
Targeted and Untargeted Metabolomic Analyses Reveal Organ Specificity of Specialized Metabolites in the Model Grass Brachypodium distachyon

 , ,
, ,
Abstract
:
1. Introduction
2. Results and Discussion
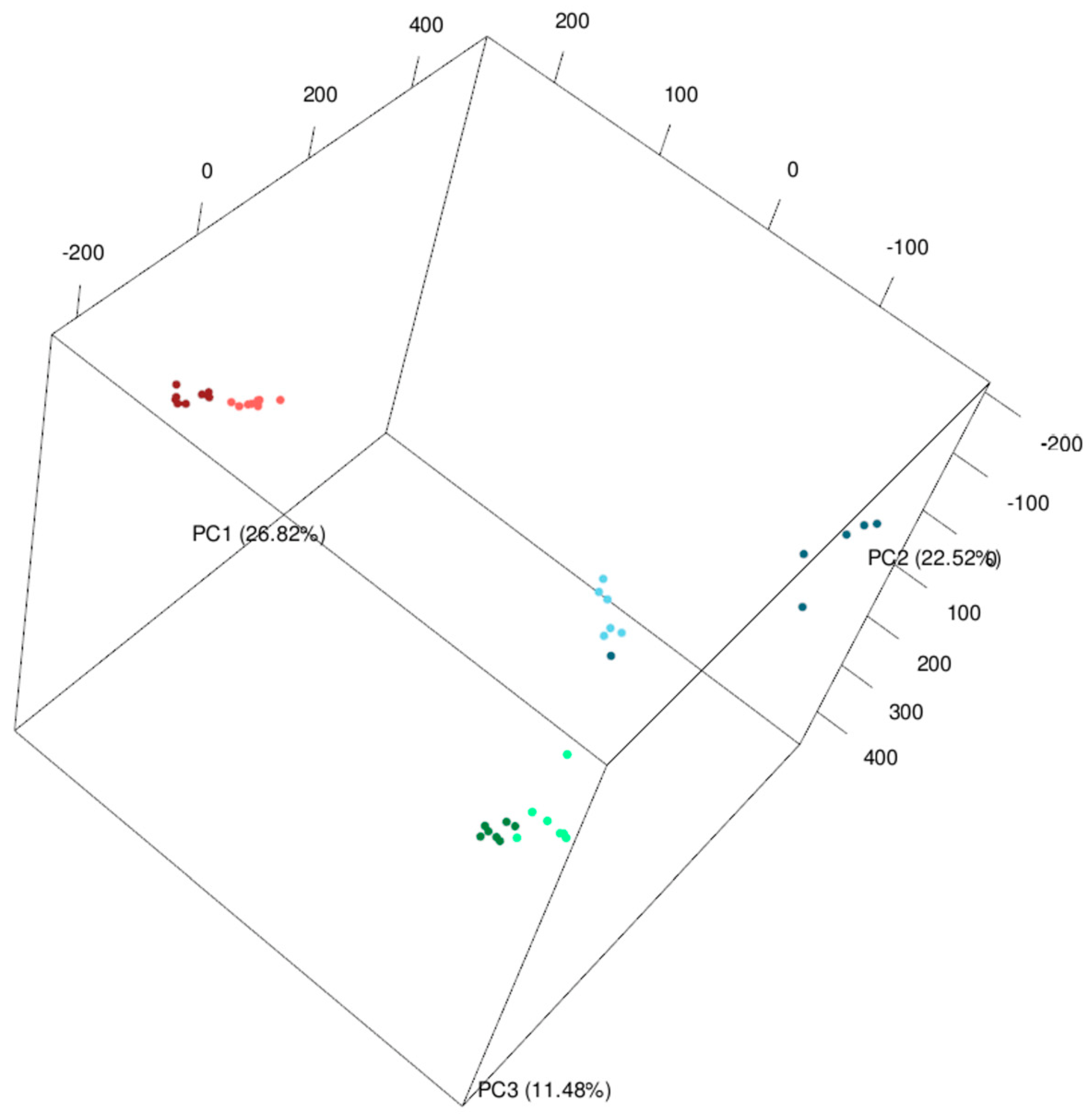
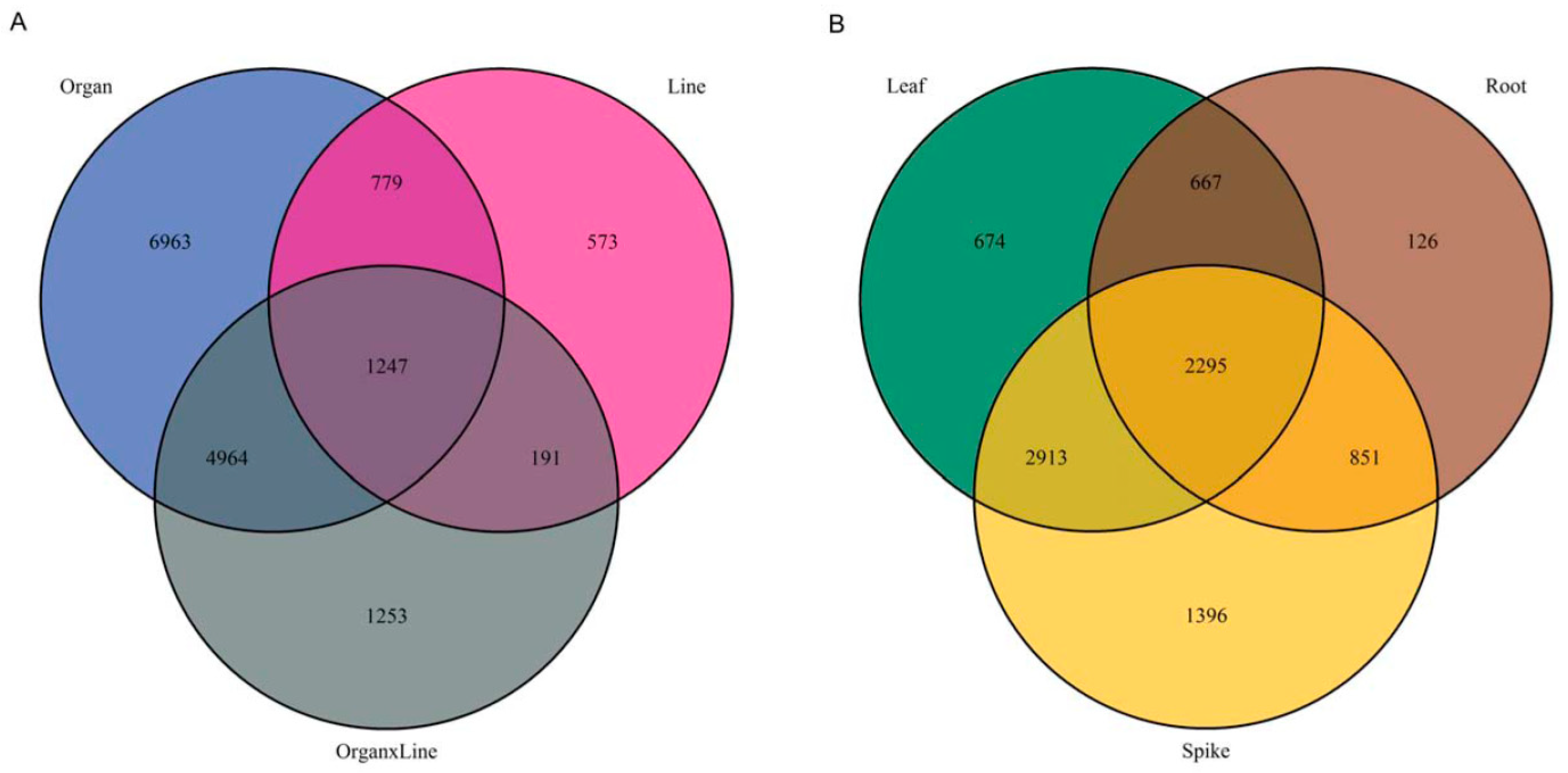
2.1. Comparison of Metabolomics Profiles in Analyzed Brachypodium Organs and Lines
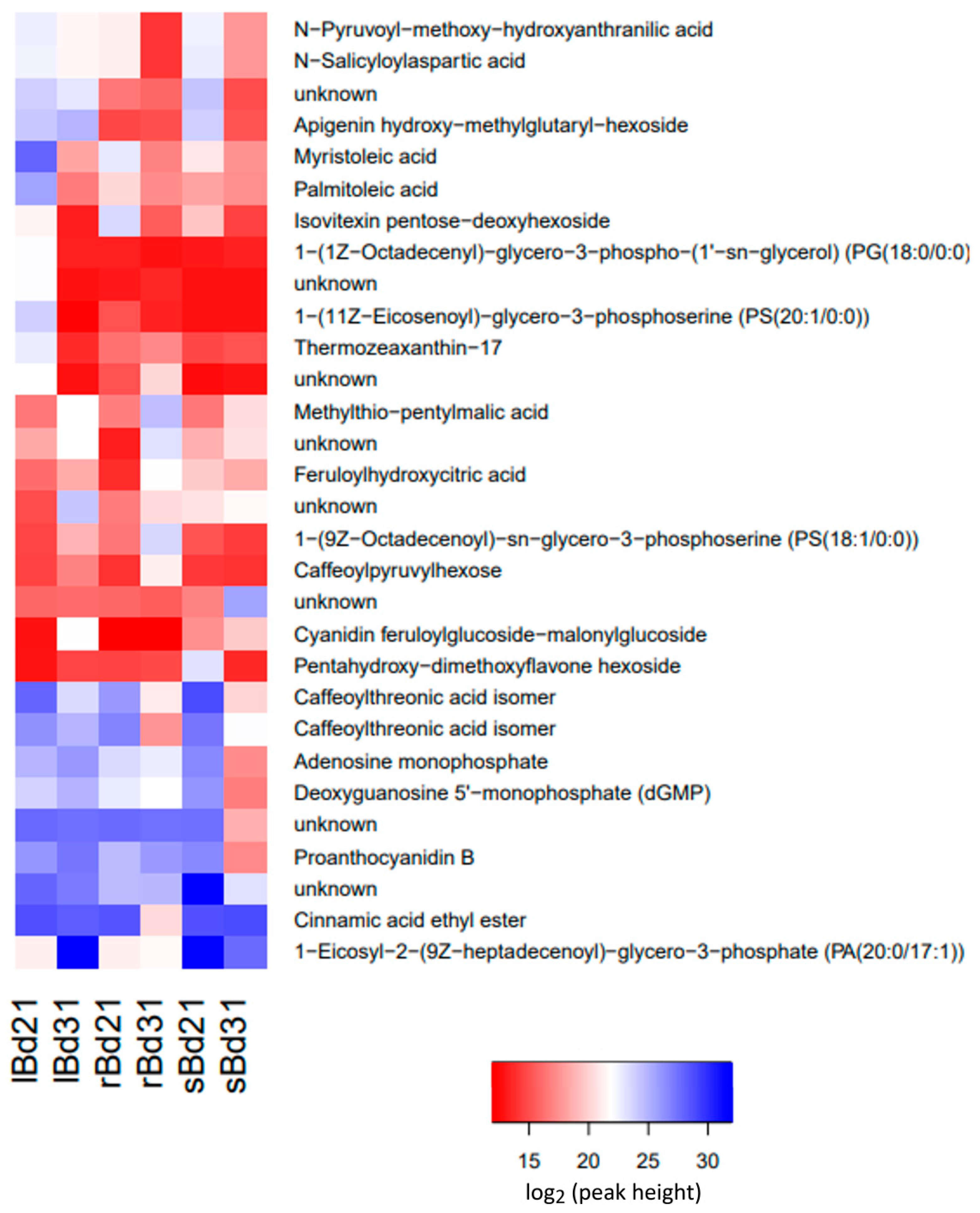
2.2. Pathway Enrichment Analysis
2.2.1. Most Represented Metabolic Pathways
2.2.2. Metabolic Pathways Distinguishing Bd21 and Bd3-1 Lines
2.3. Metabolite Identification with LC-MS Systems
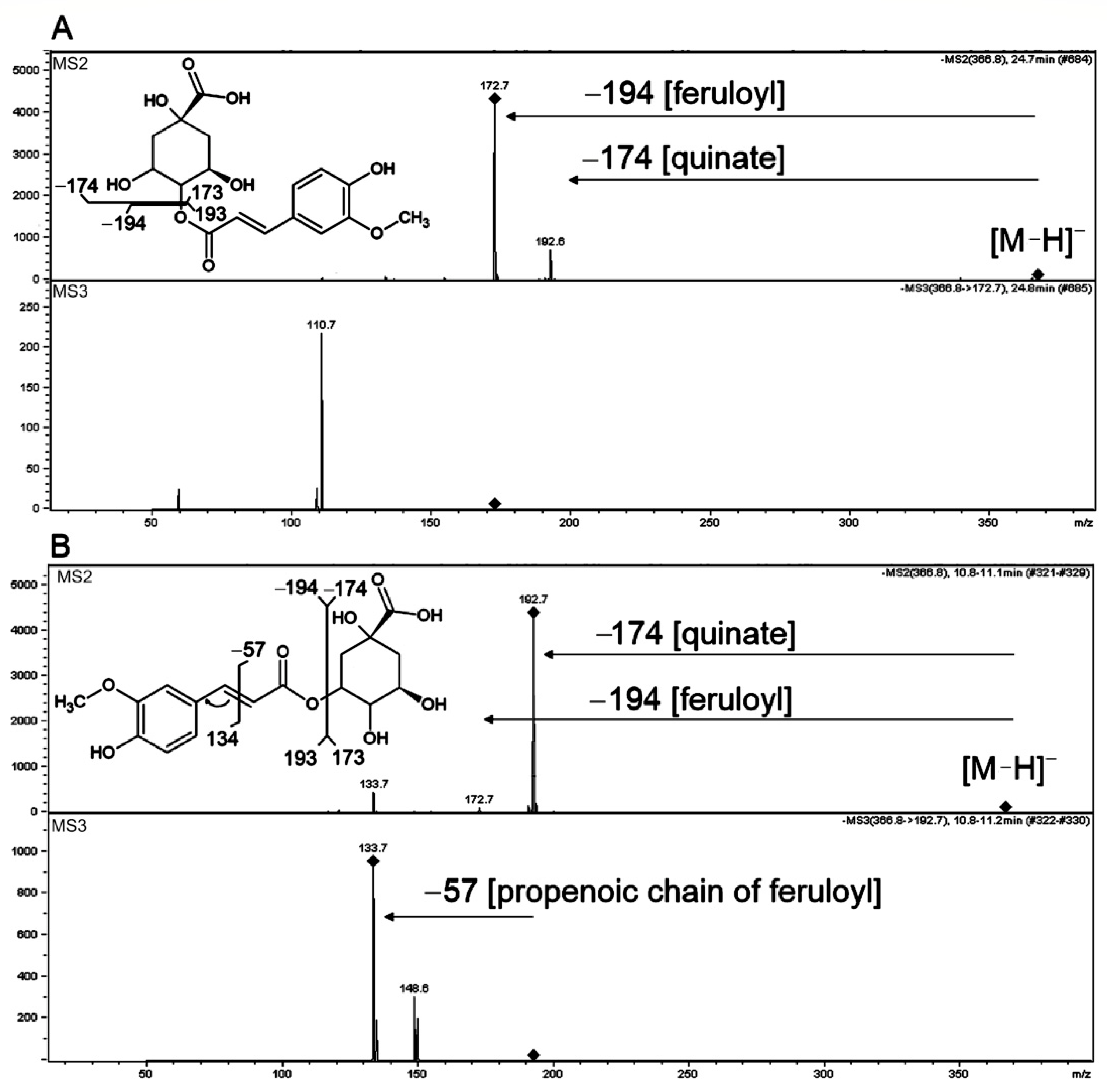
2.3.1. Hydroxycinnamoyl-Quinic Acids
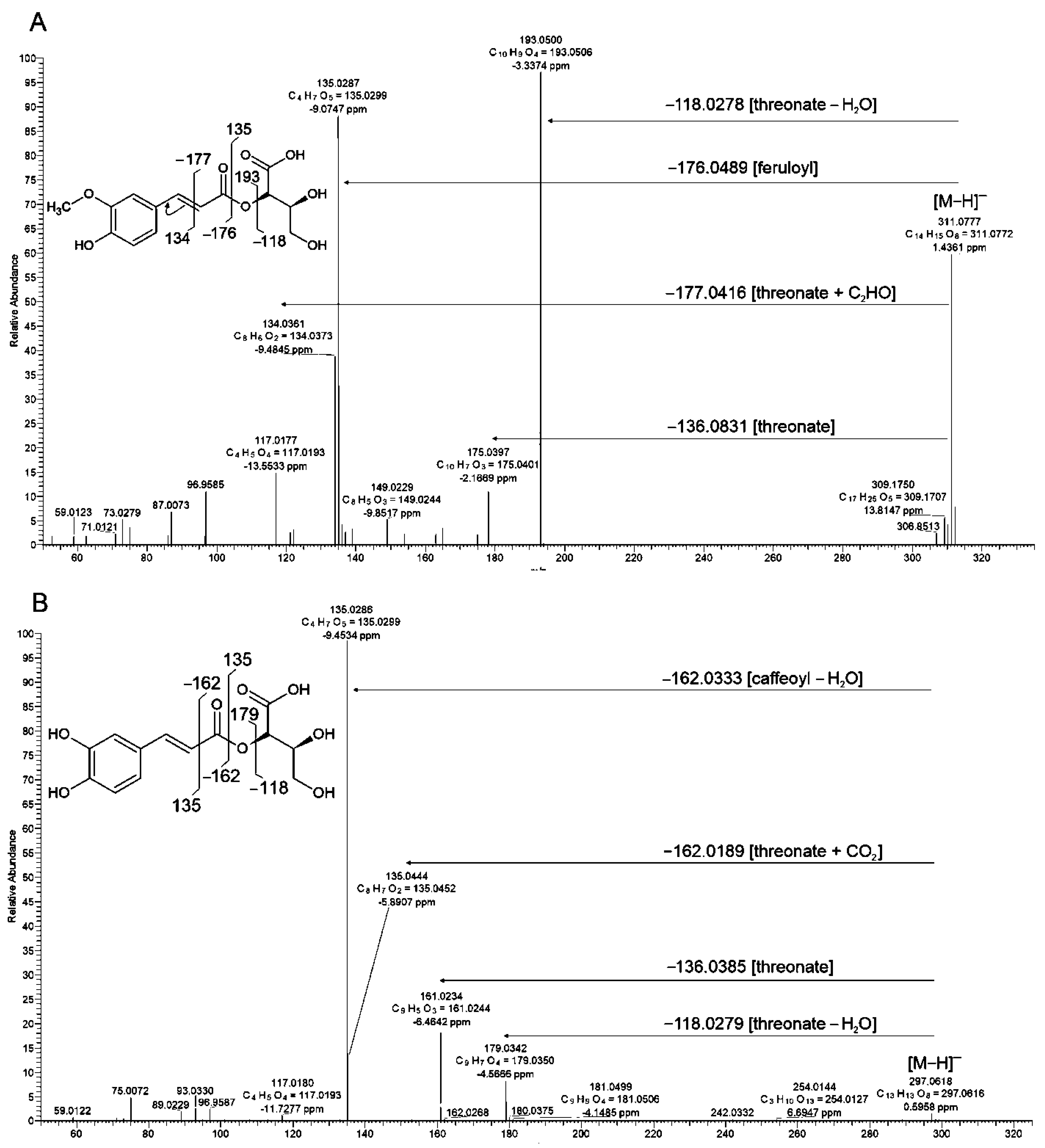
2.3.2. Esters of Hydroxycinnamic and Threonic Acids
2.3.3. Hydroxycinnamic Acid Amides
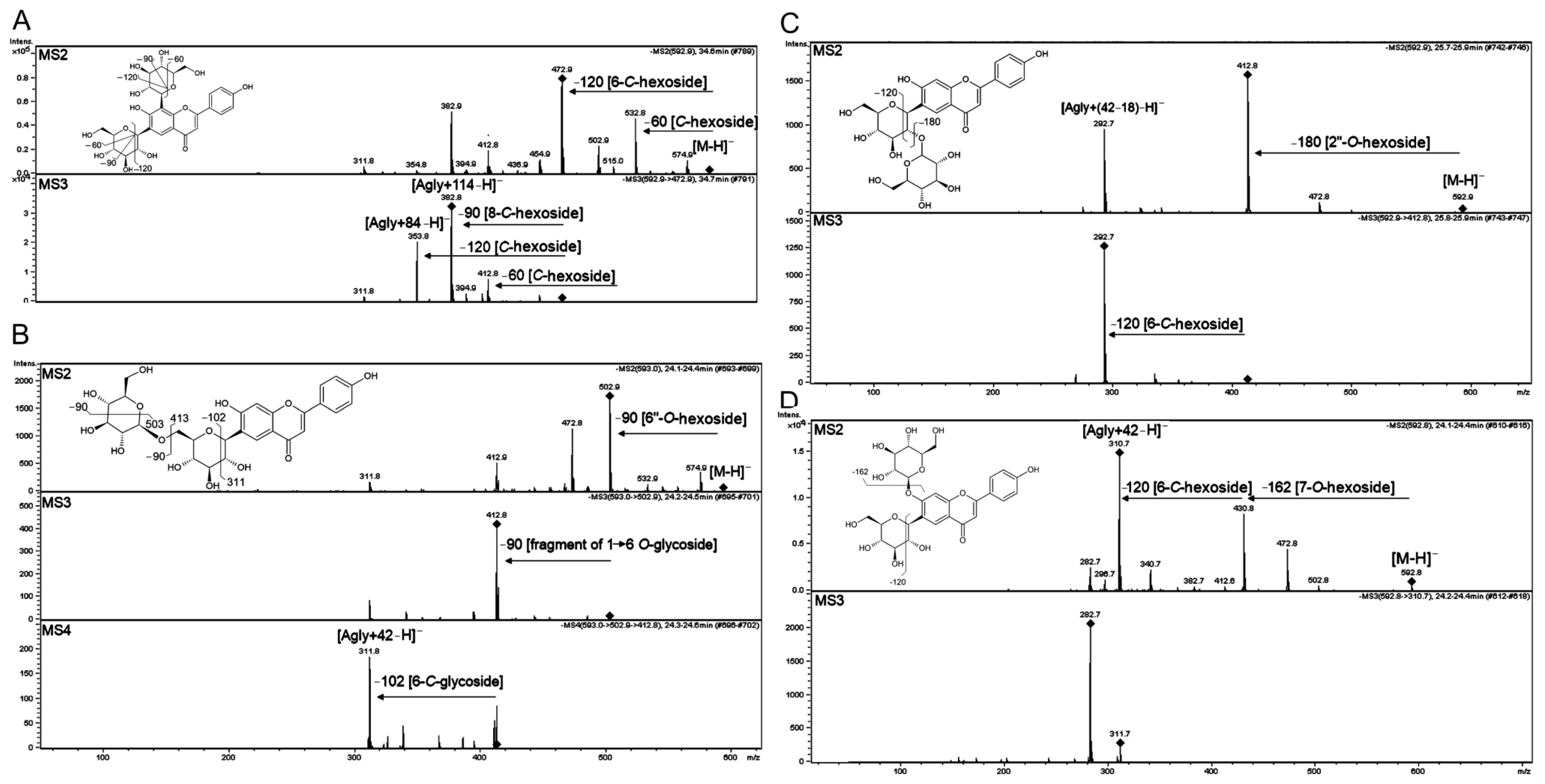
2.3.4. Flavonoid Glycosides
O-Glycosides
O,C-Glycosides
Di-C-Glycosides
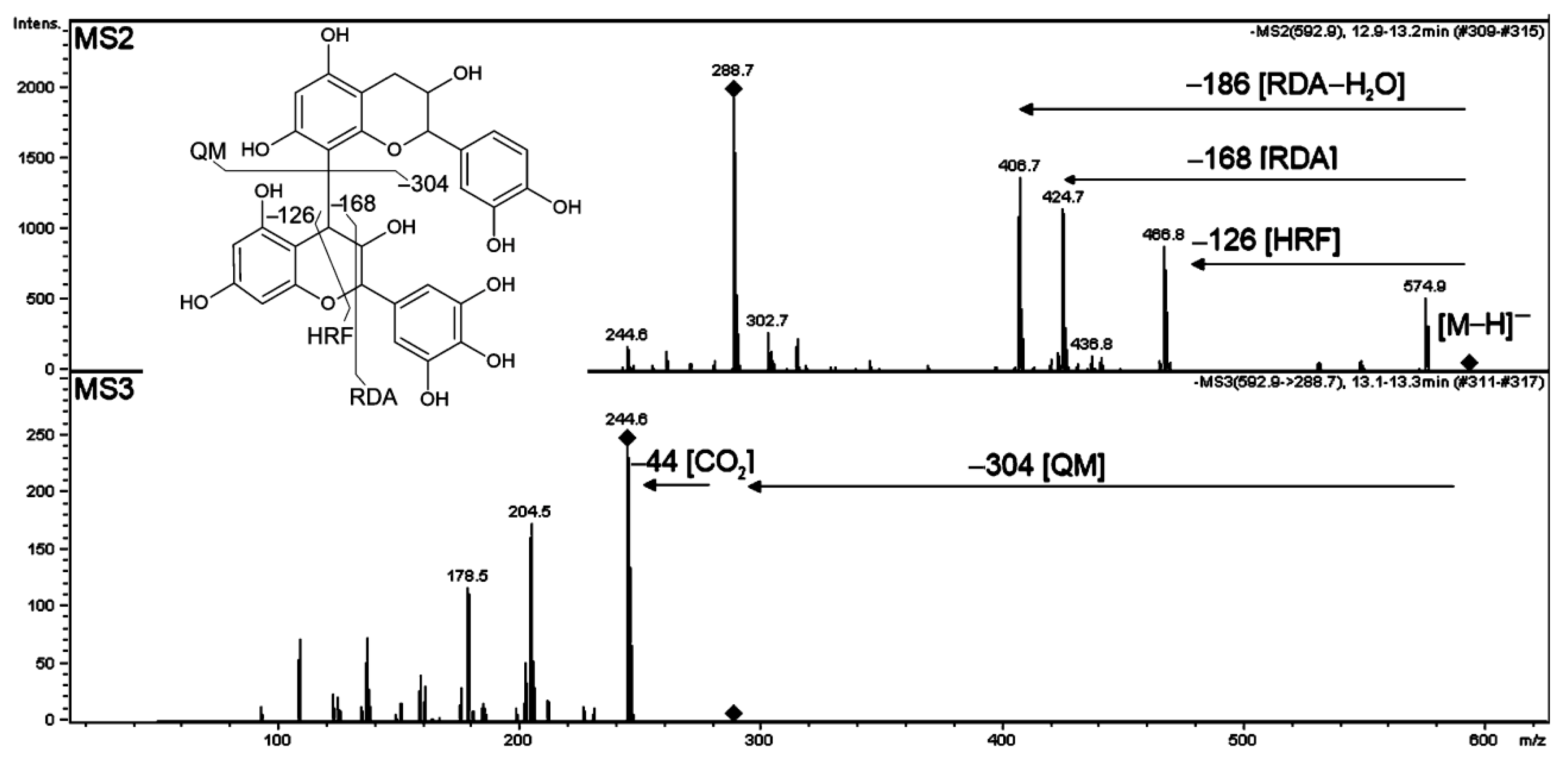
2.3.5. Acylated Flavonoids
2.3.6. Flavan-3-ols
3. Material and Methods
3.1. Plant Material
3.2. Chemicals
3.3. Metabolite Profiling
3.4. Metabolite Identification
3.5. Bioinformatic Processing
3.6. Statistical Analysis
3.7. Pathway Enrichment Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Huo, N.; Lazo, G.R.; Vogel, J.P.; You, F.M.; Ma, Y.; Hayden, D.M.; Coleman-Derr, D.; Hill, T.A.; Dvorak, J.; Anderson, O.D.; et al. The nuclear genome of Brachypodium distachyon: Analysis of BAC end sequences. Funct. Integr. Genom. 2008, 8, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Brkljacic, J.; Grotewold, E.; Scholl, R.; Mockler, T.; Garvin, D.F.; Vain, P.; Brutnell, T.; Sibout, R.; Bevan, M.; Budak, H.; et al. Brachypodium as a Model for the Grasses: Today and the Future. Plant Physiol. 2011, 157, 3–13. [Google Scholar] [CrossRef]
- The International Brachypodium Initiative. Genome sequencing and analysis of the model grass Brachypodium distachyon. Nature 2010, 463, 763–768. [Google Scholar] [CrossRef]
- Goodstein, D.M.; Shu, S.Q.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef] [PubMed]
- Tello-Ruiz, M.K.; Naithani, S.; Gupta, P.; Olson, A.; Wei, S.; Preece, J.; Jiao, Y.P.; Wang, B.; Chougule, K.; Garg, P.; et al. Gramene 2021: Harnessing the power of comparative genomics and pathways for plant research. Nucleic Acids Res. 2021, 49, D1452–D1463. [Google Scholar] [CrossRef]
- Gordon, S.P.; Contreras-Moreira, B.; Woods, D.P.; Marais, D.L.D.; Burgess, D.; Shu, S.Q.; Stritt, C.; Roulin, A.C.; Schackwitz, W.; Tyler, L.; et al. Extensive gene content variation in the Brachypodium distachyon pan-genome correlates with population structure. Nat. Commun. 2017, 8, 2184. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Nadege, S.W.; Huang, C.; Zhang, P.; Song, S.; Sun, L.; Wu, Y. Brachypodium distachyon is a suitable host plant for study of Barley yellow dwarf virus. Virus Genes 2016, 52, 299–302. [Google Scholar] [CrossRef]
- do Amaral, F.P.; Pankievicz, V.C.S.; Arisi, A.C.M.; de Souza, E.M.; Pedrosa, F.; Stacey, G. Differential growth responses of Brachypodium distachyon genotypes to inoculation with plant growth promoting rhizobacteria. Plant Mol. Biol. 2016, 90, 689–697. [Google Scholar] [CrossRef]
- Powell, J.J.; Carere, J.; Sablok, G.; Fitzgerald, T.L.; Stiller, J.; Colgrave, M.L.; Gardiner, D.M.; Manners, J.M.; Vogel, J.P.; Henry, R.J.; et al. Transcriptome analysis of Brachypodium during fungal pathogen infection reveals both shared and distinct defense responses with wheat. Sci. Rep. 2017, 7, 17212. [Google Scholar] [CrossRef]
- Kong, L.A.; Wu, D.Q.; Cui, J.K.; Huang, W.K.; Peng, H.; Peng, D.L. Testing and modelling the potential of three diploid plants in Poaceae as a new pathosystem to investigate the interactions between cereal hosts and cereal cyst nematode (Heterodera avenae). Plant Pathol. 2016, 65, 682–688. [Google Scholar] [CrossRef]
- Priest, H.D.; Fox, S.E.; Rowley, E.R.; Murray, J.R.; Michael, T.P.; Mockler, T.C. Analysis of Global Gene Expression in Brachypodium distachyon Reveals Extensive Network Plasticity in Response to Abiotic Stress. PLoS ONE 2014, 9, e87499. [Google Scholar] [CrossRef]
- Onda, Y.; Hashimoto, K.; Yoshida, T.; Sakurai, T.; Sawada, Y.; Hirai, M.Y.; Toyooka, K.; Mochida, K.; Shinozaki, K. Determination of growth stages and metabolic profiles in Brachypodium distachyon for comparison of developmental context with Triticeae crops. Proc. R. Soc. B 2015, 282, 20150964. [Google Scholar] [CrossRef]
- Onda, Y.; Inoue, K.; Sawada, Y.; Shimizu, M.; Takahagi, K.; Uehara-Yamaguchi, Y.; Hirai, M.Y.; Garvin, D.F.; Mochida, K. Genetic Variation for Seed Metabolite Levels in Brachypodium distachyon. Int. J. Mol. Sci. 2019, 20, 2348. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Alvarez, D.; Zubair, H.; Beckmann, M.; Draper, J.; Catalan, P. Diversity and association of phenotypic and metabolomic traits in the close model grasses Brachypodium distachyon, B. stacei and B. hybridum. Ann. Bot. 2017, 119, 545–561. [Google Scholar] [PubMed]
- Pasquet, J.C.; Chaouch, S.; Macadre, C.; Balzergue, S.; Huguet, S.; Martin-Magniette, M.L.; Bellvert, F.; Deguercy, X.; Thareau, V.; Heintz, D.; et al. Differential gene expression and metabolomic analyses of Brachypodium distachyon infected by deoxynivalenol producing and non-producing strains of Fusarium graminearum. BMC Genom. 2014, 15, 629. [Google Scholar] [CrossRef] [PubMed]
- Fisher, L.H.C.; Han, J.W.; Corke, F.M.K.; Akinyemi, A.; Didion, T.; Nielsen, K.K.; Doonan, J.H.; Mur, L.A.J.; Bosch, M. Linking Dynamic Phenotyping with Metabolite Analysis to Study Natural Variation in Drought Responses of Brachypodium distachyon. Front. Plant Sci. 2016, 7, 1751. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Ye, T.; Song, B.; Qi, X.; Chan, Z. Comparative physiological and metabolomic responses of four Brachypodium distachyon varieties contrasting in drought stress resistance. Acta Physiol. Plant. 2015, 37, 122. [Google Scholar] [CrossRef]
- Handakumbura, P.P.; Stanfill, B.; Rivas-Ubach, A.; Fortin, D.; Vogel, J.P.; Jansson, C. Metabotyping as a Stopover in Genome-to-Phenome Mapping. Sci. Rep. 2019, 9, 1858. [Google Scholar] [CrossRef] [PubMed]
- Skalska, A.; Beckmann, M.; Corke, F.; Savas Tuna, G.; Tuna, M.; Doonan, J.H.; Hasterok, R.; Mur, L.A.J. Metabolomic Variation Aligns with Two Geographically Distinct Subpopulations of Brachypodium distachyon before and after Drought Stress. Cells 2021, 10, 683. [Google Scholar] [CrossRef]
- Piasecka, A.; Jedrzejczak-Rey, N.; Bednarek, P. Secondary metabolites in plant innate immunity: Conserved function of divergent chemicals. New Phytol. 2015, 206, 948–964. [Google Scholar] [CrossRef]
- Weng, J.-K.; Philippe, R.N.; Noel, J.P. The Rise of Chemodiversity in Plants. Science 2012, 336, 1667–1670. [Google Scholar] [CrossRef] [PubMed]
- Vogt, T. Phenylpropanoid Biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Garvin, D.F.; Gu, Y.-Q.; Hasterok, R.; Hazen, S.P.; Jenkins, G.; Mockler, T.C.; Mur, L.A.J.; Vogel, J.P. Development of Genetic and Genomic Research Resources for Brachypodium distachyon, a New Model System for Grass Crop Research. Crop Sci. 2008, 48, S-69–S-84. [Google Scholar] [CrossRef]
- Vogel, J.P.; Garvin, D.F.; Leong, O.M.; Hayden, D.M. Agrobacterium-mediated transformation and inbred line development in the model grass Brachypodium distachyon. Plant Cell Tiss. Org. Cult. 2006, 84, 199–211. [Google Scholar] [CrossRef]
- Ingram, P.A.; Zhu, J.; Shariff, A.; Davis, I.W.; Benfey, P.N.; Elich, T. High-throughput imaging and analysis of root system architecture in Brachypodium distachyon under differential nutrient availability. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2012, 367, 1559–1569. [Google Scholar] [CrossRef]
- Cui, Y.; Lee, M.Y.; Huo, N.; Bragg, J.; Yan, L.; Yuan, C.; Li, C.; Holditch, S.J.; Xie, J.; Luo, M.-C.; et al. Fine Mapping of the Bsr1 Barley Stripe Mosaic Virus Resistance Gene in the Model Grass Brachypodium distachyon. PLoS ONE 2012, 7, e38333. [Google Scholar] [CrossRef]
- Lee, M.Y.; Yan, L.; Gorter, F.A.; Kim, B.Y.T.; Cui, Y.; Hu, Y.; Yuan, C.; Grindheim, J.; Ganesan, U.; Liu, Z.; et al. Brachypodium distachyon line Bd3-1 resistance is elicited by the barley stripe mosaic virus triple gene block 1 movement protein. J. Gen. Virol. 2012, 93, 2729–2739. [Google Scholar] [CrossRef]
- Gill, U.S.; Lee, S.; Jia, Y.; Mysore, K.S. Exploring natural variation for rice sheath blight resistance in Brachypodium distachyon. Plant Signal. Behav. 2019, 14, 1546527. [Google Scholar] [CrossRef]
- Gill, U.S.; Uppalapati, S.R.; Nakashima, J.; Mysore, K.S. Characterization of Brachypodium distachyon as a nonhost model against switchgrass rust pathogen Puccinia emaculata. BMC Plant Biol. 2015, 15, 113. [Google Scholar] [CrossRef]
- Peraldi, A.; Griffe, L.L.; Burt, C.; McGrann, G.R.D.; Nicholson, P. Brachypodium distachyon exhibits compatible interactions with Oculimacula spp. and Ramularia collo-cygni, providing the first pathosystem model to study eyespot and ramularia leaf spot diseases. Plant Pathol. 2014, 63, 554–562. [Google Scholar] [CrossRef] [Green Version]
- Kouzai, Y.; Kimura, M.; Watanabe, M.; Kusunoki, K.; Osaka, D.; Suzuki, T.; Matsui, H.; Yamamoto, M.; Ichinose, Y.; Toyoda, K.; et al. Salicylic acid-dependent immunity contributes to resistance against Rhizoctonia solani, a necrotrophic fungal agent of sheath blight, in rice and Brachypodium distachyon. New Phytol. 2018, 217, 771–783. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, X.; Yu, X.; Zhao, X.; Luo, N.; Pei, Z.; Liu, H.; Garvin, D.F. Quantitative Trait Loci Associated with Drought Tolerance in Brachypodium distachyon. Front. Plant Sci. 2017, 8, 811. [Google Scholar] [CrossRef] [PubMed]
- Bednarek, P.; Franski, R.; Kerhoas, L.; Einhorn, J.; Wojtaszek, P.; Stobiecki, M. Profiling changes in metabolism of isoflavonoids and their conjugates in Lupinus albus treated with biotic elicitor. Phytochemistry 2001, 56, 77–85. [Google Scholar] [CrossRef]
- Bednarek, P.; Schneider, B.; Svatos, A.; Oldham, N.J.; Hahlbrock, K. Structural complexity, differential response to infection, and tissue specificity of indolic and phenylpropanoid secondary metabolism in Arabidopsis roots. Plant Physiol. 2005, 138, 1058–1070. [Google Scholar] [CrossRef] [PubMed]
- Naithani, S.; Gupta, P.; Preece, J.; D’Eustachio, P.; Elser, J.L.; Garg, P.; Dikeman, D.A.; Kiff, J.; Cook, J.; Olson, A.; et al. Plant Reactome: A knowledgebase and resource for comparative pathway analysis. Nucleic Acids Res. 2019, 48, D1093–D1103. [Google Scholar] [CrossRef]
- Zhou, S.; Kremling, K.A.; Bandillo, N.; Richter, A.; Zhang, Y.K.; Ahern, K.R.; Artyukhin, A.B.; Hui, J.X.; Younkin, G.C.; Schroeder, F.C.; et al. Metabolome-Scale Genome-Wide Association Studies Reveal Chemical Diversity and Genetic Control of Maize Specialized Metabolites. Plant Cell 2019, 31, 937–955. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Minen, R.I.; Martinez, M.P.; Iglesias, A.A.; Figueroa, C.M. Biochemical characterization of recombinant UDP-sugar pyrophosphorylase and galactinol synthase from Brachypodium distachyon. Plant Physiol. Biochem. 2020, 155, 780–788. [Google Scholar] [CrossRef]
- Kroymann, J.; Textor, S.; Tokuhisa, J.G.; Falk, K.L.; Bartram, S.; Gershenzon, J.; Mitchell-Olds, T. A gene controlling variation in Arabidopsis glucosinolate composition is part of the methionine chain elongation pathway. Plant Physiol. 2001, 127, 1077–1088. [Google Scholar] [CrossRef]
- Pan, L.; Meng, C.; Wang, J.; Ma, X.; Fan, X.; Yang, Z.; Zhou, M.; Zhang, X. Integrated omics data of two annual ryegrass (Lolium multiflorum L.) genotypes reveals core metabolic processes under drought stress. BMC Plant Biol. 2018, 18, 26. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, N.; Takano, T.; Tanaka, K.; Ishige, T.; Terashima, S.; Endo, C.; Kurusu, T.; Yajima, S.; Yano, K.; Tada, Y. Comprehensive analysis of transcriptome response to salinity stress in the halophytic turf grass Sporobolus virginicus. Front. Plant Sci. 2015, 6, 241. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.M.; Zerbe, P. Specialized diterpenoid metabolism in monocot crops: Biosynthesis and chemical diversity. Phytochemistry 2020, 172, 112289. [Google Scholar] [CrossRef] [PubMed]
- Hardtke, C.S. Gibberellin Signaling: GRASs Growing Roots Dispatch. Curr. Biol. 2003, 13, R366–R367. [Google Scholar] [CrossRef]
- Reyes-Hernández, B.J.; Srivastava, A.C.; Ugartechea-Chirino, Y.; Shishkova, S.; Ramos-Parra, P.A.; Lira-Ruan, V.; Díaz de la Garza, R.I.; Dong, G.; Moon, J.-C.; Blancaflor, E.B.; et al. The root indeterminacy-to-determinacy developmental switch is operated through a folate-dependent pathway in Arabidopsis thaliana. New Phytol. 2014, 202, 1223–1236. [Google Scholar] [CrossRef] [PubMed]
- Ashihara, H.; Stasolla, C.; Fujimura, T.; Crozier, A. Purine salvage in plants. Phytochemistry 2018, 147, 89–124. [Google Scholar] [CrossRef]
- Trabucco, G.M.; Matos, D.A.; Lee, S.J.; Saathoff, A.J.; Priest, H.D.; Mockler, T.C.; Sarath, G.; Hazen, S.P. Functional characterization of cinnamyl alcohol dehydrogenase and caffeic acid O-methyltransferase in Brachypodium distachyon. BMC Biotechnol. 2013, 13, 61. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Degtyarenko, K.; De Matos, P.; Ennis, M.; Hastings, J.; Zbinden, M.; McNaught, A.; Alcantara, R.; Darsow, M.; Guedj, M.; Ashburner, M. ChEBI: A database and ontology for chemical entities of biological interest. Nucleic Acids Res. 2008, 36, D344–D350. [Google Scholar] [CrossRef]
- Kushnir, M.M.; Urry, F.M.; Frank, E.L.; Roberts, W.L.; Shushan, B. Analysis of catecholamines in urine by positive-ion electrospray tandem mass spectrometry. Clin. Chem. 2002, 48, 323–331. [Google Scholar] [CrossRef]
- Thiele, B.; Füllner, K.; Stein, N.; Oldiges, M.; Kuhn, A.J.; Hofmann, D. Analysis of amino acids without derivatization in barley extracts by LC-MS-MS. Anal. Bioanal. Chem. 2008, 391, 2663–2672. [Google Scholar] [CrossRef]
- Ogura, Y.; Ishihara, A.; Iwamura, H. Induction of Hydroxycinnamic Acid Amides and Tryptophan by Jasmonic Acid, Abscisic Acid and Osmotic Stress in Barley Leaves. Z. Naturforsch. C 2001, 56, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Sarnoski, P.J.; Boyer, R.R.; O’Keefe, S.F. Application of proanthocyanidins from peanut skins as a natural yeast inhibitory agent. J. Food Sci. 2012, 77, M242–M249. [Google Scholar] [CrossRef] [PubMed]
- Stobiecki, M.; Skirycz, A.; Kerhoas, L.; Kachlicki, P.; Muth, D.; Einhorn, J.; Mueller-Roeber, B. Profiling of phenolic glycosidic conjugates in leaves of Arabidopsis thaliana using LC/MS. Metabolomics 2006, 2, 197–219. [Google Scholar] [CrossRef]
- Parveen, I.; Wilson, T.; Donnison, I.S.; Cookson, A.R.; Hauck, B.; Threadgill, M.D. Potential sources of high value chemicals from leaves, stems and flowers of Miscanthus sinensis ‘Goliath’ and Miscanthus sacchariflorus. Phytochemistry 2013, 92, 160–167. [Google Scholar] [CrossRef]
- Klausen, K.; Mortensen, A.G.; Laursen, B.; Haselmann, K.F.; Jespersen, B.M.; Fomsgaard, I.S. Phenolic Compounds in Different Barley Varieties: Identification by Tandem Mass Spectrometry (QStar) and NMR; Quantification by Liquid Chromatography Triple Quadrupole-Linear Ion Trap Mass Spectrometry (Q-Trap). Nat. Prod. Commun. 2010, 5, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhnert, N. Hierarchical scheme for LC-MSn identification of chlorogenic acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef]
- Cuyckens, F.; Claeys, M. Determination of the glycosylation site in flavonoid mono-O-glycosides by collision-induced dissociation of electrospray-generated deprotonated and sodiated molecules. J. Mass Spectrom. 2005, 40, 364–372. [Google Scholar] [CrossRef]
- Piasecka, A.; Sawikowska, A.; Krajewski, P.; Kachlicki, P. Combined mass spectrometric and chromatographic methods for in-depth analysis of phenolic secondary metabolites in barley leaves. J. Mass Spectrom. 2015, 50, 513–532. [Google Scholar] [CrossRef]
- Handrick, V.; Vogt, T.; Frolov, A. Profiling of hydroxycinnamic acid amides in Arabidopsis thaliana pollen by tandem mass spectrometry. Anal. Bioanal. Chem. 2010, 398, 2789–2801. [Google Scholar] [CrossRef]
- Moheb, A.; Ibrahim, R.K.; Roy, R.; Sarhan, F. Changes in wheat leaf phenolome in response to cold acclimation. Phytochemistry 2011, 72, 2294–2307. [Google Scholar] [CrossRef]
- von Ropenack, E.; Parr, A.; Schulze-Lefert, P. Structural analyses and dynamics of soluble and cell wall-bound phenolics in a broad spectrum resistance to the powdery mildew fungus in barley. J. Biol. Chem. 1998, 273, 9013–9022. [Google Scholar] [CrossRef] [PubMed]
- Ferreres, F.; Andrade, P.B.; Valentão, P.; Gil-Izquierdo, A. Further knowledge on barley (Hordeum vulgare L.) leaves O-glycosyl-C-glycosyl flavones by liquid chromatography-UV diode-array detection-electrospray ionisation mass spectrometry. J. Chromatogr. 2008, 1182, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Lallemand, L.A.; Zubieta, C.; Lee, S.G.; Wang, Y.; Acajjaoui, S.; Timmins, J.; McSweeney, S.; Jez, J.M.; McCarthy, J.G.; McCarthy, A.A. A Structural Basis for the Biosynthesis of the Major Chlorogenic Acids Found in Coffee. Plant Physiol. 2012, 160, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Smirnoff, N. Ascorbic acid metabolism and functions: A comparison of plants and mammals. Free Radic. Biol. Med. 2018, 122, 116–129. [Google Scholar] [CrossRef] [PubMed]
- Parveen, I.; Winters, A.; Threadgill, M.D.; Hauck, B.; Morris, P. Extraction, structural characterisation and evaluation of hydroxycinnamate esters of orchard grass (Dactylis glomerata) as substrates for polyphenol oxidase. Phytochemistry 2008, 69, 2799–2806. [Google Scholar] [CrossRef]
- Desmet, S.; Saeys, Y.; Verstaen, K.; Dauwe, R.; Kim, H.; Niculaes, C.; Fukushima, A.; Goeminne, G.; Vanholme, R.; Ralph, J.; et al. Maize specialized metabolome networks reveal organ-preferential mixed glycosides. Comput. Struct. Biotechnol. J. 2021, 19, 1127–1144. [Google Scholar] [CrossRef] [PubMed]
- Kuczkowiak, U.; Petereit, F.; Nahrstedt, A. Hydroxycinnamic Acid Derivatives Obtained from a Commercial Crataegus Extract and from Authentic Crataegus spp. Sci. Pharm. 2014, 82, 835–846. [Google Scholar] [CrossRef]
- Renault, H.; Alber, A.; Horst, N.A.; Basilio Lopes, A.; Fich, E.A.; Kriegshauser, L.; Wiedemann, G.; Ullmann, P.; Herrgott, L.; Erhardt, M.; et al. A phenol-enriched cuticle is ancestral to lignin evolution in land plants. Nat. Commun. 2017, 8, 14713. [Google Scholar] [CrossRef]
- Krzyzanowska-Kowalczyk, J.; Pecio, L.; Moldoch, J.; Ludwiczuk, A.; Kowalczyk, M. Novel Phenolic Constituents of Pulmonaria officinalis L. LC-MS/MS Comparison of Spring and Autumn Metabolite Profiles. Molecules 2018, 23, 2277. [Google Scholar] [CrossRef]
- Cadahía, E.; Fernández de Simón, B.; Aranda, I.; Sanz, M.; Sánchez-Gómez, D.; Pinto, E. Non-targeted Metabolomic Profile of Fagus Sylvatica L. Leaves using Liquid Chromatography with Mass Spectrometry and Gas Chromatography with Mass Spectrometry. Phytochem. Anal. 2015, 26, 171–182. [Google Scholar] [CrossRef]
- Zeng, Z.C.; Liu, C.M.; Luo, S.J.; Chen, J.; Gong, E.S. The Profile and Bioaccessibility of Phenolic Compounds in Cereals Influenced by Improved Extrusion Cooking Treatment. PLoS ONE 2016, 11, e0161086. [Google Scholar] [CrossRef]
- Carere, J.; Powell, J.; Fitzgerald, T.; Kazan, K.; Gardiner, D.M. BdACT2a encodes an agmatine coumaroyl transferase required for pathogen defence in Brachypodium distachyon. Physiol. Mol. Plant Pathol. 2018, 104, 69–76. [Google Scholar] [CrossRef]
- Jin, S.; Yoshida, M. Antifungal Compound, Feruloylagmatine, Induced in Winter Wheat Exposed to a Low Temperature. Biosci. Biotechnol. Biochem. 2000, 64, 1614–1617. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, P.C.; Green, P.W.C.; Farrell, I.W.; Brankin, A.; Mvumi, B.M.; Belmain, S.R. Novel Agmatine Derivatives in Maerua edulis With Bioactivity Against Callosobruchus maculatus, a Cosmopolitan Storage Insect Pest. Front. Plant Sci. 2018, 9, 1506. [Google Scholar] [CrossRef] [PubMed]
- Wojakowska, A.; Perkowski, J.; Goral, T.; Stobiecki, M. Structural characterization of flavonoid glycosides from leaves of wheat (Triticum aestivum L.) using LC/MS/MS profiling of the target compounds. J. Mass Spectrom. 2013, 48, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Wojakowska, A.; Piasecka, A.; García-López, P.M.; Zamora-Natera, F.; Krajewski, P.; Marczak, Ł.; Kachlicki, P.; Stobiecki, M. Structural analysis and profiling of phenolic secondary metabolites of Mexican lupine species using LC–MS techniques. Phytochemistry 2013, 92, 71–86. [Google Scholar] [CrossRef]
- Asenstorfer, R.E.; Wang, Y.; Mares, D.J. Chemical structure of flavonoid compounds in wheat (Triticum aestivum L.) flour that contribute to the yellow colour of Asian alkaline noodles. J. Cereal Sci. 2006, 43, 108–119. [Google Scholar] [CrossRef]
- Frost, S.; Harborne, J.B.; King, L. Identification of the flavonoids in five chemical races of cultivated barley. Hereditas 1977, 85, 163–168. [Google Scholar] [CrossRef]
- Kachlicki, P.; Einhorn, J.; Muth, D.; Kerhoas, L.; Stobiecki, M. Evaluation of glycosylation and malonylation patterns in flavonoid glycosides during LC/MS/MS metabolite profiling. J. Mass Spectrom. 2008, 43, 572–586. [Google Scholar] [CrossRef]
- Ferreres, F.; Gil-Izquierdo, A.; Andrade, P.B.; Valentao, P.; Tomas-Barberan, F.A. Characterization of C-glycosyl flavones O-glycosylated by liquid chromatography-tandem mass spectrometry. J. Chromatogr. 2007, 1161, 214–223. [Google Scholar] [CrossRef]
- Guijas, C.; Montenegro-Burke, J.R.; Domingo-Almenara, X.; Palermo, A.; Warth, B.; Hermann, G.; Koellensperger, G.; Huan, T.; Uritboonthai, W.; Aisporna, A.E.; et al. METLIN: A Technology Platform for Identifying Knowns and Unknowns. Anal. Chem. 2018, 90, 3156–3164. [Google Scholar] [CrossRef] [PubMed]
- Falcone Ferreyra, M.L.; Rodriguez, E.; Casas, M.I.; Labadie, G.; Grotewold, E.; Casati, P. Identification of a Bifunctional Maize C- and O-Glucosyltransferase. J. Biol. Chem. 2013, 288, 31678–31688. [Google Scholar] [CrossRef] [PubMed]
- Brazier-Hicks, M.; Evans, K.M.; Gershater, M.C.; Puschmann, H.; Steel, P.G.; Edwards, R. The C-Glycosylation of Flavonoids in Cereals. J. Biol. Chem. 2009, 284, 17926–17934. [Google Scholar] [CrossRef]
- Brauch, D.; Porzel, A.; Schumann, E.; Pillen, K.; Mock, H.-P. Changes in isovitexin-O-glycosylation during the development of young barley plants. Phytochemistry 2018, 148, 11–20. [Google Scholar] [CrossRef]
- Ohkawa, M.; Kinjo, J.; Hagiwara, Y.; Hagiwara, H.; Ueyama, H.; Nakamura, K.; Ishikawa, R.; Ono, M.; Nohara, T. Three New Anti-Oxidative Saponarin Analogs from Young green Barley Leaves. Chem. Pharm. Bull. 1998, 46, 1887–1890. [Google Scholar] [CrossRef]
- Norbaek, R.; Aaboer, D.B.F.; Bleeg, I.S.; Christensen, B.T.; Kondo, T.; Brandt, K. Flavone C-glycoside, phenolic acid, and nitrogen contents in leaves of barley subject to organic fertilization treatments. J. Agric. Food Chem. 2003, 51, 809–813. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, A.; Ogura, Y.; Tebayashi, S.; Iwamura, H. Jasmonate-induced changes in flavonoid metabolism in barley (Hordeum vulgare) leaves. Biosci. Biotechnol. Biochem. 2002, 66, 2176–2182. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.Z.; Sun, J.H.; Chen, P.; Monagas, M.J.; Harnly, J.M. UHPLC-PDA-ESUHRMSn Profiling Method To Identify and Quantify Oligomeric Proanthocyanidins in Plant Products. J. Agric. Food Chem. 2014, 62, 9387–9400. [Google Scholar] [CrossRef]
- Rue, E.A.; Rush, M.D.; van Breemen, R.B. Procyanidins: A comprehensive review encompassing structure elucidation via mass spectrometry. Phytochem. Rev. 2018, 17, 1–16. [Google Scholar] [CrossRef]
- Zhu, F. Proanthocyanidins in cereals and pseudocereals. Crit. Rev. Food Sci. Nutr. 2019, 59, 1521–1533. [Google Scholar] [CrossRef]
- Verardo, V.; Cevoli, C.; Pasini, F.; Gómez-Caravaca, A.M.; Marconi, E.; Fabbri, A.; Caboni, M.F. Analysis of Oligomer Proanthocyanidins in Different Barley Genotypes Using High-Performance Liquid Chromatography–Fluorescence Detection–Mass Spectrometry and Near-Infrared Methodologies. J. Agric. Food Chem. 2015, 63, 4130–4137. [Google Scholar] [CrossRef] [PubMed]
- Mochida, K.; Uehara-Yamaguchi, Y.; Takahashi, F.; Yoshida, T.; Sakurai, T.; Shinozaki, K. Large-scale collection and analysis of full-length cDNAs from Brachypodium distachyon and integration with Pooideae sequence resources. PLoS ONE 2013, 8, e75265. [Google Scholar]
- Ölschläger, C.; Regos, I.; Zeller, F.J.; Treutter, D. Identification of galloylated propelargonidins and procyanidins in buckwheat grain and quantification of rutin and flavanols from homostylous hybrids originating from F. Esculentum × F. homotropicum. Phytochemistry 2008, 69, 1389–1397. [Google Scholar] [CrossRef]
- Stauss, R. Compendium of Growth Stage Identification Keys for Mono- and Dicotyledoneus Plants: Extended BBCH Scale; Ciba-Geigy: Basel, Switzerland, 1994; p. 94. [Google Scholar]
- Afendi, F.M.; Okada, T.; Yamazaki, M.; Hirai-Morita, A.; Nakamura, Y.; Nakamura, K.; Ikeda, S.; Takahashi, H.; Altaf-Ul-Amin, M.; Darusman, L.K.; et al. KNApSAcK Family Databases: Integrated Metabolite-Plant Species Databases for Multifaceted Plant Research. Plant Cell Physiol. 2012, 53, e1. [Google Scholar] [CrossRef] [PubMed]
- Goodman, J. Reaxys. J. Chem. Inf. Model. 2009, 49, 2897–2898. [Google Scholar] [CrossRef]
- Sayers, E.W.; Agarwala, R.; Bolton, E.E.; Brister, J.R.; Canese, K.; Clark, K.; Connor, R.; Fiorini, N.; Funk, K.; Hefferon, T.; et al. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2019, 47, D23–D28. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Oresic, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biological Pathway (KEGG) | Total | Hits | FDR | Impact |
|---|---|---|---|---|
| Flavonoid biosynthesis | 47 | 40 | 4.31 × 10−6 | 0.75288 |
| Galactose metabolism | 27 | 25 | 2.72 × 10−5 | 1 |
| Amino sugar and nucleotide sugar metabolism | 50 | 39 | 0.000177 | 0.92767 |
| Valine, leucine and isoleucine biosynthesis | 22 | 20 | 0.000426 | 0.99998 |
| Pentose and glucuronate interconversions | 17 | 16 | 0.000971 | 1 |
| Flavone and flavonol biosynthesis | 12 | 12 | 0.001539 | 1 |
| 2-Oxocarboxylic acid metabolism | 12 | 12 | 0.001539 | 1 |
| Pentose phosphate pathway | 19 | 16 | 0.010995 | 0.9532 |
| Purine metabolism | 63 | 42 | 0.011504 | 0.67941 |
| Tyrosine metabolism | 18 | 15 | 0.014584 | 0.79191 |
| Ascorbate and aldarate metabolism | 18 | 15 | 0.014584 | 0.8806 |
| Vitamin B6 metabolism | 11 | 10 | 0.026026 | 0.96153 |
| Biological Pathway Enrichment (KEGG) | Total | Hits | FDR | Impact | |
|---|---|---|---|---|---|
| Roots | Galactose metabolism | 27 | 25 | 7.41 × 10−7 | 1 |
| Diterpenoid biosynthesis | 47 | 32 | 0.001253 | 0.61959 | |
| Flavonoid biosynthesis | 47 | 32 | 0.001253 | 0.64212 | |
| Valine, leucine and isoleucine biosynthesis | 22 | 18 | 0.001253 | 0.68294 | |
| Caffeine metabolism | 10 | 10 | 0.001636 | 0 | |
| Pentose phosphate pathway | 19 | 15 | 0.007666 | 0.85473 | |
| One carbon pool by folate | 8 | 8 | 0.007666 | 1 | |
| Flavone and flavonol biosynthesis | 12 | 10 | 0.025063 | 0 | |
| 2-Oxocarboxylic acid metabolism | 12 | 10 | 0.025063 | 0 | |
| Leaves | Flavonoid biosynthesis | 47 | 42 | 2.39 × 10−6 | 0.7644 |
| Galactose metabolism | 27 | 24 | 0.001979 | 1 | |
| Valine, leucine and isoleucine biosynthesis | 22 | 20 | 0.003065 | 0.8355 | |
| Histidine metabolism | 17 | 16 | 0.004891 | 1 | |
| Flavone and flavonol biosynthesis | 12 | 12 | 0.0054 | 0 | |
| 2-Oxocarboxylic acid metabolism | 12 | 12 | 0.0054 | 0 | |
| Pentose phosphate pathway | 19 | 17 | 0.007434 | 0.99999 | |
| Vitamin B6 metabolism | 11 | 11 | 0.007908 | 0.99999 | |
| Diterpenoid biosynthesis | 47 | 34 | 0.028047 | 0.69919 | |
| Spikes | Flavonoid biosynthesis | 47 | 41 | 3.39 × 10−5 | 0.75288 |
| Galactose metabolism | 27 | 25 | 0.000367 | 1 | |
| Pentose phosphate pathway | 19 | 18 | 0.002628 | 0.99999 | |
| Valine, leucine and isoleucine biosynthesis | 22 | 20 | 0.003264 | 0.8355 | |
| Flavone and flavonol biosynthesis | 12 | 12 | 0.006808 | 0 | |
| 2-Oxocarboxylic acid metabolism | 12 | 12 | 0.006808 | 0 | |
| Vitamin B6 metabolism | 11 | 11 | 0.011173 | 0.99999 | |
| Pentose and glucuronate interconversions | 17 | 15 | 0.021023 | 0.85716 | |
| Histidine metabolism | 17 | 15 | 0.021023 | 1 | |
| Cysteine and methionine metabolism | 46 | 34 | 0.021023 | 0.75798 |
| # | Fragmentation Pathway in MSn [m/z] | Identification | Exact mass of [M+H]+ or [M−H]; [Da] | ∆ ppm | Chemical Formula | λmax [nm] | Leaves | Roots | Spikes | ChEBI | Identification Level | References | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Negative Ionization | Positive Ionization | Ion Type | Measured | Calculated | |||||||||||
| 1 | MS2: 137, 90, 64 | Dopamine | [M+H]+ | 154.08638 | 154.0864 | 0.8102 | C8H11NO2 | * | * | * | 18243 | 2 | [49] | ||
| 2 | MS2: 165, 147, 136 MS3: 147, 123 | Tyrosine | [M+H]+ | 182.081 | 182.0812 | −1.1348 | C9H11NO3 | * | * | 18186 | 2 | [50] | |||
| 3 | MS2: 116, 86 | Leucine (Isoleucine) | [M+H]+ | 132.1018 | 132.1019 | −0.5152 | C6H13NO2 | * | * | * | 25017 | 2 | [50] | ||
| 4 | MS2: 163, 89 MS3: 131 | N-Caffeoyl-putrescine | [M+H]+ | 251.13862 | 251.139 | −1.589 | C13H18N2O3 | * | * | * | 17417 | 3 | [51] | ||
| 5 | MS2: 145, 120 MS3: 79 | Phenylalanine | [M+H]+ | 166.086 | 166.0863 | −1.3614 | C9H11NO2 | 260 | * | * | * | 28044 | 2 | [50] | |
| 6 | MS2: 727, 609, 559, 541, 483, 423, 303 MS3: 559, 423, 303 | (epi)Gallocatechin trimer | [M−H]− | 913.18583 | 913.1833 | 2.791 | C45H38O21 | * | * | * | 3 | [52] | |||
| 7 | MS2: 771, 711, 593, 543, 467, 303, 289 MS3: 697, 543, 289 | Proanthocyanidins trimer A-type | [M−H]− | 897.19086 | 897.1884 | 2.779 | C45H38O20 | * | 3 | [52] | |||||
| 8 | MS2: 305, 265, 223, 205, 161, 143, 125 MS3: 223, 205 | Caffeic acid derivative | [M−H]− | 367.12504 | 367.1246 | 1.24 | C17H20O9 | * | 149782 | 2 | [53] | ||||
| 9 | MS2: 269, 209, 167 | Vanilic acid-hexoside | [M−H]− | 329.0883 | 329.0878 | 1.5932 | C14H18O9 | * | * | 2 | [54] | ||||
| 10 | MS2: 218, 89 | p-Coumaroyl-N-putrescine | [M+H]+ | 235.1441 | 235.1441 | 0.0370 | C13H18N2O | 290sh | * | * | 70431 | 2 | [51] | ||
| 11 | MS2: 248, 177, 144, 114, 98 MS3: 145 | Feruloyl-N-putrescine | [M+H]+ | 265.15424 | 265.1547 | −1.16337 | C14H20N2O3 | * | * | 9299 | 3 | [51] | |||
| 12 | MS2: 233, 119 MS3: 117, 93 | MS2: 218, 176, 147, 114, 89, 73 MS3: 147 | p-Coumaroyl-N-putrescine | [M+H]+ | 235.14375 | 235.1441 | −1.507 | C13H18N2O | * | * | 3 | [51] | |||
| 13 | MS2: 203, 159, 142, 116 | MS2: 188, 146 MS3: 146, 118 | Tryptophan | [M+H]+ | 205.097 | 205.0972 | −0.9744 | C11H12N2O2 | 285 | * | * | * | 27897 | 1 | Std |
| 14 | MS2: 299, 239, 209, 179, 137 MS3: | Hydroxybenzoic acid hexoside | [M−H]− | 299.0765 | 299.07728 | −2.349 | C13H16O8 | 282 | * | 16741 | 2 | [54] | |||
| 15 | MS2: 574, 467, 425, 407, 289 MS3: 245, 205, 177 | MS2: 595, 443, 427, 317, 307, 289 MS3: 289, 247 | Prodelphinidin B-type | [M−H]− | 593.13153 | 593.1301 | 2.472 | C30H26O13 | * | 75664# | 2 | [52,55] | |||
| 16 | MS2: 315, 153 MS3: 108 | Dihydroxybenzoic acid hexoside | [M−H]− | 315.0718 | 315.0722 | −1.1026 | C13H16O9 | 286 | * | 2 | [54] | ||||
| 17 | MS2: 160 MS3: 134, 132, 115 | Serotonin | [M+H]+ | 177.1019 | 177.1022 | −2.0138 | C₁₀H₁₂N₂O | 275, 298sh | * | * | 28790 | 1 | Std | ||
| 18 | MS2: 179, 135 MS3:117, 89, 75 | MS2: 163, 136,137 MS3: 136, 118 | Caffeoylthreonic acid | [M−H]− | 297.0611 | 297.0616 | −1.664 | C13H14O8 | * | * | * | 2 | [54] | ||
| 19 | MS2: 461, 225, 153 MS3: 108, 90 | Dihydroxybenzoic acid hexosyldeoxyhexoside | [M−H]− | 461.1299 | 461.1301 | −0.459 | C19H26O13 | 281 | * | 3 | [54] | ||||
| 20 | MS2: 863, 755, 695, 591, 407, 289, 243 MS3: 524, 283 | Catechin-gallocatechin-catechin | [M−H]− | 881.19622 | 881.1935 | 3.141 | C45H38O19 | * | 3 | [52,55] | |||||
| 21 | MS2: 179, 134, 119 | Caffeic acid | [M−H]− | 179.03439 | 179.035 | −3.307 | C9H8O4 | 304sh | * | * | 36281 | 1 | Std | ||
| 22 | MS2: 299, 239, 197, 153, 138 MS3: 182, 153,138, 121 | Syringic acid-hexoside | [M−H]− | 359.09827 | 359.09782 | 1.2393 | C15H20O10 | * | * | 2 | [54] | ||||
| 23 | MS2: 248, 177, 145 MS3: 177 | feruloyl-N-putrescine | [M+H]+ | 265.1548 | 265.1547 | 0.3229 | C14H20N2O3 | * | * | 3 | [51] | ||||
| 24 | MS2: 305, 289, 241, 225, 139 MS3: 223, 184, 139, 97 | MS2: 337, 305, 185, 153 MS3: 153, 125 | (epi)Gallocatechin O-hydroxybenzoate | [M−H]− | 425.08831 | 425.0878 | 1.187 | C22H18O9 | * | * | * | 3 | [52,55] | ||
| 25 | MS2: 439, 325, 305, 289, 191, 163, 131 MS3: 115 | (epi)Gallocatechin 3-O-gallate | [M−H]− | 457.07885 | 457.0776 | 3.859 | C22H18O11 | * | `3 | [52,55] | |||||
| 26 | MS2: 323, 193, 173, 135 MS3: 149, 135 | MS2: 353, 309, 274, 238, 177, 145 MS3: 145 | 5-Feruloylquinic acid | [M−H]− | 367.1028 | 367.1035 | −1.831 | C17H20O9 | 280sh, 320 | * | * | 86388# | 2 | [56] | |
| 27 | MS2: 607, 589, 333, 203 MS3: 333, 203 | Prodelphinidin A-type dimer (Prodelphinidin A1) | [M−H]− | 607.11102 | 607.1093 | 2.786 | C30H24O14 | * | [52,55] | ||||||
| 28 | MS2: 463, 301 | MS2: 465, 303, 229, 201 MS3: 303 | Quercetin di-O-hexoside | [M−H]− | 625.14292 | 625.141 | 3.035 | C27H30O17 | 253, 353 | * | 3 | [53,57] | |||
| 29 | MS2: 353, 179, 173 MS3: 109, 93 | MS2: 192, 165, 146 | 4-Caffeoylquinic acid | [M−H]− | 353.06743 | 353.0667 | 2.136 | C16H18O9 | 340sh, 305 | * | * | 75491 | 2 | [56] | |
| 30 | MS2: 665, 635, 563, 503, 473, 443, 383, 353 MS3: 353, 297 | Apigenin 6-C-hexoside-8-C-pentoside 7-O-hexoside | [M−H]− | 725.1924 | 725.1898 | −3.4967 | C32H38O19 | 266, 335 | 3 | [58] | |||||
| 31 | MS2: 489, 399, 369 MS3: 369 | Luteolin 6,8-di-C-hexoside | [M−H]− | 609.1435 | 609.1450 | −2.5141 | C27H30O16 | 262, 345 | * | * | 6553 | 2 | [58] | ||
| 32 | MS2: 577, 407, 289 MS3: 289, 143 | MS2: 579, 427, 291 | Procyanidin B-type dimer | [M−H]− | 577.13679 | 577.1351 | 2.843 | C30H26O12 | * | 75630 | 2 | [52,55] | |||
| 33 | MS3: 245, 205, 137, 125 MS3: 203 | MS2: 157, 139, 123 | (epi)Catechin | [M−H]− | 289.07255 | 289.0718 | 2.728 | C15H14O6 | * | 23053 | 2 | [55] | |||
| 34 | MS2:595, 483, 423, 305, 283 MS3 (609-483): 303, 179 MS3 (609-305): 289, 143 | MS2: 611, 443, 317 | Prodelphinidin B-type | [M−H]− | 609.12659 | 609.125 | 2.646 | C30H26O14 | * | 2 | [52] | ||||
| 35 | MS2: 275, 235, 218, 147, 118 MS3: 218, 147, 112 | N-p-Coumaroyl spermidine | [M+H]+ | 292.07275 | 292.0717 | 3.706 | C34H37N3O6 | 285sh | * | 2 | [59] | ||||
| 36 | MS2: 609, 301 MS3: 301, 272 | MS2: 627, 611, 465, 303 MS3: 369, 303 | Quercetin O-deoxyhexosylhexoside-O-hexoside | [M−H]− | 771.1991 | 771.189 | 0.1549 | C33H40O21 | 255, 353 | * | 3 | [58] | |||
| 37 | MS2: 463, 301 | MS2: 610, 551, 465, 303 MS3: 303 | Quercetin di-O-hexoside II | [M−H]− | 625.13971 | 625.141 | −2.099 | C27H30O17 | 253, 353 | * | * | 3 | [53,57] | ||
| 38 | MS2: 193, 134 | Ferulic acid | [M−H]− | 193.05024 | 193.0506 | −2.031 | C10H10O4 | 300sh, 326 | * | * | 17620 | 3 | [53,57] | ||
| 39 | MS2: 179, 135, 117 MS3:117, 89 | MS2: 299, 136 | Apigenin 7,4’-dimethyl ether | [M−H]− | 297.07752 | 297.0768 | 2.266 | C17H14O5 | * | 17620 | 2 | [60] | |||
| 40 | MS2: 233, 119 | MS2: 260, 217, 147, 114 MS3: 217, 98 | p-Coumaroylagmatine | [M+H]+ | 277.16547 | 277.1659 | −1.560 | C14H20N4O2 | 295sh | * | * | * | 32818 | 2 | [54] |
| 41 | MS2: 489, 447, 285, 254 MS3: 285, 254 | MS2: 567, 449, 287 | Luteolin di-O-hexoside | [M−H]− | 609.1477 | 609.1461 | 2.614 | C27H30O16 | 267, 348 | * | 2 | [61] | |||
| 42 | MS2: 311, 193, 149, 135 MS3: 135, 119 | Feruloylthreonic acid | [M−H]− | 311.07697 | 311.0772 | −0.87 | C14H16O8 | * | 3 | [57] | |||||
| 43 | MS2: 193, 173, MS3: 109, 93 | MS2: 404, 369, 277, 193 | 4-Feruloylquinic acid | [M−H]− | 367.1037 | 367.1035 | 0.7349 | C17H20O9 | * | 3 | [54] | ||||
| 44 | MS2: 562, 519, 477, 315 MS3: 357, 315, 285, 243, 199 | MS2: 641, 479, 317, 286 | Isorhamnetin di-O-hexoside | [M−H]− | 639.15509 | 639.1567 | −2.476 | C28H32O17 | 259, 369 | * | * | 60078 | 2 | [56] | |
| 45 | MS2: 695, 635, 593, 454, 473, 413, 311, 249 MS3: 473, 413 | Chrysoeriol 6-C-hexoside-8-C-pentoside 7-O-hexoside | [M−H]− | 755.20585 | 755.204 | 2.428 | C33H40O20 | 250, 348 | * | * | 3 | [57] | |||
| 46 | MS2: 519, 447, 357, 327 | MS2: 532, 464, 449, 431, 383, 353, 329, 299 MS3: 432, 413, 383, 353, 329, 320, 299 | Orientin 7-O-hexoside | [M−H]− | 609.1439 | 609.1450 | −1.8549 | C27H30O16 | 266, 349 | * | 3 | [58] | |||
| 47 | MS2: 369, 325, 163, 145, 119 MS3: 117, 95 | Sinapoyl-homovanillic acid | [M−H]− | 387.10814 | 387.1085 | −1.035 | C20H20O8 | * | 3 | [58] | |||||
| 48 | MS2: 191, 179, 173, MS3: 155, 111, 93, 71 | MS2: 146, 119, 79 | 4-p-Coumaroylquinic acid | [M−H]− | 337.0939 | 337.0929 | 2.6396 | C16H18O8 | 290sh | * | * | 1945 | 3 | [54] | |
| 49 | MS2: 290, 247, 232, 177, 152, 145, 114 MS3: 273, 247, 230, 177, 115 MS4: 145, 113 | Feruloylagmatine | [M+H]+ | 307.17636 | 307.1765 | −0.349 | C15H22N4O3 | 290sh, 320 | * | 1945 | 2 | [56] | |||
| 50 | MS2: 771, 651, 609, 429, 357, 327 MS3: 357, 327, 299 | Isoorientin 2”,6”-di-O-hexoside | [M−H]− | 771.19946 | 771.1989 | 0.6895 | C33H40O21 | 270, 344 | * | 75544 | 2 | [61] | |||
| 51 | MS2: 489, 447, 357, 327 MS3: 357, 327, 299 | MS2: 593, 449, 383, 329, 299 MS3: 431, 383, 353, 299 | Isoorientin 7-O-glucoside | [M−H]− | 609.1433 | 609.1450 | −2.7680 | C27H30O16 | 268, 348 | * | 75514 | 1 | Std; [58] | ||
| 52 | MS2: 489, 447, 327, 285, 255 MS3: 284, 226 | MS2: 449, 287, 269 MS3: 287, 259, 213 MS4: 213, 153, 133 | Luteolin 3′,7-di-O-glucoside | [M−H]− | 609.1475 | 609.1461 | 2.285 | C27H30O16 | 269, 343 | * | 75514 | 1 | Std | ||
| 53 | MS2: 593 MS3: 285, 185, 153, 131 | MS2: 595, 491, 449, 335, 311, 287 MS3: 449, 287 | Luteolin O-hexosyldeoxyhexoside-O-hexoside | [M−H]− | 755.20408 | 755.204 | 0.084 | C33H40O20 | 266, 349 | * | * | 3 | [57] | ||
| 54 | MS2: 623, 447, 315, 299 MS3: 357, 315, 299, 271, 255, 227 | MS2: 657, 641, 625, 609, 479, 317, 302, 273 MS3: 342, 317, 273 | Isorhamnetin O-hexosyldeoxyhexoside-O-hexoside | [M−H]− | 785.21381 | 785.2146 | −0.982 | C34H42O21 | 259, 369 | * | * | 3 | [53,57] | ||
| 55 | MS2: 477, 357, 315, 255, 217 MS3: 153 | Isorhamnetin hexoside | [M−H]− | 477.10435 | 477.1038 | 1.05 | C22H22O12 | 252, 369 | * | * | 3 | [53,57] | |||
| 56 | MS2: 315, 255 MS3: 153 | Isorhamnetin deoxyhexosylhexoside | [M−H]− | 623.1624 | 623.1618 | 1.0327 | C28H32O16 | - | * | 75752 or 75758 | 3 | [57] | |||
| 57 | MS2: 575, 502, 473, 413, 383 MS3 (593-473): 383, 353, 311 MS3 (593-502): 413, 383, 312 | MS2: 577, 559, 541, 529, 499, 457, 427 MS3 (595-529): 511, 427, 367 MS3 (595-577): 559, 529, 511, 481, 445, 427, 409, 380 | Apigenin 6,8-di-C-hexoside | [M−H]− | 593.1505 | 593.1512 | −1.2503 | C27H30O15 | 269, 339 | * | 3 | [53,57] | |||
| 58 | MS2:561, 519, 489, 459, 429, 399, 369 MS3:399, 369 MS4:341, 297 | MS2: 563, 545, 515, 497, 443, 413 MS3: 545, 515, 497, 485, 467, 395 MS4 (563-497): 413, 395, 312 MS4 (563-545): 509, 497, 467 | Luteolin 6-C-pentoside-8-C-hexoside | [M−H]− | 579.1492 | 579.1501 | −1.489 | C26H28O15 | 269, 348 | * | * | 69814 | 2 | [62] | |
| 59 | MS2: 399, 387, 205, 181 MS3: 372, 203 | Sinapoyl-homovanillic acid derivative | [M−H]− | 597.18205 | 597.1825 | −0.742 | 260, 335 | * | * | * | 75566 | 2 | [58] | ||
| 60 | MS2: 609, 489, MS3: 489, 429, 309 MS4: 309 | Isoorientin 2″-O-hexoside 7-O-[6″-sinapoyl]-hexoside | [M−H]− | 977.26068 | 977.2568 | 3.929 | C44H50O25 | 263, 340 | * | 3 | [58] | ||||
| 61 | MS2: 469, 307, 161 MS3: 307, 161 | Hydroxycoumarin hexoside-pentoside | [M−H]− | 469.13626 | 469.1351 | 2.367 | C20H24O12 | * | 3 | [62] | |||||
| 62 | MS2: 307, 161, 145 MS3: 161, 145, 113 | p-Coumaroyl-caffeic acid pentoside | [M−H]− | 439.12554 | 439.1246 | 2.175 | C23H22O10 | 286sh, 315 | * | 3 | [56] | ||||
| 63 | MS2: 163, 135, 119 MS3: 119 | p-Coumaroylthreonic acid | [M−H]− | 281.0672 | 281.0667 | 1.864 | C13H14O7 | 290sh | * | 3 | [56] | ||||
| 64 | MS2: 561, 489, 459, 399, 369, 327 MS3: 441, 399, 369 MS4: 341, 313 | MS2: 563, 545, 515, 497, 485, 473, 413 MS3: 545, 515, 473, 449, 413, 365 MS4 (563-473): 455, 437, 367, 341 MS4 (563-515): 449, 431, 413 | Luteolin 6-C-hexoside-8-C-pentoside | [M−H]− | 579.1368 | 579.1355 | 2.17 | C26H28O15 | 262, 345 | * | 3 | [54] | |||
| 65 | MS2: 697, 535, 373, 329, 178 MS3: 299, 284, 269, 178, 161 | Hydroxypinoresinol di-O-hexoside | [M−H]− | 697.23658 | 697.2366 | 2.377 | C32H42O17 | 278 | * | * | 3421 | 2 | [62] | ||
| 66 | MS2: 489, 429, 327, 309, 285 MS3: 327, 298 MS4: 297, 175 | Isoorientin 6”-O-hexoside | [M−H]− | 609.1440 | 609.1450 | −1.6214 | C27H30O16 | 266, 347 | * | 75353 | 3 | [58] | |||
| 67 | MS2: 449, 303, 285 MS3: 285 | Quercetin O-deoxyhexosylhexoside | [M+H]+ | 611.1594 | 611.1607 | −2.0245 | C27H30O16 | 255, 353 | * | * | 2 | [62] | |||
| 68 | MS2: 575, 533, 503, 473, 431, 311 MS3: 413, 383, 311 | Isovitexin 6”-O-hexoside | [M−H]− | 593.15198 | 593.1520 | 1.3952 | C27H30O15 | 268, 335 | * | 3 | [53,57] | ||||
| 69 | MS2: 533, 503, 473, 413, 383, 341, 293 MS3: 312, 293 | MS2: 433, 415, 397, 367, 337, 313, 283 MS3: 415, 397, 367, 337, 313, 283367, 283 MS4:283 | Isovitexin 2”-O-glucoside | [M−H]− | 593.15094 | 593.15119 | −0.42709 | C27H30O15 | 268, 335 | * | 1 | Std; [58] | |||
| 70 | MS2: 489, 429, 369, 357, 339, 309 MS3: 429, 369, 351, 339, 309, 243 | MS2: 449, 431, 383, 353, 329, 299 MS3: 431, 413, 383, 353, 329, 299 MS4: 299 | Isoorientin 2”-O-glucoside | [M−H]− | 609.1476 | 609.1461 | 2.45 | C27H30O16 | 269, 348 | * | 17379 | 1 | Std; [58] | ||
| 71 | MS2:429, 411, 357, 327, 283 MS3: 297, 283 MS4: 269 | MS2: 431, 383, 353, 329, 299 MS3: 299 | Isoorientin | [M−H]− | 447.0939 | 447.0933 | 1.4239 | C21H20O11 | * | * | 17965 | 1 | Std | ||
| 72 | MS2: 545, 503, 473, 443, 413, 383, 353, 325 MS3 (563-353): 353, 325, 297 MS3 (563-443): 383, 353, 297, 191 | MS2: 547, 529, 511, 451, 337 MS3: 530, 499, 482, 458, 391 MS4: 512, 397 | Apigenin 6-C-glucoside-8-C-arabinoside | [M−H]− | 563.1409 | 563.1406 | 0.5600 | C26H28O14 | 265, 335 | * | * | 17965 | 1 | Std | |
| 73 | MS2: 455, 503, 473, 443, 383, 353, 337 MS3: 353, 325, 297, 203 MS4: 325, 297 | Apigenin 6-C-pentoside-8-C-hexoside | [M−H]− | 563.1416 | 563.1406 | 1.7191 | C26H28O14 | 266, 335 | * | * | * | 9047 | 2 | [62] | |
| 74 | MS2: 447, 357, 327, 285 MS3: 339, 311, 297, 285 | MS2: 431, 383, 299 MS3: 299 MS4: 183, 121 | Orientin | [M−H]− | 447.0926 | 447.0933 | −1.4845 | C21H20O11 | * | * | 7781 | 3 | [62] | ||
| 75 | MS2: 503, 473, 443, 383, 353 MS3: 365, 325, 221 | MS2: 547, 500, 457 MS3: 511, 493, 409 | Apigenin 6-C-pentoside-8-C-hexoside | [M−H]− | 563.14198 | 563.1406 | 2.4 | C26H28O14 | 266, 335 | 75589 | 3 | [62] | |||
| 76 | MS2: 593, 503, 473, 431, 311, 297, 283 MS3: 311, 283 | MS2: 577, 559, 529, 409, 475, 433, 415, 397, 367, 337, 313, 283 MS3: 559, 529, 498, 415, 397, 367, 337, 283 MS4: 175 | Isovitexin 7-O-glucoside | [M−H]− | 593.1506 | 593.1215 | −0.9416 | C27H30O15 | 265, 335 | * | * | * | 75439 | 1 | Std; [58] |
| 77 | MS2: 653, 491, 329 MS3: 315, 299 | Tricin di-O-glucoside | [M−H]− | 653.17377 | 653.1723 | 2.216 | C29H34O17 | 266, 369 | * | 2 | [58] | ||||
| 78 | MS2: 463, 301 MS3: 301, 286 | Chrysoeriol di-O-hexoside | [M+H]+ | 625.1755 | 625.1763 | −1.3123 | C28H32O16 | 246, 266, 347 | * | 3 | [57] | ||||
| 79 | MS2: 574, 533, 503, 473, 413, 383 MS3: 413, 383 MS4: 355, 312 | MS2: 577, 559, 529, 499, 463, 409, 356 MS3: 541, 529, 499, 452, 427, 377 MS4: 427, 355 | Chrysoeriol 6-C-hexoside-8-C-pentoside | [M−H]− | 593.1518 | 593.1512 | 1.0147 | C27H30O15 | 250, 348 | * | 3 | [57] | |||
| 80 | MS2: 577, 503, 457, 383, 353 MS3: 383, 353 | Apigenin 6-C-hexoside-8-C-deoxyhexoside | [M−H]− | 577.15643 | 577.1563 | 0.262 | C27H30O14 | 266, 335 | * | * | 2 | [62] | |||
| 81 | MS2: 574, 533, 503, 473, 413, 383 MS3: 413, 383 MS4: 352, 338, 312 | MS2: 577, 541, 457, 529, 511, 409, 389, 345 MS3: 559, 529, 511, 427 MS4: 511 | Chrysoeriol 6-C-pentoside-8-C-hexoside | [M−H]− | 593.15277 | 593.1512 | 2.658 | C27H30O15 | 246, 267, 346 | * | 3 | [62] | |||
| 82 | MS2: 371, 209, 175 MS3: 209, 121 | MS2: 387, 373, 369, 211, 193 MS3: 211 | Blumenol C-hexoside-glucuronide | [M−H]− | 547.23957 | 547.2396 | −0.081 | C25H39O13− | 255 | * | * | * | 2 | [58] | |
| 83 | MS2: 341, 311, 283 MS3: 283, 237, 117 | MS2: 415, 397, 367, 337, 283 MS3: 283, 271 | Isovitexin | [M−H]− | 431.0994 | 431.0984 | 2.4502 | C21H20O10 | 268, 336 | * | 18330 | 1 | Std; [58] | ||
| 84 | MS2: 503, 443, 323 MS3: 323, 308 MS4: 308 | MS2: 607, 591, 542, 463, 445, 397, 367, 343, 313, 265 MS3: 445, 427, 397, 367, 343, 313 | Isoscoparin 2”-O-glucoside | [M−H]− | 623.1611 | 623.1618 | −1.1011 | C28H32O16 | 250, 348 | * | * | 75518 | 1 | Std; [58] | |
| 85 | MS2: 491, 373, 329 MS3: 315, 175 | Tricin hexosylmalonate | [M−H]− | 577.12073 | 577.1199 | 1.45 | C26H26O15 | - | * | * | 75518 | 3 | [53] | ||
| 86 | MS2: 515, 473, 443, 413, 383, 353 MS3: 325, 297 MS4: 267 | MS2: 517, 499, 481, 469, 433, 415, 397, 308 MS3:481, 463, 445, 433, 409, 379 MS4: 463, 445, 433, 397, 373, 351, 329 | Apigenin 6-C-pentoside-8-C-pentoside | [M−H]− | 535.1458 | 535.1446 | 2.2345 | C25H26O13 | 265, 335 | * | * | 3 | [53] | ||
| 87 | MS2: 371, 341, 298 MS3: 327, 313, 298 | MS2: 455, 427, 409, 397, 367, 343, 313 MS3 (463-397): 379, 313, 301, 298 MS3 (463-445): 427, 397, 367, 313, 253 MS4 (445-367): 339, 324 MS5 (367-339): 324, 311 | Isoscoparin | [M−H]− | 461.1089 | 461.1089 | 0.0285 | C22H22O11 | * | * | 18200 | 2 | [62] | ||
| 88 | MS2: 476, 329, 314 MS3: 314, 299 | MS2: 331, MS3: 315, 287, 270 | Tricin 7-O-glucoside | [M−H]− | 491.1192 | 491.1195 | −0.6682 | C23H24O12 | 266, 368 | * | * | 75349 | 1 | Std | |
| 89 | MS2: 329, 314, 299 MS3: 314, 299 | MS2: 493, 331, 315 MS3: 315, 269 | Tricin O-hexosyldeoxyhexoside | [M−H]− | 637.1769 | 637.1774 | −0.733 | C29H34O16 | 265, 367 | * | * | * | 131777 | 3 | [58] |
| 90 | MS2: 329, 313 MS3: 314 | MS2: 493, 475, 331 MS3: 331, 315 MS4: 315, 269 | Tricin O-deoxyhexoside-O-hexoside | [M−H]− | 637.19042 | 639.19196 | −2.3961 | C29H34O16 | 266, 367 | * | 3 | [53,57] | |||
| 91 | MS2: 607, 299, 284 | MS2: 463, 301 | Chrysoeriol O-hexosyldeoxyhexoside | [M−H]− | 607.16864 [M−H]− | 607.1668 | 2.959 | C28H32O15 | 249, 250, 345 | * | * | * | 3 | [53,57] | |
| 92 | MS2: 313, 299 MS3: 299, 285, 161 MS4: 271, 203, 161 | MS2: 315, 270, 253 MS3: 299, 270, 242, 207, 153 | Tricin | [M−H]− | 329.06758 | 329.0667 | 2.747 | C17H14O7 | * | * | 59979 | 3 | [53,57] | ||
| 93 | MS2: 383, 267, 249, 193, 134, 113 | Feruloylhydroxycitric acid | [M−H]− | 383.06252 | 383.062 | 1.4026 | C16H16O11 | * | * | 176361 | 3 | [58] | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piasecka, A.; Sawikowska, A.; Jedrzejczak-Rey, N.; Piślewska-Bednarek, M.; Bednarek, P. Targeted and Untargeted Metabolomic Analyses Reveal Organ Specificity of Specialized Metabolites in the Model Grass Brachypodium distachyon. Molecules 2022, 27, 5956. https://doi.org/10.3390/molecules27185956
Piasecka A, Sawikowska A, Jedrzejczak-Rey N, Piślewska-Bednarek M, Bednarek P. Targeted and Untargeted Metabolomic Analyses Reveal Organ Specificity of Specialized Metabolites in the Model Grass Brachypodium distachyon. Molecules. 2022; 27(18):5956. https://doi.org/10.3390/molecules27185956
Chicago/Turabian StylePiasecka, Anna, Aneta Sawikowska, Nicolas Jedrzejczak-Rey, Mariola Piślewska-Bednarek, and Paweł Bednarek. 2022. "Targeted and Untargeted Metabolomic Analyses Reveal Organ Specificity of Specialized Metabolites in the Model Grass Brachypodium distachyon" Molecules 27, no. 18: 5956. https://doi.org/10.3390/molecules27185956