Compensatory Growth Induced in Zebrafish Larvae after Pre-Exposure to a Microcystis aeruginosa Natural Bloom Extract Containing Microcystins



Abstract
:1. Introduction
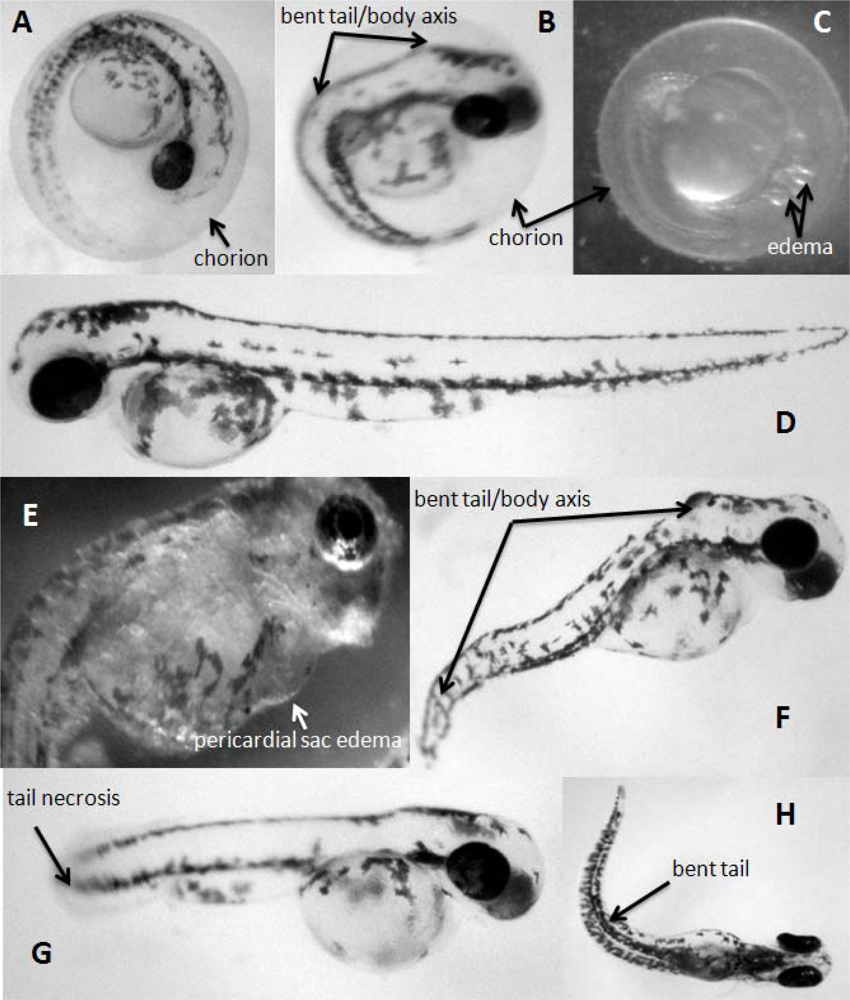
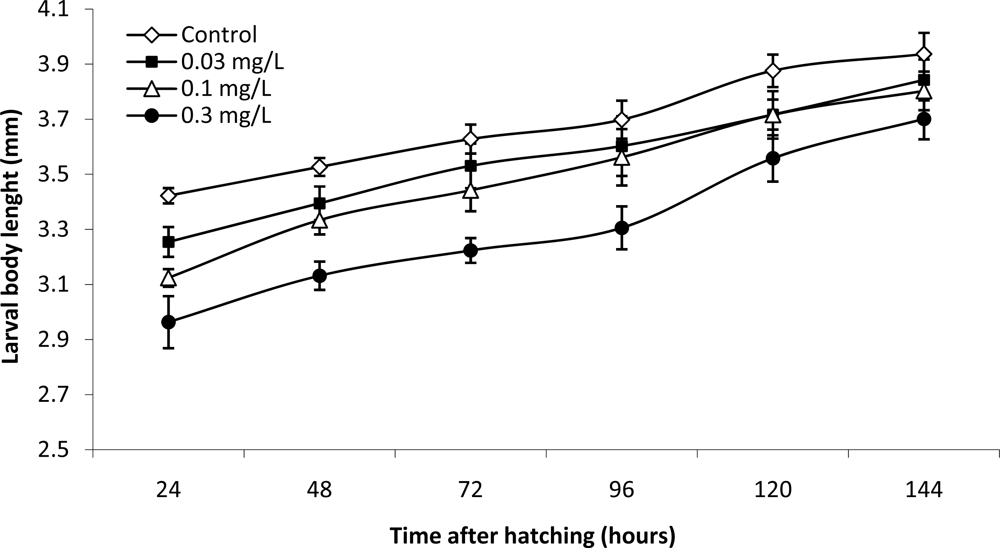
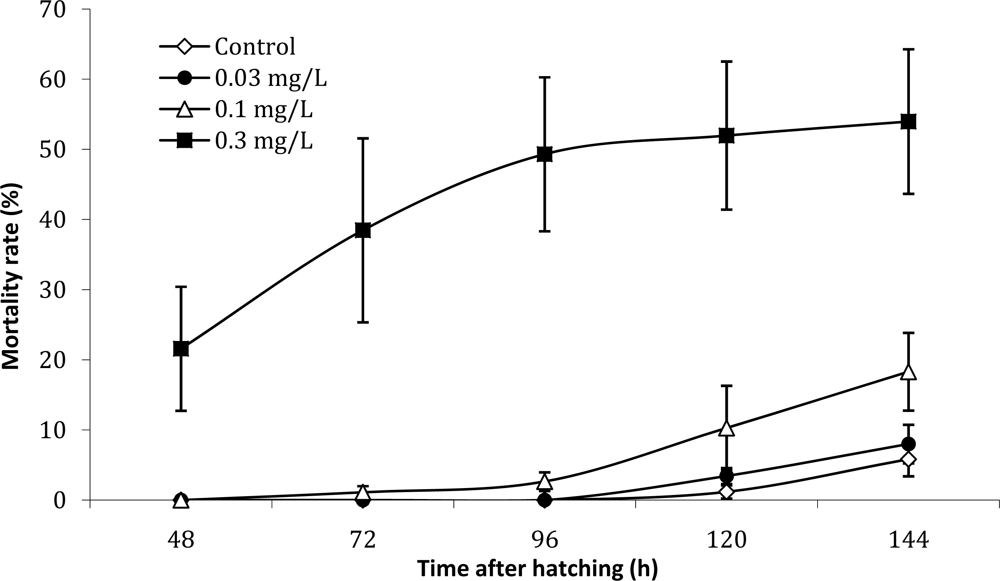
2. Results and Discussion
3. Experimental Section
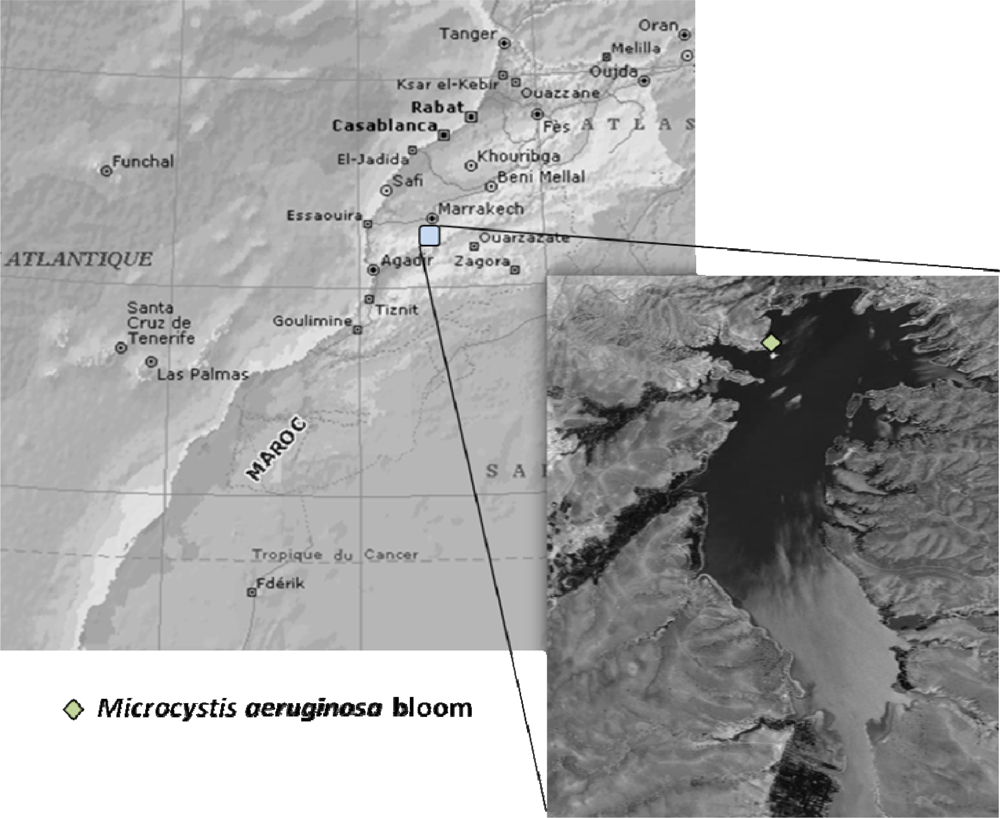
3.1. Bloom sampling
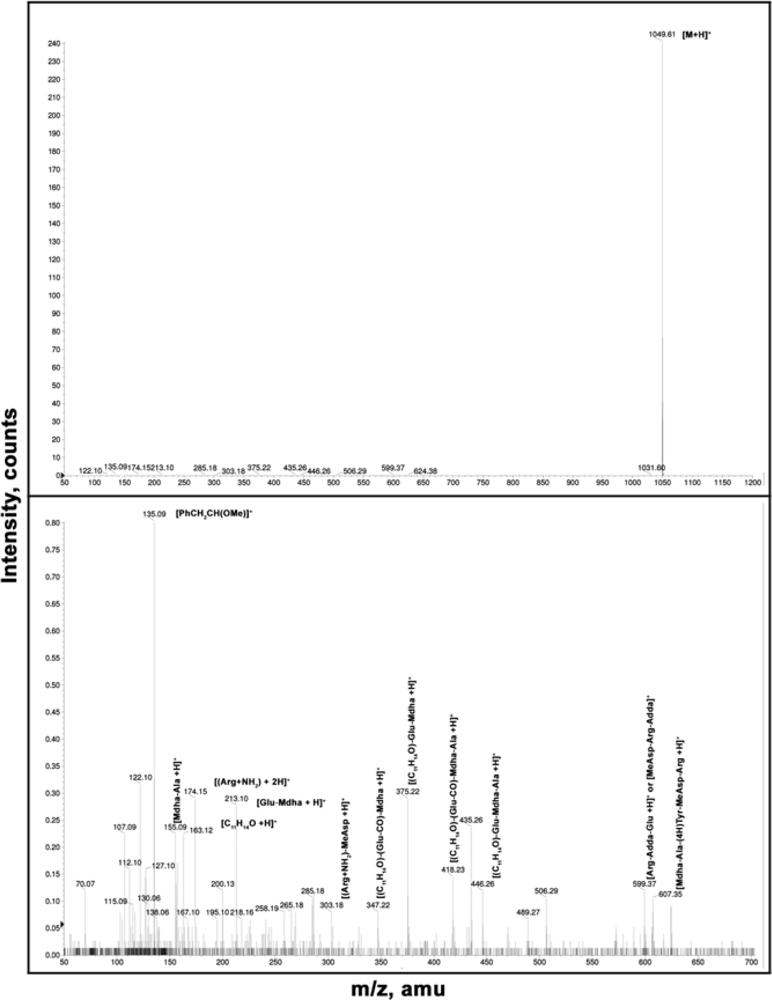
3.2. Microcystins detection and quantification
3.3. Preparation of the natural bloom extract
3.4. Exposure procedure
3.5. Calculations and statistics
4. Conclusions
Acknowledgments
References and Notes
- Chorus, I; Bartram, J. Toxic cyanobacteria in water. In A Guide to Public Health Consequences, Monitoring and Management; E and FN Spon on behalf of WHO: London, 1999. [Google Scholar]
- WHO (World Health Organization). 2003; Algae and cyanobacteria in fresh water. In Guidelines for Safe Recreational Water Environments, Volume 1, Coastal and Fresh Waters; World Health Organization: Geneva, Switzerland; pp. 136–158. [Google Scholar]
- Carmichael, WW. Cyanobacterial secondary metabolites-the cyanotoxins. J. Appl. Bacteriol 1992, 72, 445–459. [Google Scholar]
- Dawson, RM. The toxicology of microcystins. Toxicon 1998, 36, 953–962. [Google Scholar]
- Sivonen, K; Jones, G. Cyanobacterial toxins. In Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; Chorus, I, Bartram, J, Eds.; Spon, E. & F.N.: London & New-York, 1999; pp. 41–111. [Google Scholar]
- Codd, GA. Cyanobacterial toxins, the perception of water quality, and the priorisation of eutrophication control. Ecol. Eng 2000, 16, 51–60. [Google Scholar]
- Haider, S; Naithani, V; Viswanathan, PN; Kakkar, P. Cyanobacterial toxins: A growing environmental concern. Chemosphere 2003, 52, 1–21. [Google Scholar]
- Carbis, CR; Rawlin, GT; Grant, P; Mitchell, GF; Anderson, JW; McCauley, I. A study of feral carp, Cyprinus carpio L., exposed to Microcystis aeruginosa at lake Mokoan, Australia, and possible implications for fish health. J. Fish Dis 1997, 20, 81–91. [Google Scholar]
- Fischer, WJ; Dietrich, DR. Toxicity of the cyanobacterial cyclic heptapeptides toxins microcystin-LR and -RR in early life-stages of the African clawed frog (Xenopus laevis). Aquat. Toxicol 2000, 49, 189–198. [Google Scholar]
- Best, JH; Eddy, FB; Codd, GA. Effects of purified microcystin-LR and cell extracts of Microcystis strains PCC 7813 and CYA 43 on cardiac function in brown trout (Salmo trutta) alevine. Fish Physiol. Biochem 2001, 24, 171–178. [Google Scholar]
- Bury, NR; Codd, GA; Wendelaar, SE; Flik, G. Fatty acids from the cyanobacterium Microcystis aeruginosa with potent inhibitory effects on fish gills Na+/K+-ATPase activity. J. Exp. Biol 1998, 201, 81–89. [Google Scholar]
- Koop, R; Hetesa, J. Changes of Haematological indices of juvenile carp (Cyprinus carpio L.) under the influence of natural populations of cyanobacterial water blooms. Acta Vet. Brno. 2000, 69, 131–137. [Google Scholar]
- Jacquet, C; Thermes, V; de Luze, A; Puiseux-Dao, S; Bernard, C; Joly, J-S; Bourrat, F; Edery, M. Effects of microcystin-LR on development of medaka fish embryos (Oryzias latipes). Toxicon 2004, 43, 141–147. [Google Scholar]
- Mohamed, ZA; Carmichael, WW; Hussein, AA. Estimation of Microcystins in the freshwater fish Oreochromis niloticus in an Egyptian fish farm containing a Microcystis bloom. Environ. Toxicol 2003, 18, 137–141. [Google Scholar]
- Oberemm, A; Fastner, J; Steinberg, CEW. Effects of microcystin-LR and cyanobacterial crude extracts on embryolarval development of zebrafish (Danio rerio). Water Res 1997, 31, 2918–2921. [Google Scholar]
- Oberemm, A; Becker, J; Codd, GA; Steinberg, C. Effects of cyanobacterial toxins and aqueous crude extracts of cyanobacteria on the development of fish and amphibians. Environ. Toxicol 1999, 14, 77–88. [Google Scholar]
- Dvorakova, D; Dvorakova, K; Blaha, L; Marsalek, B; Knotkova, Z. Effects of cyanobacterial biomass and purified microcystins on malformations in Xenopus laevis: teratogenesis assay (FETAX). Environ. Toxicol 2002, 17, 547–555. [Google Scholar]
- Keil, C; Forchert, A; Fastner, J; Szewzyk, U; Chorus, I; Kratke, R. Toxicity and microcystin content of extracts from a Planktothrix bloom and two laboratory strains. Water Res 2002, 36, 2133–2139. [Google Scholar]
- Von Westerhagen, H. Fish Physiology; Hoar, WS, Randall, DJ, Eds.; Academic Press: San Diego, 1988; Volume 11, pp. 253–346. [Google Scholar]
- Chorus, I. Cyanotoxins: Occurrence, Causes, Consequences; Springer-Verlag: Berlin, 2001. [Google Scholar]
- Liu, Y; Song, L; Li, X; Liu, T. The toxic effects of microcystin-LR on embryo-larval and juvenile development of loach Misguruns mizolepis Gunthe. Toxicon 2002, 40, 395–399. [Google Scholar]
- Wang, PJ; Chien, MS; Wu, FJ; Chou, HN; Lee, SJ. Inhibition of embryonic development by microcystin-LR in zebrafish Danio rerio. Toxicon 2005, 45, 303–308. [Google Scholar]
- Jungmann, D; Benndorf, J. Toxicity to Daphnia of a compound extracted from laboratory and natural Microcystis spp., and the role of microcystins. Freshwater Biol 1994, 32, 13–20. [Google Scholar]
- Reinikainen, M; Kiviranta, J; Ulvi, V; Niku-Paavola, M-L. Acute toxic effects of a novel cyanobacterial toxin on the crustaceans Artemia salina and Daphnia pulex. Arch. Hydrobiol 1995, 133, 61–69. [Google Scholar]
- Feuillade, M; Jann-Para, G; Feuillade, J. Toxic compounds to Artemia from blooms and isolates of the cyanobacterium Planktothrix rubescens. Arch. Hydrobiol 1996, 138, 175–186. [Google Scholar]
- Pietsch, C; Wiegand, C; Ame, MV; Nicklisch, A; Wunderlin, D; Pflugmacher, S. The effects of a cyanobacterial crude extract on different aquatic organisms: evidence for cyanobacterial toxin modulating factors. Environ. Toxicol 2001, 16, 535–542. [Google Scholar]
- Oudra, B; Loudiki, M; Vasconcelos, V; Sabour, B; Sbiyyaa, B; Oufdou, K; Mezrioui, N. Detection and quantification of microcystins from cyanobacteria strains isolated from reservoirs and ponds in Morocco. Environ. Toxicol 2002, 17, 32–39. [Google Scholar]
- Sabour, B; Loudiki, M; Oudra, B; Vasconcelos, V; Martins, R; Oubraim, S; Fawzi. Toxicology of a Microcystis ichtyoblabe waterbloom from Lake Oued Mellah (Morocco). Environ. Toxicol 2002, 17, 24–31. [Google Scholar]
- Wiegand, C; Pflugmacher, S; Oberemm, A; Meems, N; Beattie, KA; Steinberg, CEW; Codd, GA. Uptake and effects of microcystin-LR on detoxication enzymes of early life stages of the zebra fish (Danio rerio). Environ. Toxicol 1999, 14, 89–95. [Google Scholar]
- Chernoff, N; Hunter, ES; Hall, LL; Rosen, MB; Brownie, CF; Malarkey, D; Marr, M; Herkovits, J. Lack of teratogenicity of microcystin-LR in the mouse and toad. J. Appl. Toxicol 2002, 22, 13–17. [Google Scholar]
- Ricker, WE. Computation and interpretation of biological statistics of fish populations. Bull. Fish. Res. Board Can 1975, 191, 1–382. [Google Scholar]
- Foss, A; Imsland, AK. Compensatory growth in the spotted wolfish Anarhichas minor (Olafsen) after a period of limited oxygen supply. Aquacult. Res 2002, 33, 1097–1101. [Google Scholar]
- Wilson, PN; Osbourn, DF. Compensatory growth after undernutrition in mammals and birds. Biol. Rev 1960, 35, 324–363. [Google Scholar]
- Sibly, RM; Calow, P. Physiological Ecology of Animals; Blackwell Scientific: Oxford, UK, 1986. [Google Scholar]
- Tanner, JM. Regulation of growth in size in mammals. Nature 1963, 99, 845–850. [Google Scholar]
- Monteiro, LS; Falconer, DS. Compensatory growth and sexual maturity in mice. Anim. Prod 1966, 8, 179–192. [Google Scholar]
- Björnsson, B; Sigurthorsson, G; Hemre, GI; Lie, O. Growth rate and feed conversion factor on young halibut (Hippoglossus hippoglossus L.) fed six different diets. Fiskeridirektoratets Skrifter Ernaering 1992, 5, 25–35. [Google Scholar]
- Nicieza, AG; Metcalfe, NB. Growth compensation in juvenile Atlantic salmon: Responses to depressed temperature and food availability. Ecology 1997, 78, 2385–2400. [Google Scholar]
- Maclean, A; Metcalfe, NB. Social status, access to food and compensatory growth in juvenile Atlantic salmon. J. Fish Biol 2001, 58, 1331–1346. [Google Scholar]
- Purchase, CF; Brown, JA. Stock-specific changes in growth rates, food conversion efficiencies, and energy allocation in response to temperature change in juvenile Atlantic cod. J. Fish Biol 2001, 58, 36–52. [Google Scholar]
- Speare, DJ; Arsenault, GJ. Effects of intermittent hydrogen peroxide exposure on growth and columnaris disease prevention of juvenile rainbow trout (Oncorhynchus mykiss). Can. J. Fish. Aquat. Sci 1997, 54, 2653–2658. [Google Scholar]
- Zivkov, MT. On the effect and nature of growth compensation of fish. Vest. Cesk. Spol. Zool 1982, 46, 142–160. [Google Scholar]
- Zivkov, MT. Critique of proportional hypotheses and methods for back-calculation of fish growth. Environ. Biol. Fish 1996, 46, 309–320. [Google Scholar]
- Zivkov, MT; Raikova-Petrova, GN. Growth compensation of pike-perch, Stizostedion lucioperca (L.), in the Batak and Ovcharitsa dams. Compt. Rend. Acad. Bulg. Sci 1991, 44, 45–58. [Google Scholar]
- Nicieza, AG; Reyes-Gavilan, FG; Brana, F. Differentiation in juvenile growth and bimodality patterns between northern and southern-populations of Atlantic salmon (Salmo salar L). Can. J. Zool 1994, 72, 1603–1610. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | 0.03 mg·L−1 | 0.1 mg·L−1 | 0.3 mg·L−1 | 3 mg·L−1 | |
|---|---|---|---|---|---|
| Time of beginning of hatching (h) | 73±0 | 76±0 | 77±0 | 84±0 | - |
| Hatching duration (h) | 11±0 | 22±0 | 23±0 | 38±0 | - |
| Hatching rate (%) | 66.6±1.19a | 52.2±1.82b | 47.7±3b | 45.5±3.44b | 0 |
| Malformations (%) | 2.42±1.19a | 7.11±0.73a | 9.08±1.15a | 18.05±2.44b | - |
| Larval length at hatching (mm) | 3.15±0.04a | 3.06±0.08a | 2.84±0.08b | 2.83±0.14b | - |
| Larval length after 90% of hatching (mm) | 3.30±0.04a | 3.25±0.05a | 3.12±0.03b | 3.13±0.05b | - |
| Specific growth rate (%) | 0.24±0.01a | 0.30±0.03b | 0.40±0.02c | 0.43±0.02c | - |
| Time (min) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | 3 | 33 | 40 | 50 | 55 | 60 | 62 | 65 | |
| Eluent A (%) | 65 | 65 | 55 | 55 | 35 | 0 | 0 | 65 | 65 |
| Eluent B (%) | 35 | 35 | 45 | 45 | 65 | 100 | 100 | 35 | 35 |
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/). This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ghazali, I.E.; Saqrane, S.; Carvalho, A.P.; Ouahid, Y.; Oudra, B.; Fernandez Del Campo, F.; Vasconcelos, V. Compensatory Growth Induced in Zebrafish Larvae after Pre-Exposure to a Microcystis aeruginosa Natural Bloom Extract Containing Microcystins. Int. J. Mol. Sci. 2009, 10, 133-146. https://doi.org/10.3390/ijms10010133
Ghazali IE, Saqrane S, Carvalho AP, Ouahid Y, Oudra B, Fernandez Del Campo F, Vasconcelos V. Compensatory Growth Induced in Zebrafish Larvae after Pre-Exposure to a Microcystis aeruginosa Natural Bloom Extract Containing Microcystins. International Journal of Molecular Sciences. 2009; 10(1):133-146. https://doi.org/10.3390/ijms10010133
Chicago/Turabian StyleGhazali, Issam El, Sanaa Saqrane, Antonio Paulo Carvalho, Youness Ouahid, Brahim Oudra, Francisca Fernandez Del Campo, and Vitor Vasconcelos. 2009. "Compensatory Growth Induced in Zebrafish Larvae after Pre-Exposure to a Microcystis aeruginosa Natural Bloom Extract Containing Microcystins" International Journal of Molecular Sciences 10, no. 1: 133-146. https://doi.org/10.3390/ijms10010133
APA StyleGhazali, I. E., Saqrane, S., Carvalho, A. P., Ouahid, Y., Oudra, B., Fernandez Del Campo, F., & Vasconcelos, V. (2009). Compensatory Growth Induced in Zebrafish Larvae after Pre-Exposure to a Microcystis aeruginosa Natural Bloom Extract Containing Microcystins. International Journal of Molecular Sciences, 10(1), 133-146. https://doi.org/10.3390/ijms10010133