva M, Idakieva K, Todinova S, Yancheva D, Paunova-Krasteva T, Ossowicz P, Janus E. Structural, Thermal, and Storage Stability of Rapana Thomasiana Hemocyanin in the Presence of Cholinium-Amino Acid-Based Ionic Liquids. Molecules. 2021; 26(6):1714. https://doi.org/10.3390/molecules26061714
Chicago/Turabian StyleGuncheva, Maya, Krassimira Idakieva, Svetla Todinova, Denitsa Yancheva, Tsvetelina Paunova-Krasteva, Paula Ossowicz, and Ewa Janus. 2021. "Structural, Thermal, and Storage Stability of Rapana Thomasiana Hemocyanin in the Presence of Cholinium-Amino Acid-Based Ionic Liquids" Molecules 26, no. 6: 1714. https://doi.org/10.3390/molecules26061714