

UPLC-QE-Orbitrap-Based Cell Metabolomics and Network Pharmacology to Reveal the Mechanism of N-Benzylhexadecanamide Isolated from Maca (Lepidium meyenii Walp.) against Testicular Dysfunction
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Different Maca Polar Extracts Affect TM3 Cell Activity
2.2. Identification of NBH
2.3. Effects of NBH on TM3 Cell Viability and Testosterone Content
2.4. Metabolomics Analysis
2.4.1. Analyzing System Stability and QC Sample Detection
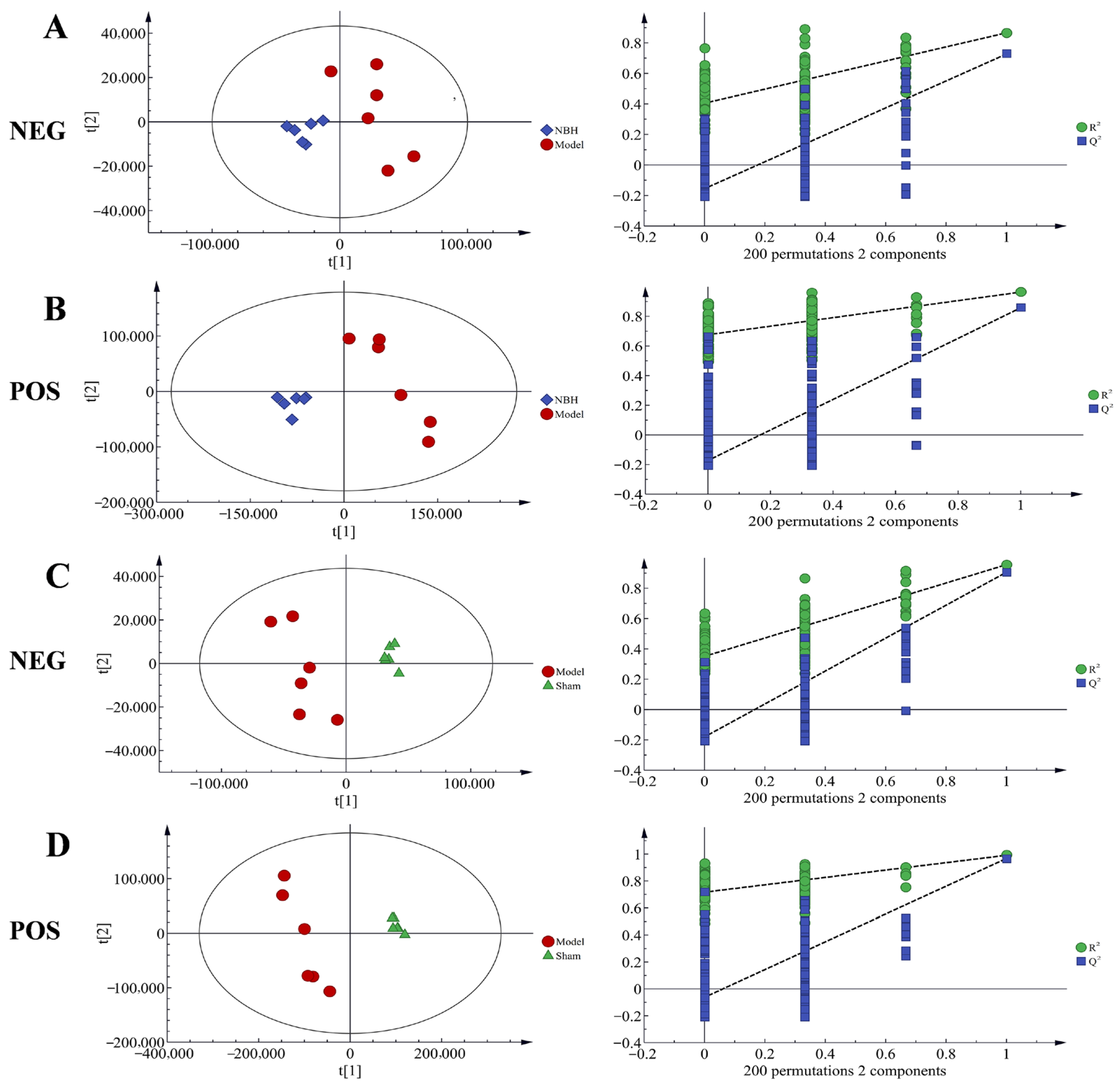
2.4.2. Statistical Analysis of Multivariate Data
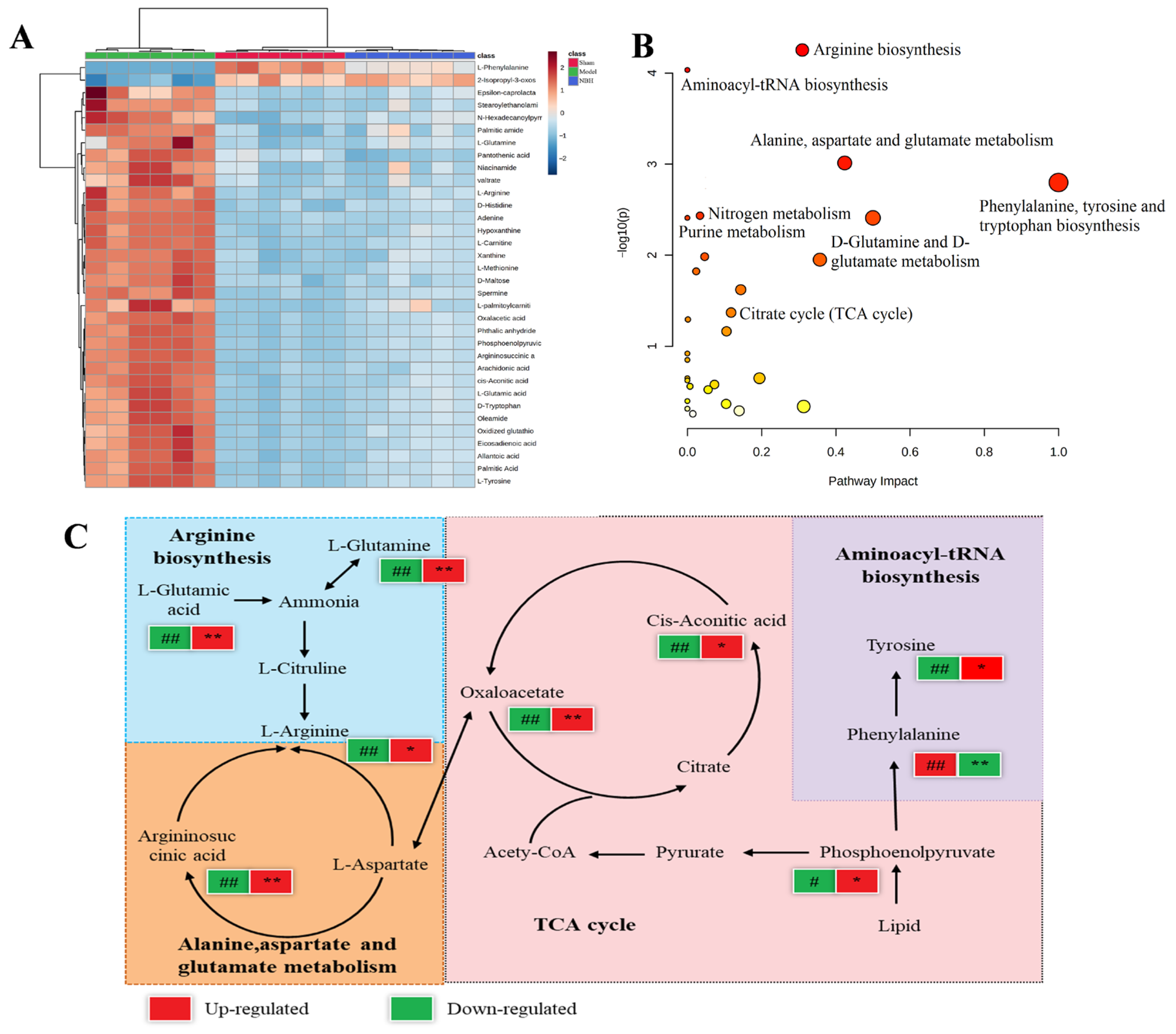
2.4.3. Biomarker Identification
2.4.4. Metabolic Pathway Analysis
2.5. Network Pharmacology Analysis
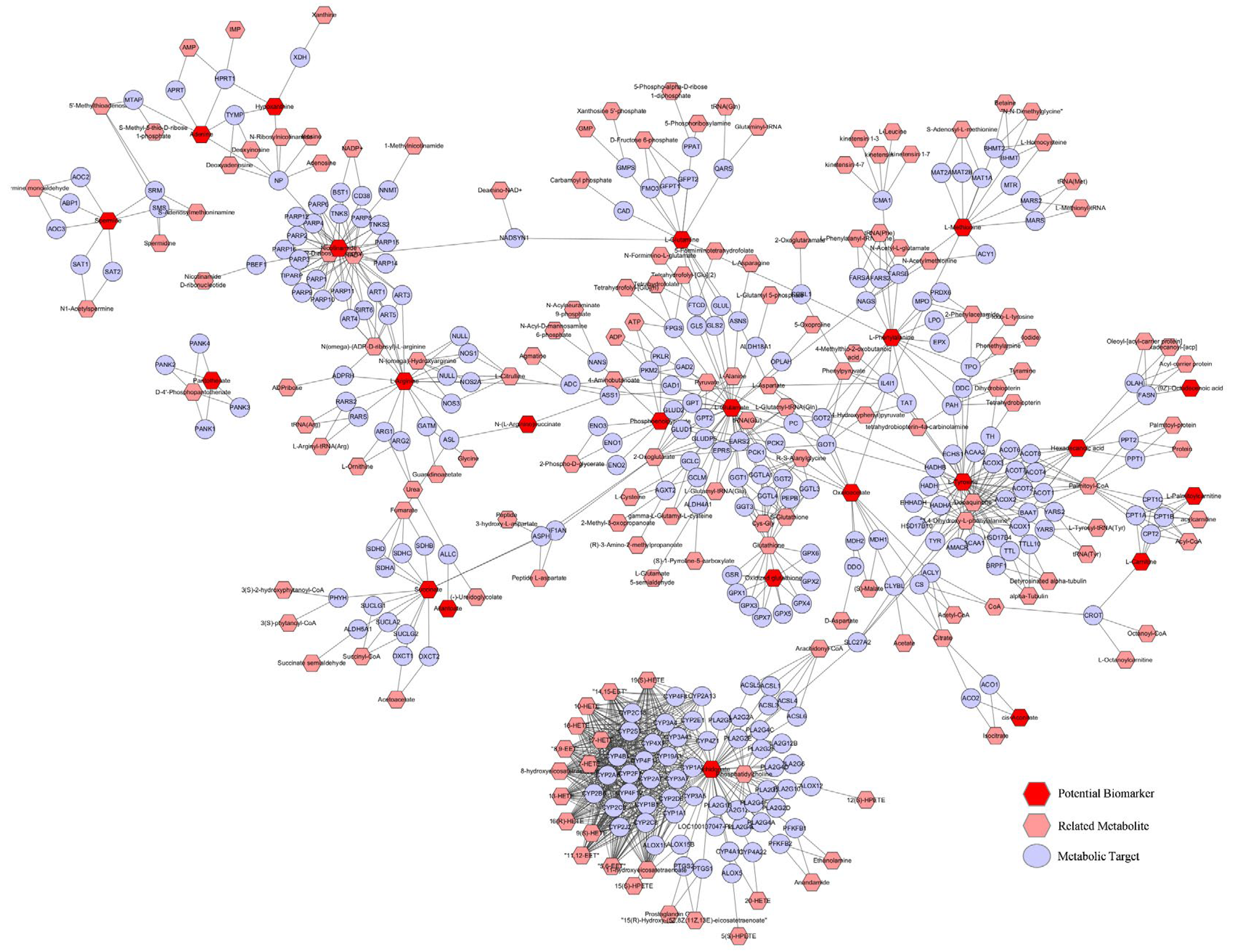
2.5.1. Analysis of Metabolic Targets Related to Potential Biomarkers
2.5.2. Targets Predicted by the Network That Are Related to the NBH Alleviation of TDF
2.5.3. PPI Network Analysis
2.5.4. Analysis of the ‘Potential Biomarker-Target-Pathway’ Network
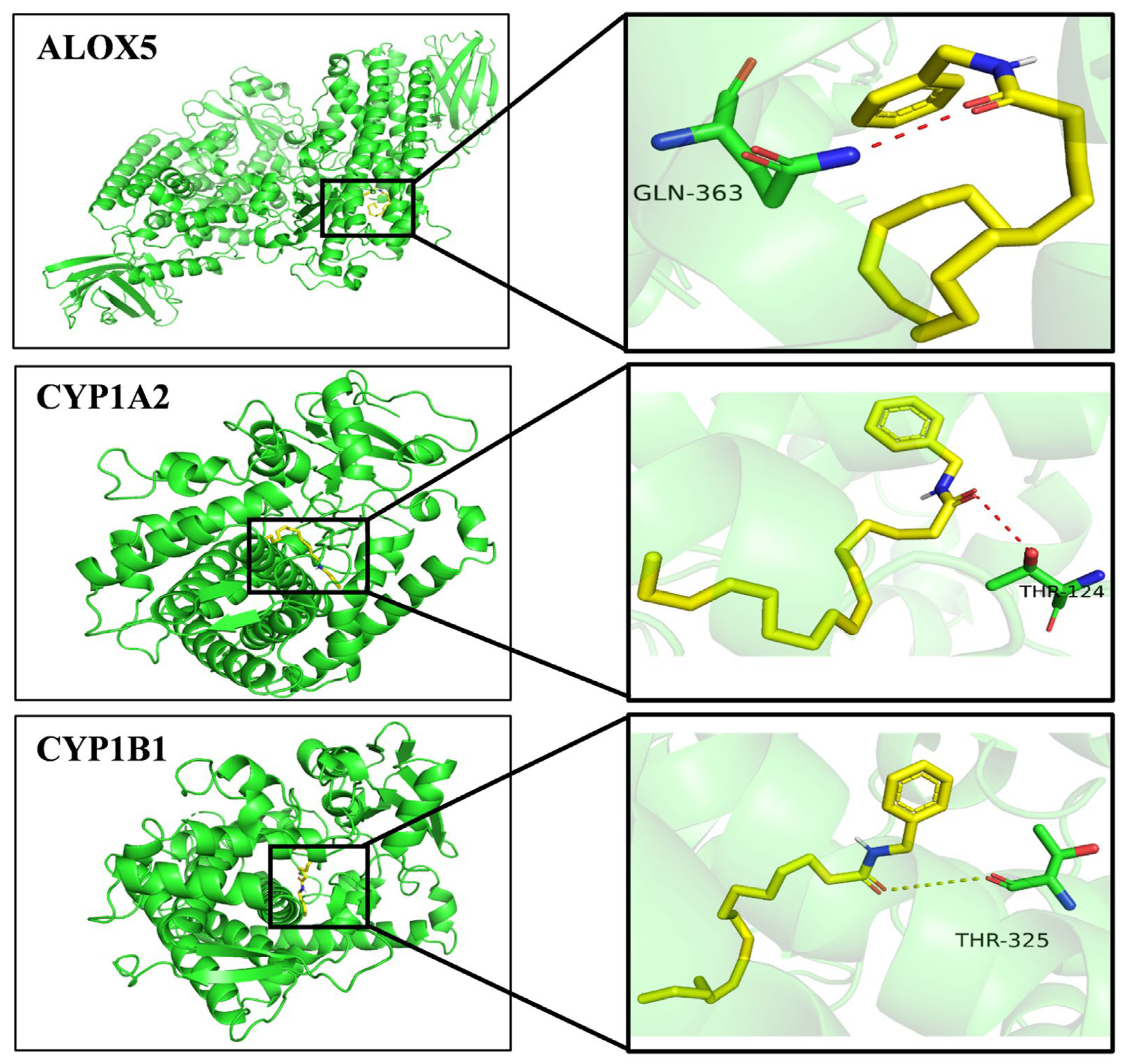
2.5.5. A Molecular Docking Results
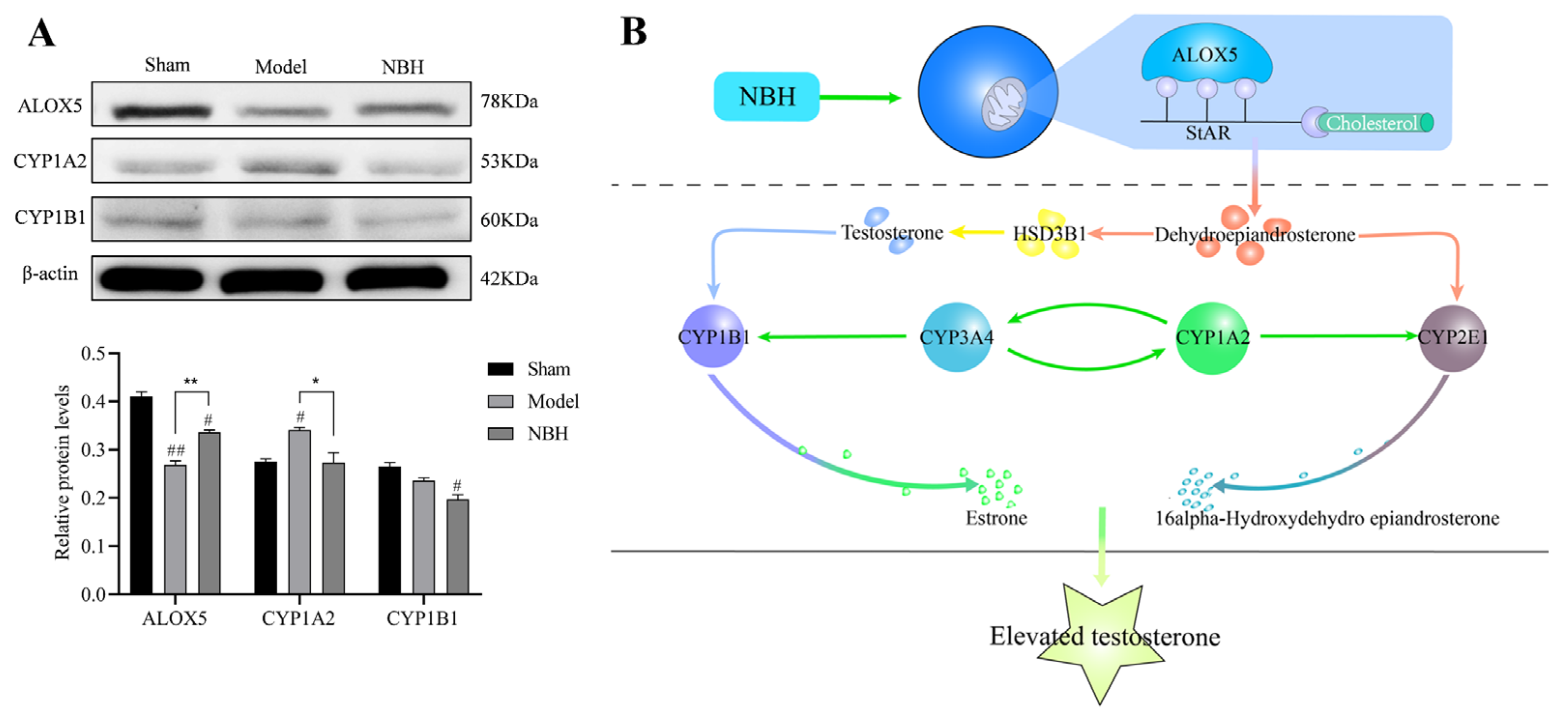
3. Discussion
4. Materials and Methods
4.1. Reagents and Material
4.2. Plant Names
4.3. Preparation of Maca Extract
4.4. Isolation and Identification of NBH
4.5. Cell Culture
4.6. Cell Viability Assay
4.7. Testosterone Levels Testing
4.8. Metabolomics Analysis
4.8.1. Preparation of Cell Samples for Sham, Model and NBH Groups
4.8.2. Metabolite Extraction
4.8.3. LC–MS/MS Analysis
4.8.4. Multivariate Statistical Analysis
Data Processing and Biomarker Selection
Hierarchical Cluster Analysis and Volcano Plots
Metabolic Pathway Analysis
4.9. Network Pharmacology Study
4.9.1. Analysis of Metabolic Targets
4.9.2. NBH Network Prediction Target Selection
4.9.3. Construction of a Network of Interactions between TDF-Related Proteins and NBH-Induced Proteins
4.9.4. GO Process and KEGG Pathway Enrichment Analysis
4.9.5. Molecular Docking and Construction of a ‘Potential Biomarker–Target-Pathway’ Network
4.9.6. Western Blot Assay
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Yao, S.; Wei, X.; Deng, W.; Wang, B.; Cai, J.; Huang, Y.; Lai, X.; Qiu, Y.; Wang, Y.; Guan, Y. Nestin-Dependent Mitochondria-ER Contacts Define Stem Leydig Cell Differentiation to Attenuate Male Reproductive Ageing. Nat. Commun. 2022, 13, 4020–4038. [Google Scholar] [CrossRef]
- Yu, S.; Han, B.; Bai, X.; Liu, S.; Xing, X.; Zhao, D.; Liu, M.; Wang, S. The Cold-Soaking Extract of Chinese Yam (Dioscorea opposita Thunb.) Protects against Erectile Dysfunction by Ameliorating Testicular Function in Hydrocortisone-Induced KDS-Yang Rats and in Oxidatively Damaged TM3 Cells. J. Ethnopharmacol. 2020, 263, 113223. [Google Scholar] [CrossRef]
- Yu, S.; Han, B.; Xing, X.; Li, Y.; Zhao, D.; Liu, M.; Wang, S. A Protein from Dioscorea polystachya (Chinese Yam) Improves Hydrocortisone-Induced Testicular Dysfunction by Alleviating Leydig Cell Injury via Upregulation of the Nrf2 Pathway. Oxidative Med. Cell. Longev. 2021, 2021, 3575016. [Google Scholar] [CrossRef]
- Basilicata, M.; di Lauro, M.; Campolattano, V.; Marrone, G.; Celotto, R.; Mitterhofer, A.P.; Bollero, P.; di Daniele, N.; Noce, A. Natural Bioactive Compounds in the Management of Oral Diseases in Nephropathic Patients. Int. J. Environ. Res. Public. Health 2022, 19, 1665. [Google Scholar] [CrossRef]
- Homayoonfal, M.; Asemi, Z.; Yousefi, B. Potential Anticancer Properties and Mechanisms of Thymoquinone in Osteosarcoma and Bone Metastasis. Cell. Mol. Biol. Lett. 2022, 27, 21–49. [Google Scholar] [CrossRef]
- Alvarado, A.T.; Navarro, C.; Pineda, M.; Villanueva, L.; Muñoz, A.M.; Bendezú, M.R.; Chávez, H.; García, J.A. Activity of Lepidium Meyenii Walp (Purple Maca) in Immunosuppressed Oryctolagus Cuniculus (Albino Rabbits). Pharmacia 2022, 69, 501–507. [Google Scholar] [CrossRef]
- Gao, X.C.; Lv, J.W.; Li, C.N.; Zhang, N.X.; Tian, L.L.; Han, X.Y.; Zhang, H.; Sun, J.M. Screening of the Active Component Promoting Leydig Cell Proliferation from Lepidium meyenii Using HPLC-ESI-MS/MS Coupled with Multivariate Statistical Analysis. Molecules 2019, 24, 2101. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Furumichi, M.; Morishima, K.; Tanabe, M. New Approach for Understanding Genome Variations in KEGG. Nucleic Acids Res. 2019, 47, 590–595. [Google Scholar] [CrossRef]
- Sarchielli, E.; Comeglio, P.; Filippi, S.; Cellai, I.; Guarnieri, G.; Marzoppi, A.; Cipriani, S.; Vignozzi, L.; Morelli, A.; Maggi, M. Neuroprotective Effects of Testosterone in the Hypothalamus of an Animal Model of Metabolic Syndrome. Int. J. Mol. Sci. 2021, 22, 1589. [Google Scholar] [CrossRef]
- Lanfranco, F.; Zitzmann, M.; Simoni, M.; Nieschlag, E. Serum Adiponectin Levels in Hypogonadal Males: Influence of Testosterone Replacement Therapy. Clin. Endocrinol. 2004, 60, 500–507. [Google Scholar] [CrossRef]
- Gonçalves, G.D.; Semprebon, S.C.; Biazi, B.I.; Mantovani, M.S.; Fernandes, G.S.A. Bisphenol a Reduces Testosterone Production in TM3 Leydig Cells Independently of Its Effects on Cell Death and Mitochondrial Membrane Potential. Reprod. Toxicol. 2018, 76, 26–34. [Google Scholar] [CrossRef]
- Komatsu, T.; Tabata, M.; Kubo-Irie, M.; Shimizu, T.; Suzuki, K.I.; Nihei, Y.; Takeda, K. The Effects of Nanoparticles on Mouse Testis Leydig Cells in Vitro. Toxicol. Vitr. 2008, 22, 1825–1831. [Google Scholar] [CrossRef]
- Shin, M.J.; Jeon, Y.K.; Kim, I.J. Testosterone and Sarcopenia. World J. Mens. Health 2018, 36, 192–198. [Google Scholar] [CrossRef]
- Bendre, S.; Murray, P.J. Clomiphene Citrate Effectively Increases Testosterone in Obese, Young, Hypogonadal Men. Reprod. Syst. Sex. Disord. 2015, 4, 155. [Google Scholar] [CrossRef]
- Zhang, N.X.; Lv, J.W.; Jin, P.; Li, J.F.; Bian, X.F.; Zhang, H.; Sun, J.M. H-1 NMR Metabonomic Investigations of N-Benzylhexadecanamide Induced Proliferation and Iestosterone Secretion of Mouse Testicular Leydig Cellst. Chem. J. Chin. Univ. Chin. 2019, 40, 1832–1839. [Google Scholar]
- Low, K.L.; Tomm, R.J.; Ma, C.; Tobiansky, D.J.; Floresco, S.B.; Soma, K.K. Effects of Aging on Testosterone and Androgen Receptors in the Mesocorticolimbic System of Male Rats. Horm. Behav. 2020, 120, 104689. [Google Scholar] [CrossRef]
- Krivoruchko, A.; Zhang, Y.; Siewers, V.; Chen, Y.; Nielsen, J. Microbial Acetyl-CoA Metabolism and Metabolic Engineering. Metab. Eng. 2015, 28, 28–42. [Google Scholar] [CrossRef]
- Seen, S. Chronic Liver Disease and Oxidative Stress—A Narrative Review. Expert. Rev. Gastroenterol. Hepatol. 2021, 15, 1021–1035. [Google Scholar] [CrossRef]
- Li, Y.-C.; Hu, X.-Q.; Xiao, L.-J.; Hu, Z.-Y.; Guo, J.; Zhang, K.-Y.; Song, X.-X.; Liu, Y.-X. An Oligonucleotide Microarray Study on Gene Expression Profile in Mouse Testis of Experimental Cryptorchidism. Front. Biosci. 2006, 11, 2465–2482. [Google Scholar] [CrossRef]
- Ji, Z.; Shen, Y.; Feng, X.; Kong, Y.; Shao, Y.; Meng, J.; Zhang, X.; Yang, G. Deregulation of Lipid Metabolism: The Critical Factors in Ovarian Cancer. Front. Oncol. 2020, 10, 593017. [Google Scholar] [CrossRef]
- Zheng, J. Energy Metabolism of Cancer: Glycolysis versus Oxidative Phosphorylation (Review). Oncol. Lett. 2012, 4, 1151–1157. [Google Scholar] [CrossRef]
- Aghazadeh, Y.; Zirkin, B.R.; Papadopoulos, V. Pharmacological Regulation of the Cholesterol Transport Machinery in Steroidogenic Cells of the Testis. Vitam. Horm. 2015, 98, 189–227. [Google Scholar] [CrossRef]
- Chen, F.; Ghosh, A.; Lin, J.; Zhang, C.; Pan, Y.; Thakur, A.; Singh, K.; Hong, H.; Tang, S. 5-Lipoxygenase Pathway and Its Downstream Cysteinyl Leukotrienes as Potential Therapeutic Targets for Alzheimer’s Disease. Brain Behav. Immun. 2020, 88, 844–855. [Google Scholar] [CrossRef]
- Cummings, M.; Massey, K.A.; Mappa, G.; Wilkinson, N.; Hutson, R.; Munot, S.; Saidi, S.; Nugent, D.; Broadhead, T.; Wright, A.I.; et al. Integrated Eicosanoid Lipidomics and Gene Expression Reveal Decreased Prostaglandin Catabolism and Increased 5-Lipoxygenase Expression in Aggressive Subtypes of Endometrial Cancer. J. Pathol. 2019, 247, 21–34. [Google Scholar] [CrossRef]
- Nebert, D.W. Aryl Hydrocarbon Receptor (AHR): “Pioneer Member” of the Basic-Helix/Loop/Helix Per-Arnt-Sim (BHLH/PAS) Family of “Sensors” of Foreign and Endogenous Signals. Prog. Lipid Res. 2017, 67, 38–57. [Google Scholar] [CrossRef]
- Sun, Q.Y.; Zhou, H.H.; Mao, X.Y. Emerging Roles of 5-Lipoxygenase Phosphorylation in Inflammation and Cell Death. Oxid. Med. Cell. Longev. 2019, 2019, 2749173. [Google Scholar] [CrossRef]
- Miller, W.L. Steroidogenesis: Unanswered Questions. Trends Endocrinol. Met. 2017, 28, 771–793. [Google Scholar] [CrossRef]
- Sumińska, M.; Bogusz-Górna, K.; Wegner, D.; Fichna, M. Non-Classic Disorder of Adrenal Steroidogenesis and Clinical Dilemmas in 21-Hydroxylase Deficiency Combined with Backdoor Androgen Pathway. Mini-Review and Case Report. Int. J. Mol. Sci. 2020, 21, 4622. [Google Scholar] [CrossRef]
- Tarabasz, D.; Szczeblewski, P.; Laskowski, T.; Płaziński, W.; Baranowska-Wójcik, E.; Szwajgier, D.; Kukula-Koch, W.; Meissner, H.O. The Distribution of Glucosinolates in Different Phenotypes of Lepidium peruvianum and Their Role as Acetyl-and Butyrylcholinesterase Inhibitors—In Silico and In Vitro Studies. Int. J. Mol. Sci. 2022, 23, 4858. [Google Scholar] [CrossRef]
- Wang, S.; Yang, X.; Wang, W.; Zhang, Y.; Li, T.; Zhao, L.; Bao, Y.; Meng, X. Interpretation of the Absorbed Constituents and Pharmacological Effect of Spica Schizonepetae Extract on Non-Small Cell Lung Cancer. PLoS ONE 2021, 16, e0248700. [Google Scholar] [CrossRef]
- Li, Z.; Lu, Y.; Guo, Y.; Cao, H.; Wang, Q.; Shui, W. Comprehensive Evaluation of Untargeted Metabolomics Data Processing Software in Feature Detection, Quantification and Discriminating Marker Selection. Anal. Chim. Acta 2018, 1029, 50–57. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Mode | Type | A | N | R2X | R2Y | Q2 | Title |
|---|---|---|---|---|---|---|---|---|
| Model 1 | Neg | PCA | 4 | 24 | 0.775 | - | - | Total |
| Model 2 | Pos | PCA | 4 | 24 | 0.696 | - | - | Total |
| Model 3 | Neg | PLS-DA | 3 | 18 | 0.888 | 0.676 | 0.301 | Total |
| Model 4 | Pos | PLS-DA | 3 | 18 | 0.536 | 0.903 | 0.315 | Total |
| Model 5 | Neg | PLS-DA | 2 | 12 | 0.86 | 0.866 | 0.727 | NBH/Model |
| Model 6 | Pos | PLS-DA | 2 | 12 | 0.5 | 0.964 | 0.858 | NBH/Model |
| Model 7 | Neg | PLS-DA | 2 | 12 | 0.881 | 0.955 | 0.905 | Model/Sham |
| Model 8 | Pos | PLS-DA | 2 | 12 | 0.598 | 0.991 | 0.967 | Model/Sham |
| No. | Annotated Metabolite | HMDB | Formula | Retention Time (min) | Molecular Weight | Adducts | Detected m/z | Model/Sham | NBH/Model |
|---|---|---|---|---|---|---|---|---|---|
| 1 | l-Arginine | HMDB0000517 | C6H14N4O2 | 0.788 | 174.113 | [M + H]+ | 175.0005 | ↓ b | ↑ c |
| 2 | d-Maltose | HMDB0000163 | C12H22O11 | 0.85 | 342.2965 | [M + H]+ | 343.1235 | ↓ b | ↑ d |
| 3 | Hypoxanthine | HMDB0000157 | C5H4N4O | 1.024 | 136.0427 | [M − H]− | 135.0352 | ↓ b | ↑ d |
| 4 | Xanthine | HMDB0000292 | C5H4N4O2 | 1.082 | 152.0354 | [M + H]+ | 153.0418 | ↓ b | ↑ c |
| 5 | L-Carnitine | HMDB0000062 | C7H16NO3 | 1.09 | 162.1139 | [M − H]− | 160.8409 | ↓ a | ↑ c |
| 6 | Oxidized glutathione | HMDB0003337 | C20H32N6O12S2 | 1.13 | 612.1575 | [M − H]− | 611.1449 | ↓ b | ↑ c |
| 7 | Niacinamide | HMDB0001406 | C6H6N2O | 1.15 | 122.0494 | [M − H]− | 121.9428 | ↓ b | ↑ d |
| 8 | Adenine | HMDB0000034 | C5H5N5 | 1.23 | 135.0558 | [M + H]+ | 136.0635 | ↓ a | ↑ d |
| 9 | Spermine | HMDB0001256 | C10H26N4 | 1.24 | 202.2176 | [M + H]+ | 203.9353 | ↓ b | ↑ c |
| 10 | l-Methionine | HMDB0000696 | C5H11NO2S | 1.27 | 149.0524 | [M + H]+ | 150.0596 | ↓ b | ↑ d |
| 11 | l-Phenylalanine | HMDB0000159 | C9H11NO2 | 2.64 | 165.0806 | [M + H]+ | 166.0879 | ↑ b | ↓ d |
| 12 | Pantothenic acid | HMDB0000210 | C9H17NO5 | 3.9 | 219.1125 | [M + H]+ | 220.1199 | ↓ b | ↑ d |
| 13 | d-Histidine | HMDB0250763 | C6H9N3O2 | 4.16 | 155.0709 | [M + H]+ | 156.992 | ↓ b | ↑ d |
| 14 | l-Glutamic acid | HMDB0000148 | C5H9NO4 | 5.19 | 147.0545 | [M + H]+ | 148.0617 | ↓ b | ↑ d |
| 15 | d-Tryptophan | HMDB0013609 | C11H12N2O2 | 5.52 | 204.0917 | [M + Na]+ | 227.9845 | ↓ b | ↑ d |
| 16 | Epsilon-caprolactam | HMDB0062769 | C6H11NO | 5.658 | 113.0825 | [M + H]+ | 114.0842 | ↓ b | ↑ d |
| 17 | Palmitic Acid | HMDB0000220 | C16H32O2 | 12.36 | 256.4241 | [M + Na]+ | 279.1611 | ↓ a | ↑ d |
| 18 | valtrate | HMDB0034493 | C22H30O8 | 15.055 | 212.1993 | [M − H]− | 211.0746 | ↓ b | ↑ d |
| 19 | Phosphoenolpyruvic acid | HMDB0000263 | C3H5O6P | 15.06 | 168.0432 | [M − H]− | 167.3693 | ↓ a | ↑ c |
| 20 | Argininosuccinic acid | HMDB0000052 | C10H18N4O6 | 15.68 | 290.2731 | [M + H]+ | 291.4325 | ↓ b | ↑ d |
| 21 | Allantoic acid | HMDB0001209 | C4H8N4O4 | 17.83 | 176.1307 | [2M − H]− | 351.2263 | ↓ b | ↑ d |
| 22 | Eicosadienoic acid | HMDB0005060 | C20H36O2 | 17.96 | 308.4986 | [M − H]− | 307.2645 | ↓ b | ↑ d |
| 23 | Phthalic anhydride | HMDB0256501 | C8H4O3 | 20.45 | 148.0174 | [M + H]+ | 149.0252 | ↓ b | ↑ d |
| 24 | Oleamide | HMDB0002117 | C18H35NO | 21.35 | 281.2741 | [M − H]− | 280.3808 | ↓ b | ↑ d |
| 25 | Oxalacetic acid | HMDB0000223 | C4H4O5 | 22.06 | 132.0824 | [M − H]− | 131.0426 | ↓ b | ↑ d |
| 26 | Arachidonic acid | HMDB0001043 | C20H32O2 | 22.29 | 304.2408 | [M − H]− | 303.2335 | ↓ b | ↑ d |
| 27 | l-Tyrosine | HMDB0000158 | C9H11NO3 | 22.57 | 181.0757 | [M − H]− | 180.9721 | ↓ b | ↑ c |
| 28 | N-Hexadecanoylpyrrolidine | HMDB0032740 | C20H39NO | 23.558 | 309.3026 | [M + H]+ | 310.8517 | ↓ b | ↑ d |
| 29 | Stearoylethanolamide | HMDB13078 | C18H37NO | 23.713 | 327.5473 | [M + H]+ | 328.9317 | ↓ a | ↑ c |
| 30 | cis-Aconitic acid | HMDB0000461 | C6H6O6 | 23.74 | 174.1082 | [M − H]− | 173.0821 | ↓ b | ↑ c |
| 31 | Palmitic amide | HMDB0012273 | C16H33NO | 23.958 | 255.4393 | [M + H]+ | 256.2741 | ↓ a | ↑ c |
| 32 | 2-Isopropyl-3-oxosuccinate | HMDB0012149 | C7H10O5 | 24.05 | 174.1513 | [2M + Na]+ | 371.3187 | ↑ b | ↓ d |
| 33 | l-palmitoylcarnitine | HMDB0240774 | C23H45NO4 | 24.44 | 399.6077 | [M − H]− | 398.3234 | ↓ b | ↑ d |
| 34 | l-Glutamine | HMDB0000641 | C5H10N2O3 | 24.52 | 146.0142 | [M − H]− | 145.6231 | ↓ b | ↑ d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, K.-Y.; Li, C.-N.; Zhang, N.-X.; Gao, X.-C.; Shen, J.-M.; Cheng, D.-D.; Wang, Y.-L.; Zhang, H.; Lv, J.-W.; Sun, J.-M. UPLC-QE-Orbitrap-Based Cell Metabolomics and Network Pharmacology to Reveal the Mechanism of N-Benzylhexadecanamide Isolated from Maca (Lepidium meyenii Walp.) against Testicular Dysfunction. Molecules 2023, 28, 4064. https://doi.org/10.3390/molecules28104064
Zhang K-Y, Li C-N, Zhang N-X, Gao X-C, Shen J-M, Cheng D-D, Wang Y-L, Zhang H, Lv J-W, Sun J-M. UPLC-QE-Orbitrap-Based Cell Metabolomics and Network Pharmacology to Reveal the Mechanism of N-Benzylhexadecanamide Isolated from Maca (Lepidium meyenii Walp.) against Testicular Dysfunction. Molecules. 2023; 28(10):4064. https://doi.org/10.3390/molecules28104064
Chicago/Turabian StyleZhang, Kai-Yue, Chun-Nan Li, Nan-Xi Zhang, Xiao-Chen Gao, Jia-Ming Shen, Duan-Duan Cheng, Yue-Long Wang, Hui Zhang, Jing-Wei Lv, and Jia-Ming Sun. 2023. "UPLC-QE-Orbitrap-Based Cell Metabolomics and Network Pharmacology to Reveal the Mechanism of N-Benzylhexadecanamide Isolated from Maca (Lepidium meyenii Walp.) against Testicular Dysfunction" Molecules 28, no. 10: 4064. https://doi.org/10.3390/molecules28104064