The Individual and Combined Effects of Deoxynivalenol and Aflatoxin B1 on Primary Hepatocytes of Cyprinus Carpio

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
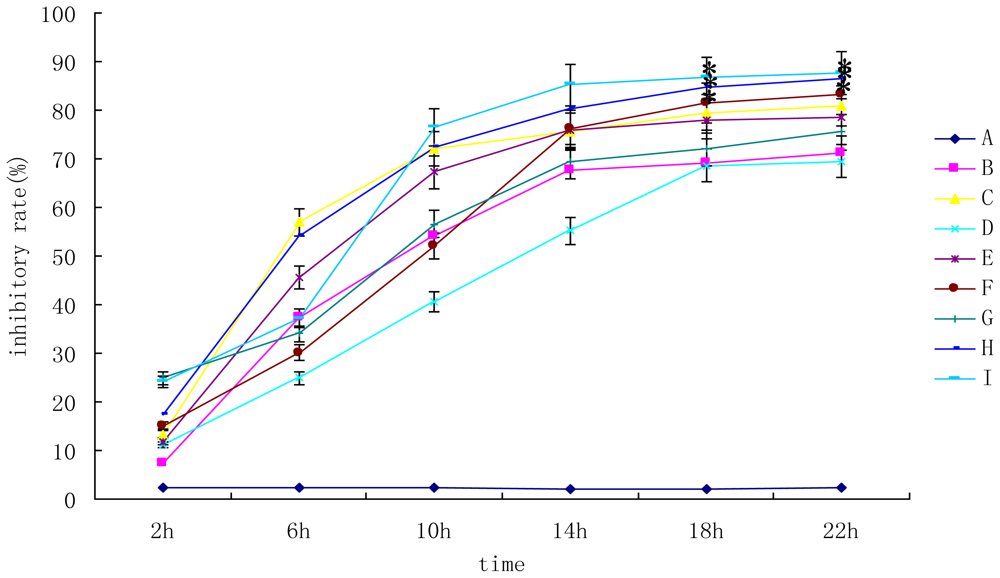
2.1. Individual and Combined Toxicity of DON and AFB1 on Primary Hepatocytes
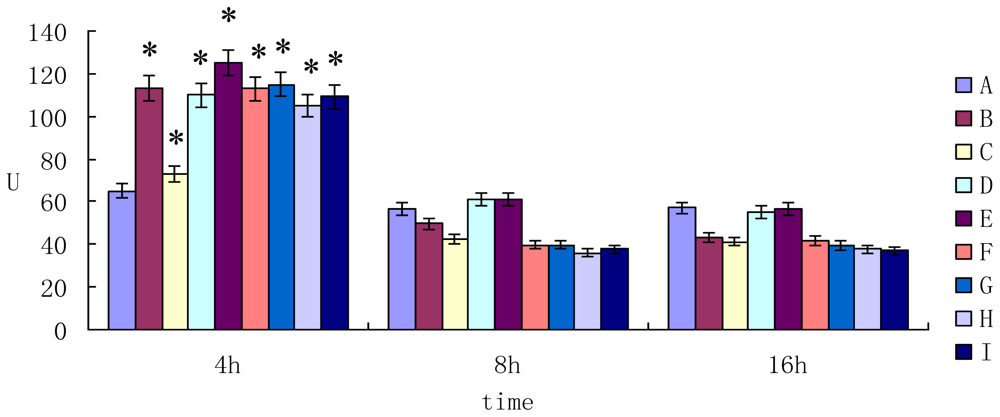
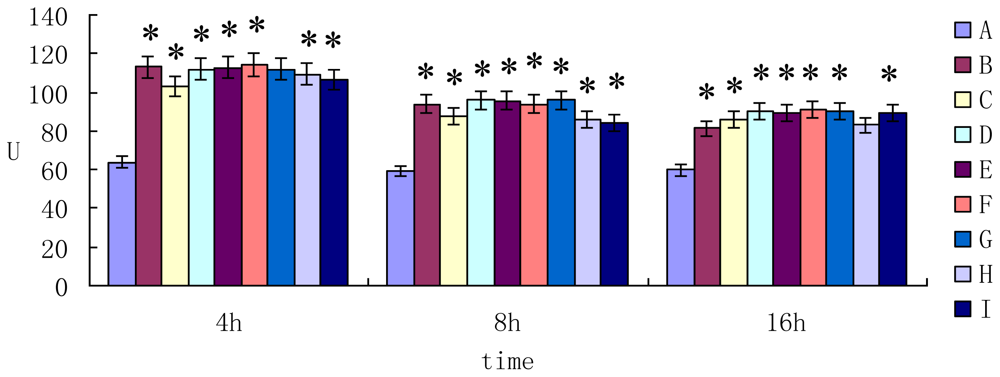
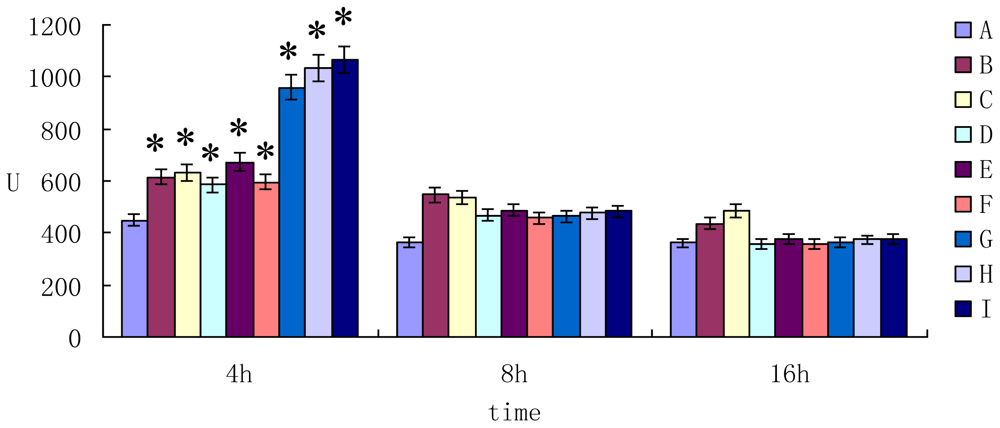
2.2. The Enzyme Activity of AST ALT and LDH in Cell Supernatant
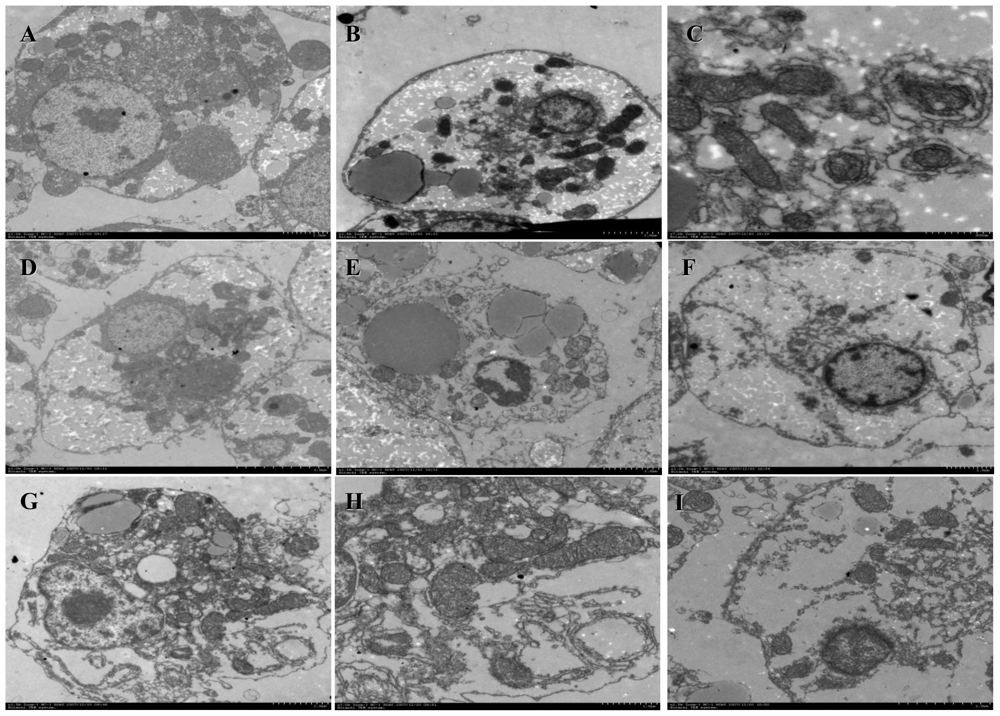
2.3. Morphological Observation on Cells
3. Experimental Section
3.1. Mycotoxin
3.2. Cell Cultures and Treatments
3.3. Cytotoxicity Assay by MTT Test
3.4. The Enzyme Activity of AST, ALT and LDH in Cell Supernatant
3.5. Morphological Observation on Cells
3.6. Data Analysis
4. Conclusions
Acknowledgements
References
- Liu, ZJ; Zhu, ZY; Roberg, K; Faras, A; Guise, K; Kapuscinski, AR; Hackett, PB. Isolation and characterization of beta-actin gene of carp (Cyprinus carpio). DNA Seq 1990, 1, 125–136. [Google Scholar]
- Hutanasu, C; Sfarti, C; Trifan, A; Hutanasu, M; Stanciu, C. Aflatoxin contamination of food: additional risk factor for chronic liver diseases. Rev. Med. Chir. Soc. Med. Nat. Iasi 2009, 113, 1061–1065. [Google Scholar]
- Tchana, AN; Moundipa, PF; Tchouanguep, FM. Aflatoxin contamination in food and body fluids in relation to malnutrition and cancer status in Cameroon. Int. J. Environ. Res. Public Health 2010, 7, 178–188. [Google Scholar]
- Kitya, D; Bbosa, GS; Mulogo, E. Aflatoxin levels in common foods of South Western Uganda: a risk factor to hepatocellular carcinoma. Eur. J. Cancer Care (Engl.) 2010, 19, 516–521. [Google Scholar]
- Kumagai, S; Nakajima, M; Tabata, S; Ishikuro, E; Tanaka, T; Norizuki, H; Itoh, Y; Aoyama, K; Fujita, K; Kai, S; Sato, T; Saito, S; Yoshiike, N; Sugita-Konishi, Y. Aflatoxin and ochratoxin A contamination of retail foods and intake of these mycotoxins in Japan. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess 2008, 25, 1101–1106. [Google Scholar]
- Hussain, M; Gabal, MA; Wilson, T; Summerfelt, RC. Effect of aflatoxin-contaminated feed on morbidity and residues in walleye fish. Vet. Hum. Toxicol 1993, 35, 396–398. [Google Scholar]
- Halver, JE; Ashley, LM; Smith, RR. Aflatoxicosis in coho salmon. Natl. Cancer Inst. Monogr 1969, 31, 141–155. [Google Scholar]
- Gallagher, EP. Using salmonid microarrays to understand the dietary modulation of carcinogenesis in rainbow trout. Toxicol. Sci 2006, 90, 1–4. [Google Scholar]
- Bailey, GS; Williams, DE; Wilcox, JS; Loveland, PM; Coulombe, RA; Hendricks, JD. Aflatoxin B1 carcinogenesis and its relation to DNA adduct formation and adduct persistence in sensitive and resistant salmonid fish. Carcinogenesis 1988, 9, 1919–1926. [Google Scholar]
- Gallagher, EP; Eaton, DL. In vitro biotransformation of aflatoxin B1 (AFB1) in channel catfish liver. Toxicol. Appl. Pharmacol 1995, 132, 82–90. [Google Scholar]
- Abdel-Wahhab, MA; Hasan, AM; Aly, SE; Mahrous, KF. Adsorption of sterigmatocystin by montmorillonite and inhibition of its genotoxicity in the Nile tilapia fish (Oreachromis nilaticus). Mutat. Res 2005, 582, 20–27. [Google Scholar]
- Sato, S; Matsushima, T; Tanaka, N; Sugimura, T; Takashima, F. Hepatic tumors in the guppy (Lebistes reticulatus) induced by aflatoxin B1, dimethylintrosamine, and 2-acetylaminofluorene. J. Natl. Cancer Inst 1973, 50, 767–778. [Google Scholar]
- Sahoo, PK; Mukherjee, SC. The effect of dietary immunomodulation upon Edwardsiella tarda vaccination in healthy and immunocompromised Indian major carp (Labeo rohita). Fish Shellfish Immunol 2002, 12, 1–16. [Google Scholar]
- Wood, GE; Carter, L, Jr. Limited survey of deoxynivalenol in wheat and corn in the United States. J. Assoc. Off. Anal. Chem 1989, 72, 38–40. [Google Scholar]
- Rotter, BA; Prelusky, DB; Pestka, JJ. Toxicology of deoxynivalenol (vomitoxin). J. Toxicol. Environ. Health 1996, 48, 1–34. [Google Scholar]
- Ali, N; Sardjono; Yamashita, A; Yoshizawa, T. Natural co-occurrence of aflatoxins and Fusarium mycotoxins (fumonisins, deoxynivalenol, nivalenol and zearalenone) in corn from Indonesia. Food Addit. Contam 1998, 15, 377–384. [Google Scholar]
- Tacon, AG; Metian, M. Aquaculture feed and food safety. Ann. N. Y. Acad. Sci 2008, 1140, 50–59. [Google Scholar]
- Billings, PC; Uwaifo, AO; Heidelberger, C. Influence of benzoflavone on aflatoxin B1-induced cytotoxicity, mutation, and transformation of C3H/10T1/2 cells. Cancer Res 1983, 43, 2659–2663. [Google Scholar]
- Loquet, C; Wiebel, FJ. Geno- and cytotoxicity of nitrosamines, aflatoxin B1, and benzo[a]-pyrene in continuous cultures of rat hepatoma cells. Carcinogenesis 1982, 3, 1213–1218. [Google Scholar]
- Palanee, T; Dutton, MF; Chuturgoon, AA. Cytotoxicity of aflatoxin B1 and its chemically synthesised epoxide derivative on the A549 human epithelioid lung cell line. Mycopathologia 2001, 151, 155–159. [Google Scholar]
- Zuckerman, AJ; Tsiquaye, KN; Fulton, F. Tissue culture of human embryo liver cells and the cytotoxicity of aflatoxin B1. Br. J. Exp. Pathol 1967, 48, 20–27. [Google Scholar]
- Iwaki, M; Kitagawa, T; Akamatsu, Y; Aibara, K. Cytotoxic effects of aflatoxin B1 and its association with cellular components in chicken embryo primary cultured cells. Biochim. Biophys. Acta 1990, 1035, 146–153. [Google Scholar]
- Gabliks, J; Barter, S. Comparative cytotoxicity of aflatoxin B1 and saxitoxin in cell cultures. Mol. Toxicol 1987, 1, 209–216. [Google Scholar]
- Golli-Bennour, EE; Kouidhi, B; Bouslimi, A; Abid-Essefi, S; Hassen, W; Bacha, H. Cytotoxicity and genotoxicity induced by aflatoxin B1, ochratoxin A, and their combination in cultured Vero cells. J. Biochem. Mol. Toxicol 2010, 24, 42–50. [Google Scholar]
- Pal, L; Dublecz, K; Weber, M; Balogh, K; Erdelyi, M; Szigeti, G; Mezes, M. Effect of combined treatment with aflatoxin B1 and T-2 toxin and metabolites on some production traits and lipid peroxide status parameters of broiler chickens. Acta Vet. Hung 2009, 57, 75–84. [Google Scholar]
- Orsi, RB; Oliveira, CA; Dilkin, P; Xavier, JG; Direito, GM; Correa, B. Effects of oral administration of aflatoxin B1 and fumonisin B1 in rabbits (Oryctolagus cuniculus). Chem. Biol. Interact 2007, 170, 201–208. [Google Scholar]
- McKean, C; Tang, L; Tang, M; Billam, M; Wang, Z; Theodorakis, CW; Kendall, RJ; Wang, JS. Comparative acute and combinative toxicity of aflatoxin B1 and fumonisin B1 in animals and human cells. Food Chem. Toxicol 2006, 44, 868–876. [Google Scholar]
- Li, B; Guo, H. Study on the combined toxicity of aflatoxin B1 and deoxynivalenol. Wei Sheng Yan Jiu 2000, 29, 393–395. [Google Scholar]
- Yanhong, F; Chenghua, H; Guofang, L; Haibin, Z. Optimization of the isolation and cultivation of Cyprinus carpio primary hepatocytes. Cytotechnology 2008, 58, 85–92. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar]





© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
He, C.-H.; Fan, Y.-H.; Wang, Y.; Huang, C.-Y.; Wang, X.-C.; Zhang, H.-B. The Individual and Combined Effects of Deoxynivalenol and Aflatoxin B1 on Primary Hepatocytes of Cyprinus Carpio. Int. J. Mol. Sci. 2010, 11, 3760-3768. https://doi.org/10.3390/ijms11103760
He C-H, Fan Y-H, Wang Y, Huang C-Y, Wang X-C, Zhang H-B. The Individual and Combined Effects of Deoxynivalenol and Aflatoxin B1 on Primary Hepatocytes of Cyprinus Carpio. International Journal of Molecular Sciences. 2010; 11(10):3760-3768. https://doi.org/10.3390/ijms11103760
Chicago/Turabian StyleHe, Cheng-Hua, Yan-Hong Fan, Ying Wang, Chao-Ying Huang, Xi-Chun Wang, and Hai-Bin Zhang. 2010. "The Individual and Combined Effects of Deoxynivalenol and Aflatoxin B1 on Primary Hepatocytes of Cyprinus Carpio" International Journal of Molecular Sciences 11, no. 10: 3760-3768. https://doi.org/10.3390/ijms11103760