Occurence and Bioactivities of Funicone-Related Compounds


Abstract
:1. Introduction
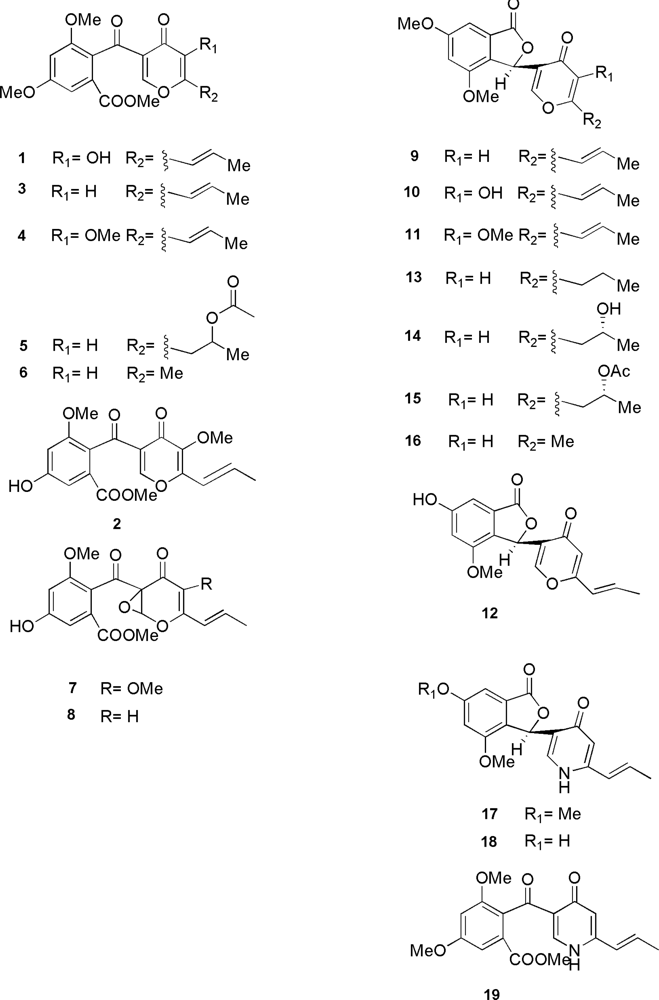
2. Molecular Structures and Producing Species
3. Methodology for Production and Extraction of Funicone-Related Compounds
4. Biological Activities of Funicone-Related Compounds
4. Conclusions
Appendix: Synthetic Media Used for Production of Funicone-Related Compounds (Composition Per Litre, Except for Semisolid Medium G)

{kind=link}
{kind=link}
| Medium A [3] | |
| Glucose | 40 g |
| Ammonium tartrate | 4.7 g |
| KH2PO4 | 1.02 g |
| MgSO4 | 0.51 g |
| KCl | 0.51 g |
| FeSO4 | 0.01 g |
| Agar | |
| Medium B [15] | |
| Glucose | 9% |
| NaNO3 | 0.2% |
| KH2PO4 | 0.1% |
| MgSO4·7H2O | 0.05% |
| KCl | 0.05% |
| FeSO4·7H2O | 0.001% |
| pH = 6.3 | |
| Medium C [8] | |
| Commercial sugar (saccharose ?) | 4% |
| Corn steep liquor | 2% |
| Medium D [7] | |
| Malt extract | 20 g |
| Glucose | 50 g |
| Peptone | 3 g |
| Medium E [10] | |
| PDB | 2.4% |
| Malt extract | 0.5% |
| Mg3(PO4)2·8H2O | 0.5% |
| Agar | 0.1% |
| pH = 6.0 | |
| Medium F [21] | |
| Glucose | 10 g |
| Peptone | 2 g |
| Yeast extract | 1 g |
| NaCl | 30 g |
| Medium F1 [22] | |
| Glucose | 10 g |
| Peptone | 2 g |
| Yeast extract | 1 g |
| NaCl | 2 g |
| pH = 7.0 | |
| Medium G [28] | |
| Grain | 7.5 g |
| Bran | 7.5 g |
| Yeast extract | 0.5 g |
| Sodium tartrate | 0.1 g |
| FeSO4·7H2O | 0.01 g |
| Sodium glutamate | 0.1 g |
| Pure corn oil | 0.1 mL |
| Water | 30 mL |
| Medium H [19] | |
| Carrot extract | ? |
| Glucose | 50 g |
| NaNO3 | 3 g |
| KH2PO4 | 1 g |
| MgSO4·7H2O | 0.5 g |
| KCl | 0.5 g |
| FeSO4·7H2O | 0.01 g |
| pH = 4.5 | |
| AD-2 [9] | |
| Glucose | 150 g |
| Glycerol | 20 g |
| Yeast extract | 4 g |
| NaNO3 | 1 g |
| Sodium glutamate | 3 g |
| Na2HPO4 | 0.5 g |
| MgSO4·7H2O | 1 g |
| Trace element solution | 1 mL |
| CaCO3 | 8 g |
| pH = 7.0 | |
| M-1-D/coconut [18] | |
| Inositol | 5 g |
| Thiamine | 0.5 g |
| Biotin | 0.5 g |
| Coconut water | 12 mL |
| Wickerham [11] | |
| Glucose | 10 g |
| Bacto-peptone | 5 g |
| Yeast extract | 3 g |
| Malt extract | 3 g |
| pH = 7.3 |
References and Notes
- Merlini, L; Nasini, G; Selva, A. The structure of funicone. A new metabolite from Penicillium funiculosum Thom. Tetrahedron 1970, 26, 2739–2749. [Google Scholar]
- Biourge, P. Les moisissures du groupe Penicillium Link. Cellule 1923, 33, 7–331. [Google Scholar]
- Locci, R; Merlini, L; Nasini, G; Rogers Locci, J. Mitorubrinic acid and related compounds from a strain of Penicillium funiculosum Thom. Giorn. Microbiol 1967, 15, 93–102. [Google Scholar]
- Pitt, JI. The genus Penicillium and its teleomorphic states Eupenicillium and Talaromyces; Academic Press: London-New York, 1979; pp. 1–634. [Google Scholar]
- Komai, S; Hosoe, T; Itabashi, T; Nozawa, K; Okada, K; de Campos Takaki, GM; Chikamori, M; Yaguchi, T; Fukushima, K; Miyaji, M; Kawai, K. A new funicone derivative isolated from Talaromyces flavus. Mycotoxins 2004, 54, 15–19. [Google Scholar]
- Komai, S; Hosoe, T; Itabashi, T; Nozawa, K; Yaguchi, T; Fukushima, K; Kawai, K. New vermistatin derivatives isolated from Penicillium simplicissimum. Heterocycles 2005, 65, 2771–2776. [Google Scholar]
- Kimura, Y; Yoshinari, T; Shimada, A; Hamasaki, T. Isofunicone, a pollen growth inhibitor produced by the fungus, Penicillium sp. Phytochemistry 1995, 40, 629–631. [Google Scholar]
- Sassa, T; Nukina, M; Suzuki, Y. Deoxyfunicone, a new γ-pyrone metabolite from a resorcylide producing fungus (Penicillium sp.). Agr. Biol. Chem 1991, 55, 2415–2416. [Google Scholar]
- Singh, SB; Jayasuriya, H; Dewey, R; Polishook, JD; Dombrowski, AW; Zink, DL; Guan, Z; Collado, J; Platas, G; Pelaez, F; Felock, PJ; Hazuda, DJ. Isolation, structure, and HIV-1-integrase inhibitory activity of structurally diverse fungal metabolites. J. Ind. Microbiol. Biotechnol 2003, 30, 721–731. [Google Scholar]
- Arai, M; Comoda, H; Okuda, T; Wang, H; Tabata, N; Masuma, R; Yamaguchi, Y; Omura, S. Funicone-related compounds, potentiators of antifungal miconazole activity, produced by Talaromyces flavus FKI-0076. J. Antib 2002, 55, 172–180. [Google Scholar]
- Rusman, Y. Isolation of new secondary metabolites from sponge-associated and plant-derived endophytic fungi. inaugural-dissertation zur erlangung des doktorgrades der mathematischnaturwissenschaftlichen fakultät der heinrich-heine-universität düsseldorf, 2006; pp. 1–303.
- de Stefano, S; Nicoletti, R; Milone, A; Zambardino, S. 3-O-Methylfunicone, a fungitoxic metabolite produced by the fungus Penicillium pinophilum. Phytochemistry 1999, 52, 1399–1401. [Google Scholar]
- Nozawa, K; Nakajima, S; Kawai, KI; Udagawa, SI. A γ-pyrone derivative, rapicone, from Ramichloridium apiculatum. Phytochemistry 1992, 31, 4177–4179. [Google Scholar]
- De Stefano, S; Nicoletti, R; Zambardino, S; Milone, A. Structure elucidation of a novel funicone-like compound produced by Penicillium pinophilum. Nat. Prod. Lett 2002, 16, 207–211. [Google Scholar]
- Fuska, J; Fuskova, A; Nemec, P. Vermistatin, antibiotikum s cytotoxickými účinkami, produkované Penicillium vermiculatum. Biologia 1979, 34, 735–739. [Google Scholar]
- Fuska, J; Uhrin, D; Proksa, B; Voticky, Z; Ruppeldt, J. The structure of vermistatin, a new metabolite from Penicillium vermiculatum. J. Antib 1986, 39, 1605–1608. [Google Scholar]
- Massias, M; Molho, L; Rebuffat, S; Cesario, M; Guilhen, J; Pascard, C; Bodo, B. Vermiculinol and vermiculidiol, macrodiolides from the fungus Penicillium vermiculatum. Phytochemistry 1989, 28, 1491–1494. [Google Scholar]
- Upadhyay, RK; Strobel, GA; Coval, SJ; Clardy, J. Fijiensin, the first phytotoxin from Mycosphaerella fijiensis, the causative agent of black sigatoka disease. Experientia 1990, 46, 982–984. [Google Scholar]
- Murtaza, N; Husain, SA; Sarfaraz, TB; Sultana, N; Faizi, S. Isolation and identification of vermistatin, ergosterol, stearic acid and mannitol, metabolic products of Penicillium verruculosum. Planta Med 1997, 63, 191. [Google Scholar]
- Dethoup, T; Manoch, L; Kijjoa, A; Pinto, M; Gales, L; Damas, AM; Silva, AMS; Eaton, G; Herz, W. Merodrimanes and other constituents from Talaromyces thailandiasis. J. Nat. Prod 2007, 70, 1200–1202. [Google Scholar]
- Xia, X; Huang, H; She, Z; Cai, J; Lan, L; Zhang, J; Fu, L; Vrijmoed, LLP; Lin, Y. Structural and biological properties of vermistatin and two new vermistatin derivatives isolated from the marine-mangrove endophytic fungus Guignardia sp. No. 4382. Helv. Chim. Acta 2007, 90, 1925–1931. [Google Scholar]
- Xia, X; Liu, F; She, Z; Yang, L; Li, M; Vrijmoed, LLP; Lin, Y. 1H and 13C NMR assignments for 6-demethylvermistatin and two penicillide derivatives from the mangrove fungus Guignardia sp. (No. 4382) from the South China Sea. Mag. Reson. Chem 2008, 46, 693–696. [Google Scholar]
- Proksa, B; Adamcova, J; Fuska, J. Detection and assay of secondary metabolites of Penicillium vermiculatum DANG. J. Chromatogr 1994, 665, 185–190. [Google Scholar]
- de Stefano, S; Nicoletti, R. Isolamento dell’acido 2-metil-4-idrossibenzoico come estere metilico dal fungo Penicillium pinophilum. Suo possibile ruolo nella biogenesi dei composti a scheletro funiconico. Il Tabacco 1999, 7, 25–28. [Google Scholar]
- Kamino, T; Kuramochi, K; Kobayashi, S. A concise approach to 5-substituted-4-pyrones from kojic acid. Tetr. Lett 2003, 44, 7349–7351. [Google Scholar]
- Parrish, FW; Wiley, J; Simmons, G; Long, L, Jr. Production of aflatoxins and kojic acid by species of Aspergillus and Penicillium. Appl. Microbiol 1966, 14, 139. [Google Scholar]
- Bentley, R. From miso, sak and shoyu to cosmetics: A century of science for kojic acid. Nat. Prod. Rep 2006, 23, 1046–1062. [Google Scholar]
- Ge, HM; Shen, Y; Zhu, CH; Tan, SH; Ding, H; Song, YC; Tan, RX. Penicidones A-C, three cytotoxic alkaloidal metabolites of an endophytic Penicillium sp. Phytochemistry 2008, 69, 571–576. [Google Scholar]
- Fukuda, T; Yamaguchi, Y; Masuma, R; Tomoda, H; Omura, S. Citridones, new potentiators of antifungal miconazole activity, produced by Penicillium sp. FKI-1938. I. Taxonomy, fermentation, isolation and biological properties. J. Antib 2005, 58, 309–314. [Google Scholar]
- Adamcová, J; Proksa, B; Fuska, J. Regulation of biosynthesis of vermiculin and vermistatin in Penicillium vermiculatum. Folia Microbiol 1992, 37, 50–52. [Google Scholar]
- Proksa, B; Sturdíková, M; Mojumdar, SC; Fuska, J. Production of (–)-mitorubrinic acid by Penicillium vermiculatum. Folia Microbiol 1997, 42, 133–135. [Google Scholar]
- Singh, SB; Pelaez, F; Hazuda, DJ; Lingham, RB. Discovery of natural product inhibitors of HIV-1 integrase at Merck. Drugs Fut 2005, 30, 277–299. [Google Scholar]
- Nicoletti, R; De Stefano, M; de Stefano, S; Trincone, A; Marziano, F. Antagonism against Rhizoctonia solani and fungitoxic metabolite production by some Penicillium isolates. Mycopathologia 2004, 158, 465–474. [Google Scholar]
- Nicoletti, R; Carella, A. Composti a scheletro funiconico prodotti da specie di Penicillium. Petria 2004, 14, 1–11. [Google Scholar]
- Stammati, A; Nicoletti, R; De Stefano, S; Zampaglioni, F; Zucco, F. Cytostatic properties of a novel compound derived from Penicillium pinophilum: an in vitro study. Altern. Lab. Anim 2002, 30, 1–7. [Google Scholar]
- Buommino, E; Nicoletti, R; Gaeta, GM; Orlando, M; Ciavatta, ML; Baroni, A; Tufano, MA. 3-O-Methylfunicone induces apoptosis and hsp70 activation in HeLa cells. Cell Prolif 2004, 37, 413–426. [Google Scholar]
- Nicoletti, R; Buommino, E; De Filippis, A; Lopez-Gresa, MP; Manzo, E; Carella, A; Petrazzuolo, M; Tufano, MA. Bioprospecting for antagonistic Penicillium strains as a resource of new antitumor compounds. World J. Microbiol. Biotechnol 2008, 24, 189–195. [Google Scholar]
- Baroni, A; De Luca, A; De Filippis, A; Petrazzuolo, M; Manente, L; Nicoletti, R; Tufano, MA; Buommino, E. 3-O-Methylfunicone, a metabolite from Penicillium pinophilum, inhibits proliferation of human melanoma cells by causing G2/M arrest and inducing apoptosis. Cell Prolif 2009, 42. in press. [Google Scholar]
- Buommino, E; Boccellino, M; De Filippis, A; Petrazzuolo, M; Cozza, V; Nicoletti, R; Ciavatta, ML; Quagliuolo, L; Tufano, MA. 3-O-methylfunicone produced by Penicillium pinophilum affects cell motility of breast cancer cells, downregulating αvβ5 integrin and inhibiting metalloproteinase-9 secretion. Mol. Carcinog 2007, 46, 930–940. [Google Scholar]
- Edwards, ML; Stemerick, DM; Sunkara, PS. Chalcones: a new class of antimitotic agents. J. Med. Chem 1990, 33, 1948–1954. [Google Scholar]
- Zamilpa, A; Herrera-Ruiz, M; del Olmo, E; López-Pérez, JL; Tortoriello, J; San Feliciano, A. Anxiolytic effects of benzalphthalides. Bioorg. Med. Chem. Lett 2005, 15, 3483–3486. [Google Scholar]
- Proksa, B; Uhrínová, S; Adamcová, J; Fuska, J. Hydrogenation of vermistatin. Monatsh. für Chem 1992, 123, 251–256. [Google Scholar]
- Balaž, Š; Uher, M; Brtko, J; Veverka, M; Bransova, J; Dobias, J; Podova, M; Buchvald, J. Relationship between antifungal activity and hydrophobicity of kojic acid derivatives. Folia Microbiol 1993, 38, 387–391. [Google Scholar]
- Frisvad, JC; Filtenborg, O; Samson, RA; Stolk, AC. Chemotaxonomy of the genus Talaromyces. Antonie Leeuw 1990, 57, 179–189. [Google Scholar]
- Van Reenen-Hoekstra, ES; Frisvad, JC; Samson, RA; Stolk, AC. The Penicillium funiculosum complex – well defined species and problematic taxa. In Modern concepts in Penicillium and Aspergillus classification; Samson, RA, Pitt, JI, Eds.; Plenum Press: New York, USA, 1990; pp. 173–191. [Google Scholar]
- Frisvad, JC; Smedsgaard, J; Larsen, TO; Samson, RA. Mycotoxins, drugs and other extrolites produced by species in Penicillium subgenus Penicillium. Stud. Mycol 2004, 49, 201–242. [Google Scholar]


| Compound | Formula | Mol. weight | Melting point (°C) | UV absorbance (nm) |
|---|---|---|---|---|
| Acetoxy-dihydrovermistatin | C20H20O8 | 388.1160 | undetermined | 210, 246, 305 |
| Actofunicone | C21H22O9 | 418.1264 | - | 208, 246, 316 |
| 6-Demethylvermistatin | C17H14O6 | 314.0787 | 195–196 | |
| Deoxyfunicone | C19H18O7 | 358.1053 | 127.5–128.5 | 220, 249, 314 |
| 14,15-Dihydrovermistatin | C18H18O6 | 330.1103 | 143–145 | 210, 245, 303 |
| 9,14-Epoxy-11-deoxyfunicone | C19H18O8 | 374.0975 | 166–167 | 203, 282 |
| Funicone | C19H18O8 | 374.1004 | 176–178 | 245, 310, 368 |
| Hydroxy-dihydrovermistatin | C18H18O7 | 346.1064 | 184–185 | 210, 248, 306 |
| Hydroxyvermistatin | C18H16O7 | 344.0887 | 226–227 | |
| Isofunicone | C19H18O8 | 374.1004 | 215–218 | 249, 285, 293 |
| Methoxyvermistatin | C19H18O7 | 358.1053 | 198–199 | |
| 3-O-Methyl-5,6-epoxyfunicone | C20H20O9 | 404.1107 | undetermined | 210, 312 |
| 3-O-Methylfunicone | C20H20O8 | 388.1160 | 185–187 | 249, 293 |
| Penicidone A | C18H18NO5 | 328.1179 | undetermined | 206, 237 |
| Penicidone B | C17H16NO5 | 314.1023 | undetermined | 209, 239 |
| Penicidone C | C19H20NO6 | 358.1285 | undetermined | 215, 253 |
| Penisimplicissin | C16H14O6 | 302.0790 | 185–186 | 210, 247, 305 |
| Rapicone | C17H16O7 | 332.0896 | 162–163 | 244, 314 |
| Vermistatin | C18H16O6 | 328.0950 | 213–214 | 210, 262, 302 |
| Compound | Species | Reference | Substrate * | Conditions of growth | Extraction | Purification |
|---|---|---|---|---|---|---|
| Acetoxy-dihydrovermistatin | P. simplicissimum | 6 | Moist rice | 21 d, 25°C | CH2Cl2-MeOH; EtOAc | CC; LPLC; HPLC |
| Actofunicone | T. flavus | 10 | Medium E | 5 d, 27°C, AG | Me2CO; EtOAc | MPLC; HPLC |
| 6-Demethylvermistatin | Guignardia sp. | 22 | Medium F1 | 25 d, 28°C | EtOAc | CC |
| Deoxyfunicone | Penicillium sp. | 8 | Medium C | 6 d, 25°C, AG | EtOAc | CC |
| Penicillium sp. | 9 | AD2 | 21 d, 22°C, AG | MeEtCO | CC; HPLC; TLC | |
| T. flavus | 10 | Medium E | 5 d, 27°C, AG | Me2CO; EtOAc | MPLC; HPLC | |
| 14,15-Dihydroverm-istatin | P. simplicissimum | 6 | Moist rice | 21 d, 25°C | CH2Cl2-MeOH; EtOAc | CC; LPLC; HPLC |
| E. euglaucum | 11 | Wickerham | 21 d, RT | EtOAc | CC | |
| 9,14-Epoxy-11-deoxy-funicone | T. flavus | 5 | Moist barley grains | 21 d, 25°C | CH2Cl2-MeOH; EtOAc | CC; LPLC |
| Funicone | P. funiculosum (?) | 3 | Medium A | 4 d | Et2O | TLC |
| T. flavus | 5 | Moist barley grains | 21 d, 25°C | CH2Cl2-MeOH; EtOAc | CC; LPLC | |
| P. simplicissimum | 6 | Moist rice | 21 d, 25°C | CH2Cl2-MeOH; EtOAc | CC; LPLC; HPLC | |
| Hydroxy-dihydrovermistatin | P. simplicissimum | 6 | Moist rice | 21 d, 25°C | CH2Cl2-MeOH; EtOAc | CC; LPLC; HPLC |
| T. thailandiasis | 20 | Moist rice | 30 d, 28°C | EtOAc; CHCl3 | CC | |
| Hydroxyvermistatin | Guignardia sp. | 21 | Medium F | 30 d, RT | EtOAc | CC; TLC |
| Isofunicone | Penicillium sp. | 7 | Medium D | 21 d, 24°C, ST | EtOAc | CC |
| Methoxyvermistatin | P. simplicissimum | 6 | Moist rice | 21 d, 25°C | CH2Cl2-MeOH; EtOAc | CC; LPLC; HPLC |
| Guignardia sp. | 21 | Medium F | 30 d, RT | EtOAc | CC | |
| 3-O-Methyl-5,6-epoxyfunicone | P. pinophilum | 14 | PDB | 21 d, 25°C, ST | CH3Cl; Me2CO-MeOH | TLC |
| 3-O-Methylfunicone | P. pinophilum | 12 | PDB | 21 d, 25°C, ST | CH3Cl; Me2CO-MeOH | TLC |
| Penicidones | Penicillium sp. | 28 | Medium G | 20 d, 28°C | MeOH; EtOAc | CC |
| Penisimplicissin | P. simplicissimum | 6 | Moist rice | 21 d, 25°C | CH2Cl2-MeOH; EtOAc | CC; LPLC; HPLC |
| T. thailandiasis | 20 | Moist rice | 30 d, 28°C | EtOAc; CHCl3 | CC | |
| Rapicone | R. apiculatum | 13 | PDB | 21 d, 28°C, ST | CH2Cl2 | CC; LPLC |
| Vermistatin | T. flavus | 15 | Medium B | 5–6 d, 28°C, AG | CH2Cl2 | TLC |
| 17 | Czapek-Dox broth | 30 d, 27°C, AG | EtOAc | CC; TLC | ||
| 10 | Medium E | 5 d, 27°C, AG | Me2CO; EtOAc | MPLC; HPLC | ||
| 5 | Moist barley grains | 21 d, 25°C | CH2Cl2-MeOH; EtOAc | CC; LPLC | ||
| M. fijiensis | 18 | M-1-D/coconut | 28 d, 26°C, AG, 12h | MeOH; EtOAc | TLC; HPLC | |
| P. verruculosum | 19 | Medium H | 17 d, 24°C | EtOAc | CC | |
| P. simplicissimum | 6 | Moist rice | 21 d, 25°C | CH2Cl2-MeOH; EtOAc | CC; LPLC; HPLC | |
| E. euglaucum | 11 | Wickerham | 21 d, RT | EtOAc | CC | |
| T. thailandiasis | 20 | Moist rice | 30 d, 28°C | EtOAc; CHCl3 | CC | |
| Guignardia sp. | 21 | Medium F | 30 d, RT | EtOAc | CC; TLC |
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nicoletti, R.; Manzo, E.; Ciavatta, M.L. Occurence and Bioactivities of Funicone-Related Compounds. Int. J. Mol. Sci. 2009, 10, 1430-1444. https://doi.org/10.3390/ijms10041430
Nicoletti R, Manzo E, Ciavatta ML. Occurence and Bioactivities of Funicone-Related Compounds. International Journal of Molecular Sciences. 2009; 10(4):1430-1444. https://doi.org/10.3390/ijms10041430
Chicago/Turabian StyleNicoletti, Rosario, Emiliano Manzo, and Maria Letizia Ciavatta. 2009. "Occurence and Bioactivities of Funicone-Related Compounds" International Journal of Molecular Sciences 10, no. 4: 1430-1444. https://doi.org/10.3390/ijms10041430
APA StyleNicoletti, R., Manzo, E., & Ciavatta, M. L. (2009). Occurence and Bioactivities of Funicone-Related Compounds. International Journal of Molecular Sciences, 10(4), 1430-1444. https://doi.org/10.3390/ijms10041430