Polyunsaturated Fatty Acids and Modulation of Cholesterol Homeostasis in THP-1 Macrophage-Derived Foam Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
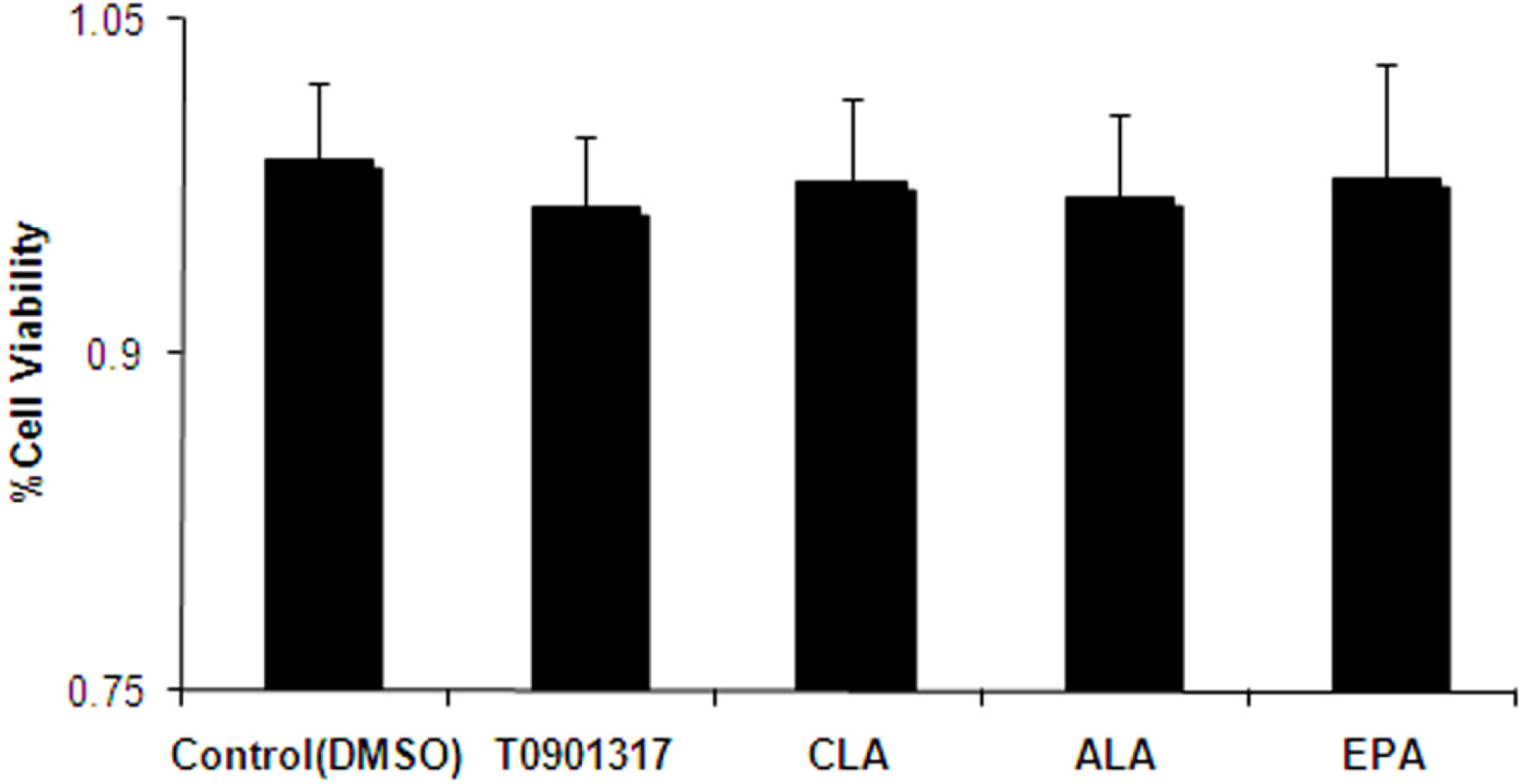
2.1. Toxicology Assay
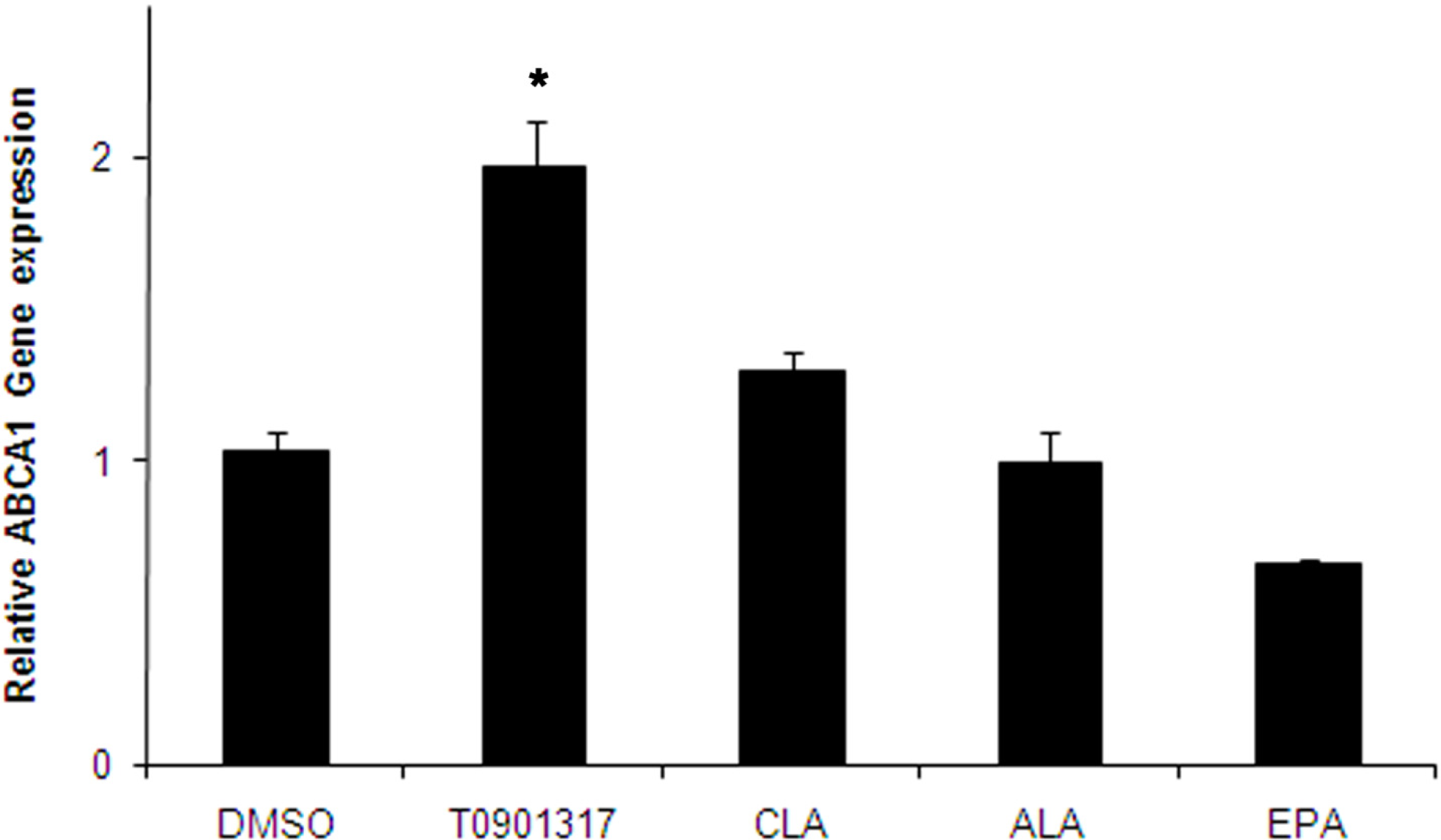
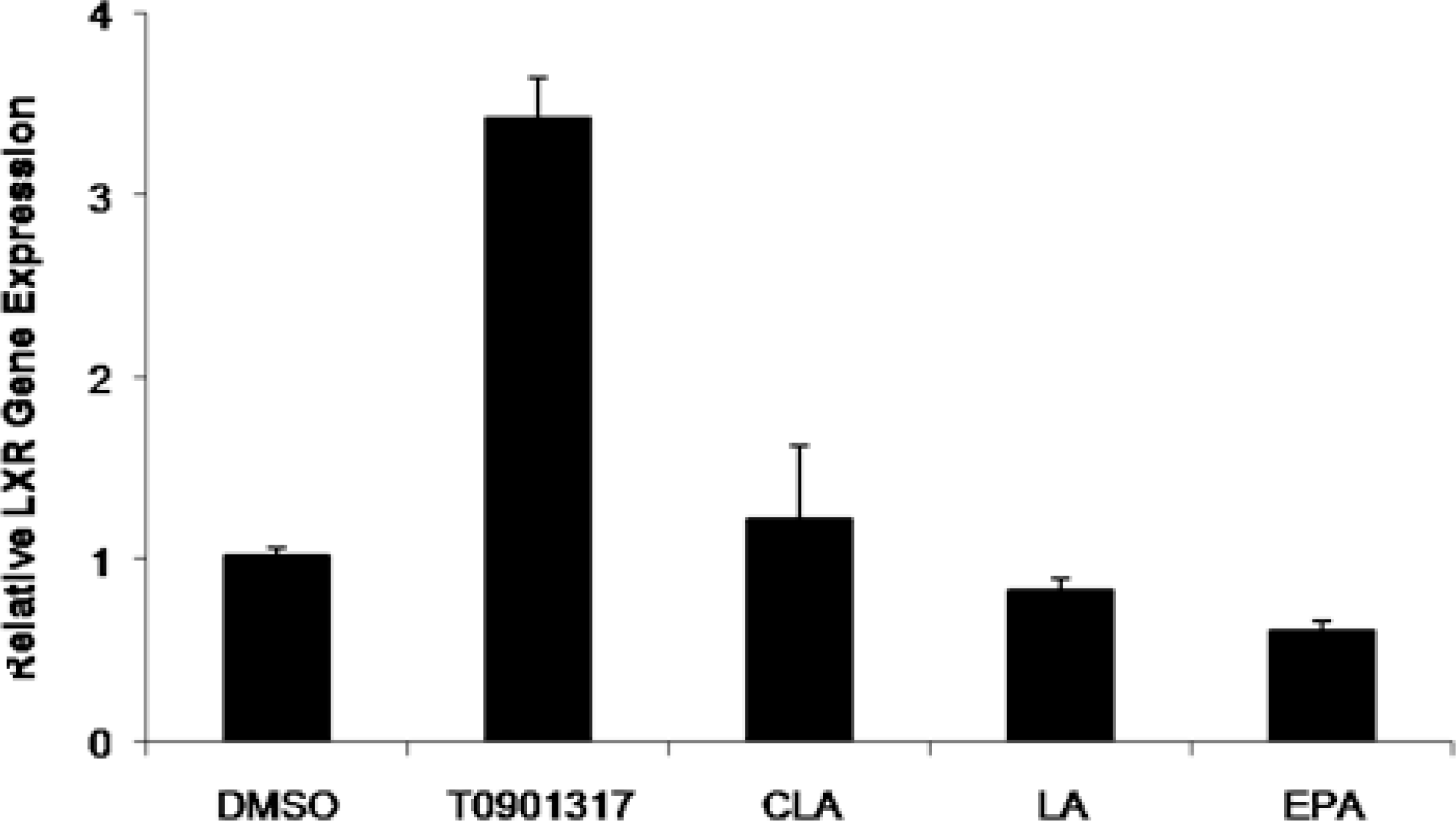
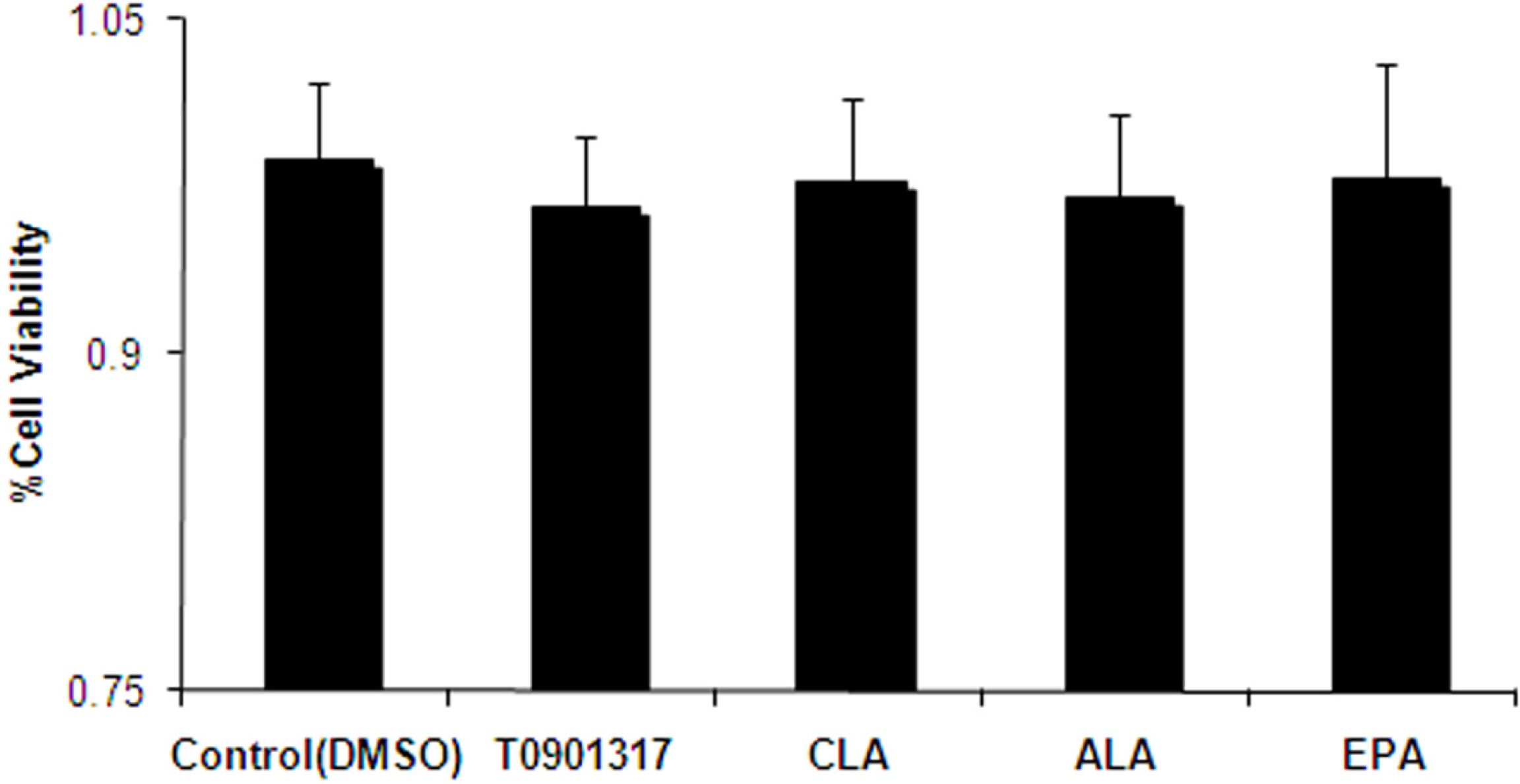
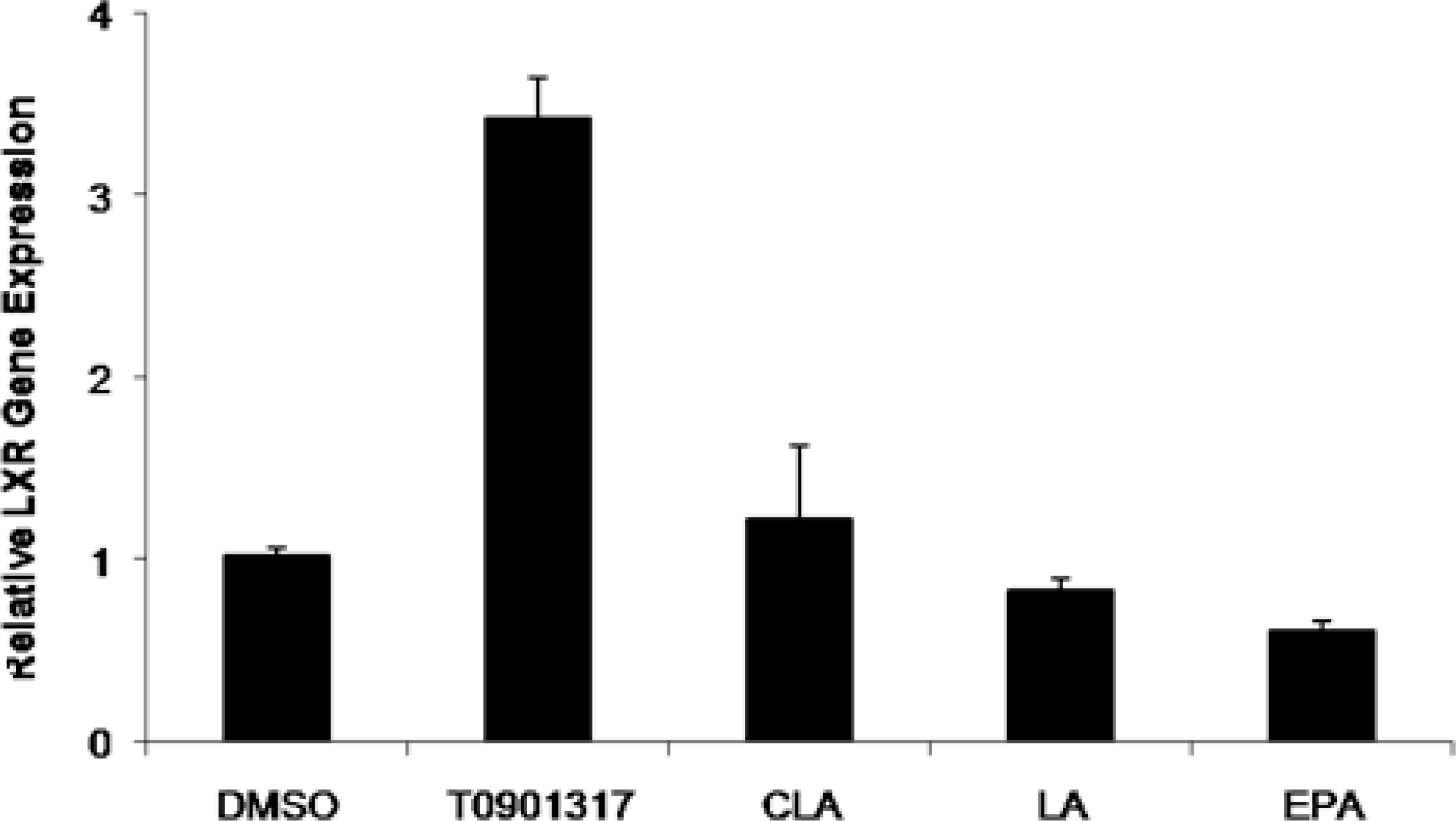
2.2. Effect of Fatty Acids Treatment on the Expression of LXRα and ABCA1 Genes in THP-1-Derived Macrophage Foam Cells
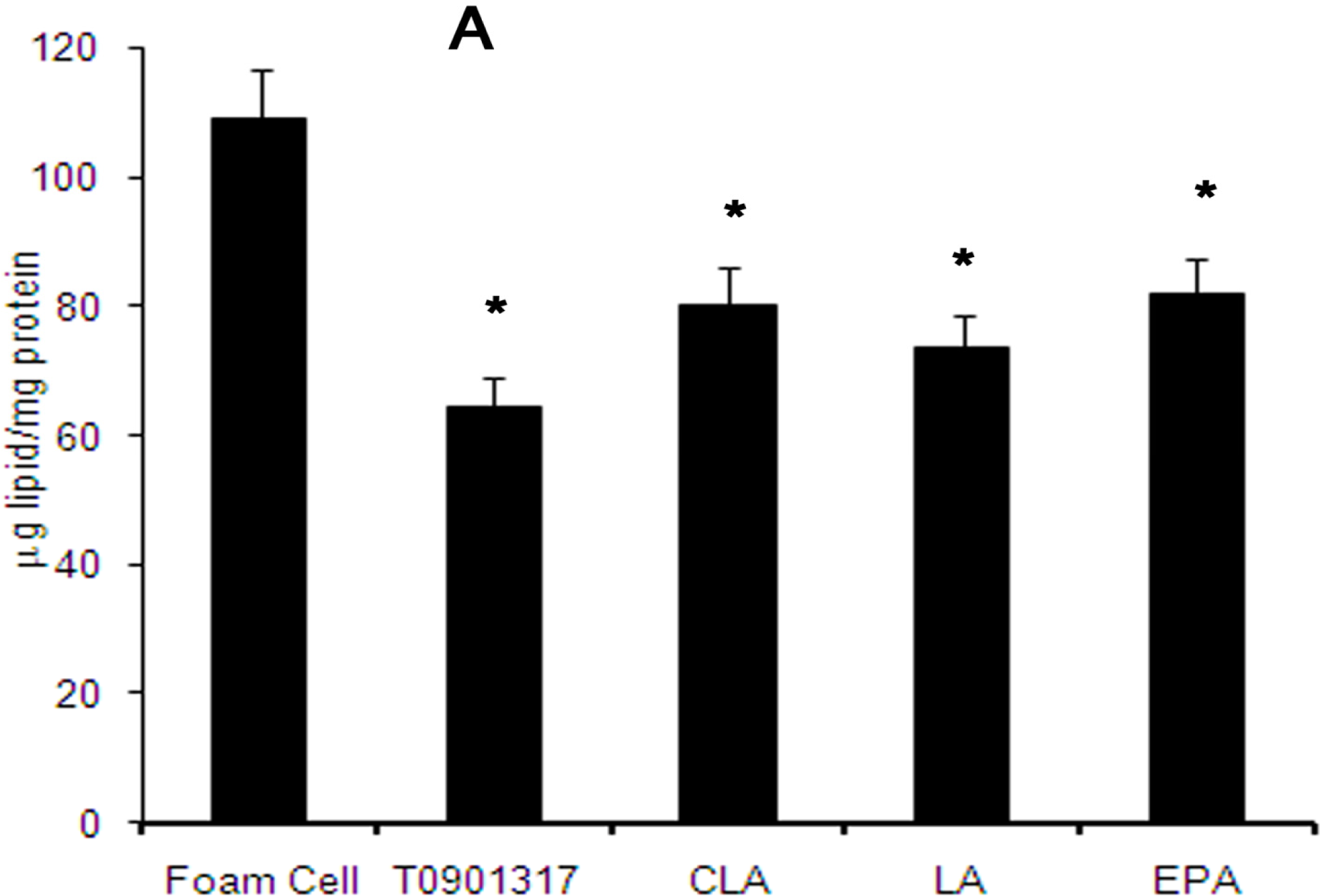
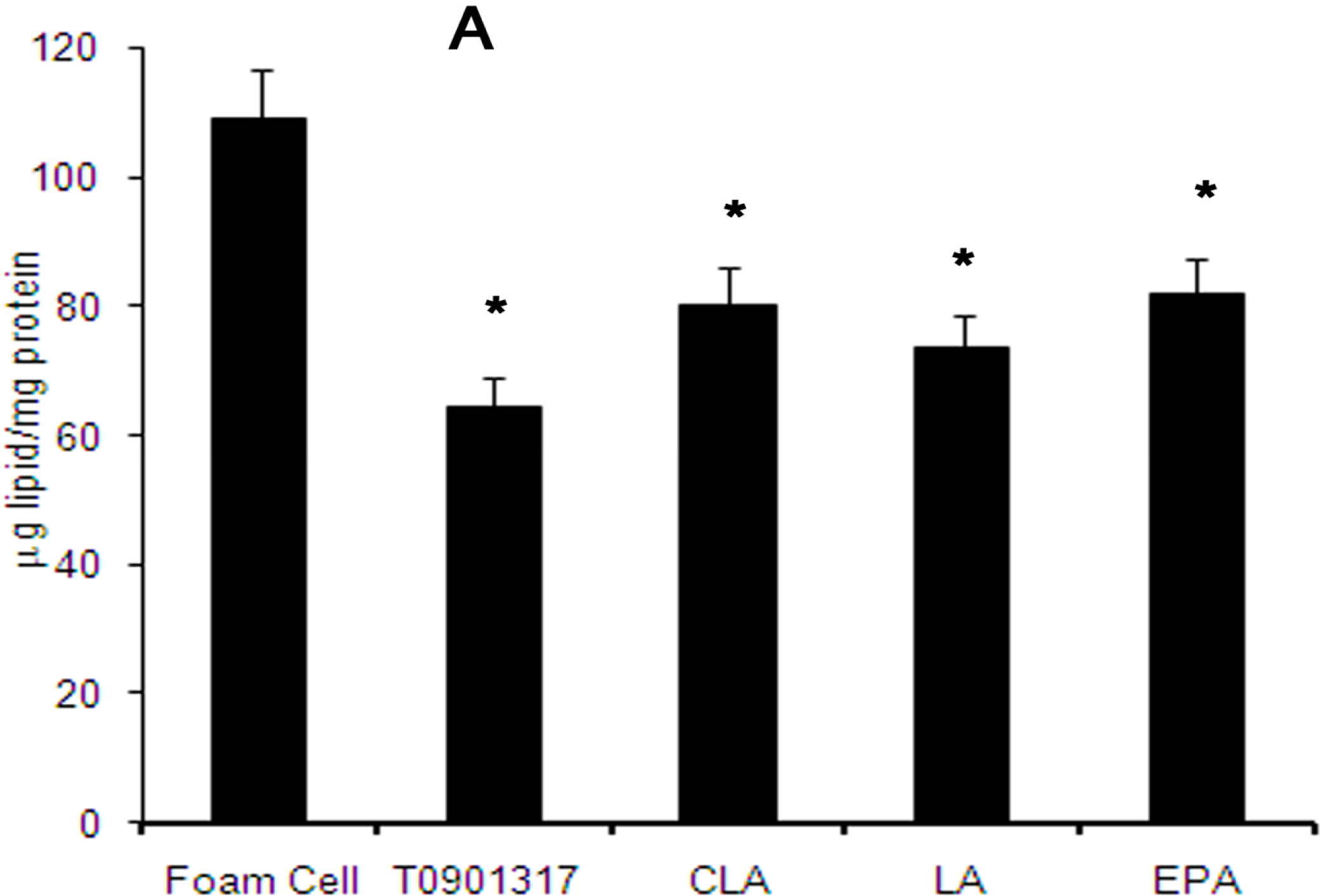
2.3. Distribution of Total Cellular Cholesterol in Cholesterol-Loaded Macrophages
3. Materials and Methods
3.1. Chemicals
3.2. Cell Culture
3.3. Isolation and Acetylating of LDL
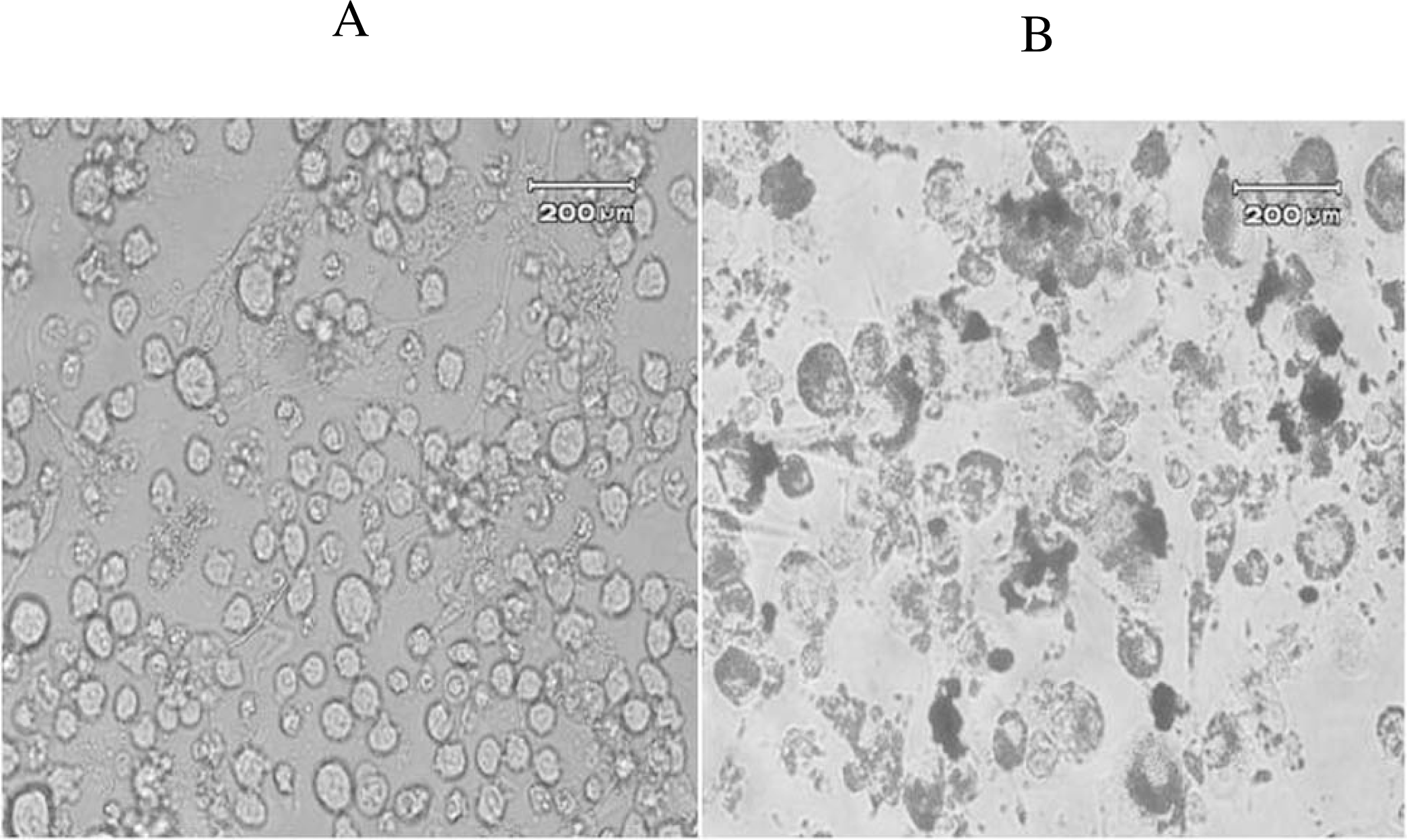
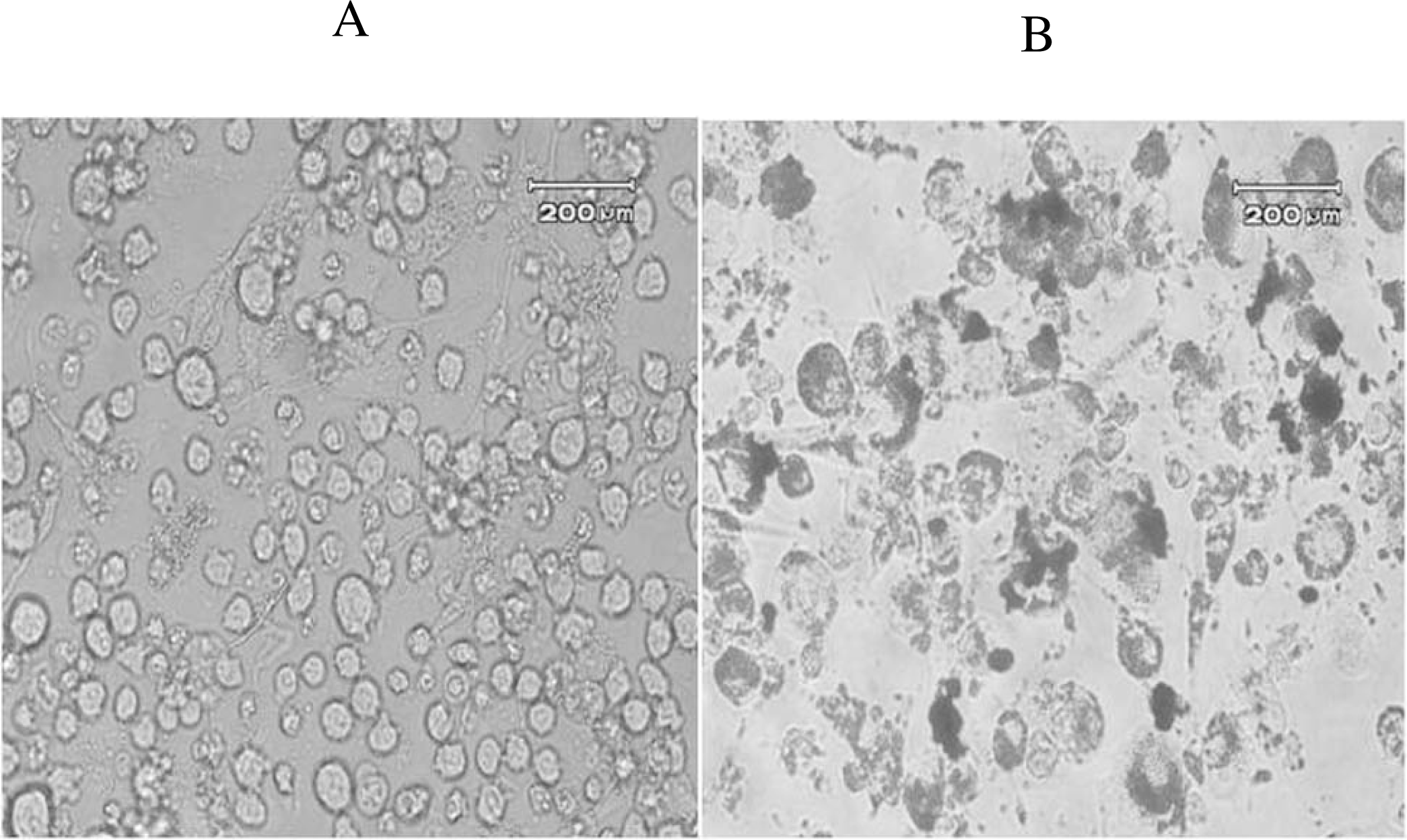
3.4. Oil Red-O Staining
3.5. Fatty Acids and T0901317 Treatment
3.6. RNA Isolation and Real-Time RT-PCR
3.7. Toxicology Assay
3.8. Intracellular Cholesterol Measurement
3.9. Statistical Analysis
4. Conclusion
References
- Ross, R. Atherosclerosis-an inflammatory disease. N. Engl. J. Med 1999, 340, 115–126. [Google Scholar]
- von Eckardstein, A; Nofer, JR; Assmann, G. High density lipoproteins and arteriosclerosis. Role of cholesterol efflux and reverse cholesterol transport. Arterioscler. Thromb. Vasc. Biol 2001, 21, 13–27. [Google Scholar]
- Bodzioch, M; Orso, E; Klucken, J; Langmann, T; Bottcher, A; Diederich, W; Drobnik, W; Barlage, S; Buchler, C; Porsch-Ozcurumez, M; et al. The gene encoding ATP-binding cassette transporter 1 is mutated in Tangier disease. Nat. Genet 1999, 22, 347–351. [Google Scholar]
- Brooks-Wilson, A; Marcil, M; Clee, SM; Zhang, LH; Roomp, K; van Dam, M; Yu, L; Brewer, C; Collins, JA; Molhuizen, HO; et al. Mutations in ABC1 in Tangier disease and familial high-density lipoprotein deficiency. Nat. Genet 1999, 22, 336–345. [Google Scholar]
- Rust, S; Rosier, M; Funke, H; Real, J; Amoura, Z; Piette, JC; Deleuze, JF; Brewer, HB; Duverger, N; Denefle, P; et al. Tangier disease is caused by mutations in the gene encoding ATP-binding cassette transporter 1. Nat. Genet 1999, 22, 352–355. [Google Scholar]
- Costet, P; Luo, Y; Wang, N; Tall, AR. Sterol-dependent transactivation of the ABC1 promoter by the liver X receptor/retinoid X receptor. J. Biol. Chem 2000, 275, 28240–28245. [Google Scholar]
- Repa, JJ; Mangelsdorf, DJ. The role of orphan nuclear receptors in the regulation of cholesterol homeostasis. Annu. Rev. Cell Dev. Biol 2000, 16, 459–481. [Google Scholar]
- Schwartz, K; Lawn, RM; Wade, DP. ABC1 gene expression and ApoA-I-mediated cholesterol efflux are regulated by LXR. Biochem. Biophys. Res. Commun 2000, 274, 794–802. [Google Scholar]
- Venkateswaran, A; Laffitte, BA; Joseph, SB; Mak, PA; Wilpitz, DC; Edwards, PA; Tontonoz, P. Control of cellular cholesterol efflux by the nuclear oxysterol receptor LXR alpha. Proc. Natl. Acad. Sci. USA 2000, 97, 12097–12102. [Google Scholar]
- Uehara, Y; Engel, T; Li, Z; Goepfert, C; Rust, S; Zhou, X; Langer, C; Schachtrup, C; Wiekowski, J; Lorkowski, S; Assmann, G; von Eckardstein, A. Polyunsaturated fatty acids and acetoacetate downregulate the expression of the ATP-binding cassette transporter A1. Diabetes 2002, 51, 2922–2928. [Google Scholar]
- Suresh, Y; Das, UN. Long-chain polyunsaturated fatty acids and chemically induced diabetes mellitus. Effect of omega-3 fatty acids. Nutrition 2003, 19, 213–228. [Google Scholar]
- Hwang, D. Fatty acids and immune responses--a new perspective in searching for clues to mechanism. Annu. Rev. Nutr 2000, 20, 431–456. [Google Scholar]
- Ntambi, JM; Choi, Y; Park, Y; Peters, JM; Pariza, MW. Effects of conjugated linoleic acid (CLA) on immune responses, body composition and stearoyl-CoA desaturase. Can. J. Appl. Physiol 2002, 27, 617–628. [Google Scholar]
- Harris, WS; Lu, G; Rambjor, GS; Walen, AI; Ontko, JA; Cheng, Q; Windsor, SL. Influence of n-3 fatty acid supplementation on the endogenous activities of plasma lipases. Am. J. Clin. Nutr 1997, 66, 254–260. [Google Scholar]
- Rambjor, GS; Walen, AI; Windsor, SL; Harris, WS. Eicosapentaenoic acid is primarily responsible for hypotriglyceridemic effect of fish oil in humans. Lipids 1996, 31, S45–S49. [Google Scholar]
- Schwartz, J. Role of polyunsaturated fatty acids in lung disease. Am. J. Clin. Nutr 2000, 71, 393S–396S. [Google Scholar]
- Siscovick, DS; Raghunathan, T; King, I; Weinmann, S; Bovbjerg, VE; Kushi, L; Cobb, LA; Copass, MK; Psaty, BM; Lemaitre, R; Retzlaff, B; Knopp, RH. Dietary intake of long-chain n-3 polyunsaturated fatty acids and the risk of primary cardiac arrest. Am. J. Clin. Nutr 2000, 71, 208S–212S. [Google Scholar]
- Skerrett, PJ; Hennekens, CH. Consumption of fish and fish oils and decreased risk of stroke. Prev. Cardiol 2003, 6, 38–41. [Google Scholar]
- von Schacky, C. n-3 fatty acids and the prevention of coronary atherosclerosis. Am. J. Clin. Nutr 2000, 71, 224S–227S. [Google Scholar]
- Toomey, S; Roche, H; Fitzgerald, D; Belton, O. Regression of pre-established atherosclerosis in the apoE-/- mouse by conjugated linoleic acid. Biochem. Soc. Trans 2003, 31, 1075–1079. [Google Scholar]
- Hu, YW; Ma, X; Li, XX; Liu, XH; Xiao, J; Mo, ZC; Xiang, J; Liao, DF; Tang, CK. Eicosapentaenoic acid reduces ABCA1 serine phosphorylation and impairs ABCA1-dependent cholesterol efflux through cyclic AMP/protein kinase A signaling pathway in THP-1 macrophage-derived foam cells. Atherosclerosis 2009, 204, e35–e43. [Google Scholar]
- Lu, TT; Repa, JJ; Mangelsdorf, DJ. Orphan nuclear receptors as eLiXiRs and FiXeRs of sterol metabolism. J. Biol. Chem 2001, 276, 37735–37738. [Google Scholar]
- Joseph, SB; Laffitte, BA; Patel, PH; Watson, MA; Matsukuma, KE; Walczak, R; Collins, JL; Osborne, TF; Tontonoz, P. Direct and indirect mechanisms for regulation of fatty acid synthase gene expression by liver X receptors. J. Biol. Chem 2002, 277, 11019–11025. [Google Scholar]
- Ou, J; Tu, H; Shan, B; Luk, A; DeBose-Boyd, RA; Bashmakov, Y; Goldstein, JL; Brown, MS. Unsaturated fatty acids inhibit transcription of the sterol regulatory element-binding protein-1c (SREBP-1c) gene by antagonizing ligand-dependent activation of the LXR. Proc. Natl. Acad. Sci. USA 2001, 98, 6027–6032. [Google Scholar]
- Schultz, JR. Role of LXRs in control of lipogenesis. Genes Dev 2000, 14, 2831–2838. [Google Scholar]
- Laffitte, BA; Repa, JJ; Joseph, SB; Wilpitz, DC; Kast, HR; Mangelsdorf, DJ; Tontonoz, P. LXRs control lipid-inducible expression of the apolipoprotein E gene in macrophages and adipocytes. Proc. Natl. Acad. Sci. USA 2001, 98, 507–512. [Google Scholar]
- Venkateswaran, A. Human white/murine ABC8 mRNA levels are highly induced in lipid-loaded macrophages. A transcriptional role for specific oxysterols. J. Biol. Chem 2000, 275, 14700–14707. [Google Scholar]
- Joseph, SB; McKilligin, E; Pei, L; Watson, MA; Collins, AR; Laffitte, BA; Chen, M; Noh, G; Goodman, J; Hagger, GN; et al. Synthetic LXR ligand inhibits the development of atherosclerosis in mice. Proc. Natl. Acad. Sci. USA 2002, 99, 7604–7609. [Google Scholar]
- Tobin, KA; Steineger, HH; Alberti, S; Spydevold, O; Auwerx, J; Gustafsson, JA; Nebb, HI. Cross-talk between fatty acid and cholesterol metabolism mediated by liver X receptor-alpha. Mol. Endocrinol 2000, 14, 741–752. [Google Scholar]
- Pawar, A; Botolin, D; Mangelsdorf, DJ; Jump, DB. The role of liver X receptor-alpha in the fatty acid regulation of hepatic gene expression. J. Biol. Chem 2003, 278, 40736–40743. [Google Scholar]
- Rustan, AC; Nossen, JO; Osmundsen, H; Drevon, CA. Eicosapentaenoic acid inhibits cholesterol esterification in cultured parenchymal cells and isolated microsomes from rat liver. J. Biol. Chem 1988, 263, 8126–8132. [Google Scholar]
- Pal, S; Davis, PJ. Effects of different types of polyunsaturated fatty acids on cholesterol esterification in human fibroblasts. Biochem. Int 1991, 25, 281–288. [Google Scholar]
- Uehara, Y; Miura, S; von Eckardstein, A; Abe, S; Fujii, A; Matsuo, Y; Rust, S; Lorkowski, S; Assmann, G; Yamada, T; Saku, K. Unsaturated fatty acids suppress the expression of the ATP-binding cassette transporter G1 (ABCG1) and ABCA1 genes via an LXR/RXR responsive element. Atherosclerosis 2007, 191, 11–21. [Google Scholar]
- Desvergne, B; Wahli, W. Peroxisome proliferator-activated receptors: Nuclear control of metabolism. Endocr. Rev 1999, 20, 649–688. [Google Scholar]
- Chinetti-Gbaguidi, G; Rigamonti, E; Helin, L; Mutka, AL; Lepore, M; Fruchart, JC; Clavey, V; Ikonen, E; Lestavel, S; Staels, B. Peroxisome proliferator-activated receptor alpha controls cellular cholesterol trafficking in macrophages. J. Lipid Res 2005, 46, 2717–2725. [Google Scholar]
- Ringseis, R; Wen, G; Saal, D; Eder, K. Conjugated linoleic acid isomers reduce cholesterol accumulation in acetylated LDL-induced mouse RAW264.7 macrophage-derived foam cells. Lipids 2008, 43, 913–923. [Google Scholar]
- Moya-Camarena, SY; Vanden Heuvel, JP; Blanchard, SG; Leesnitzer, LA; Belury, MA. Conjugated linoleic acid is a potent naturally occurring ligand and activator of PPARalpha. J. Lipid Res 1999, 40, 142614–142633. [Google Scholar]
- Hirakata, M; Tozawa, R; Imura, Y; Sugiyama, Y. Comparison of the effects of pioglitazone and rosiglitazone on macrophage foam cell formation. Biochem. Biophys. Res. Commun 2004, 323, 782–788. [Google Scholar]
- Davis, PJ. n-3 and n-6 polyunsaturated fatty acids have different effects on acyl-CoA: Cholesterol acyltransferase in J774 macrophages. Biochem. Cell Biol 1992, 70, 1313–1318. [Google Scholar]
- Pietsch, A; Weber, C; Goretzki, M; Lornz, RL. N-3 but not N-6 fatty acids reduce the expression of the combined adhesion and scavenger receptor CD36 in human monocytic cells. Cell Biochem. Funct 1995, 13, 211–216. [Google Scholar]
- Chinetti, G; Gbaguidi, FG; Griglio, S; Mallat, A; Antonucci, M; Poulain, P; Chapman, J; Fruchart, J-C; Tedgui, A; Najib-Fruchart, J; Staels, B. CLA-1/SR-BI is expressed in atherosclerotic lesion macrophages and regulated by activators of peroxisome proliferator-activated receptors. Circulation 2000, 101, 2411–2417. [Google Scholar]
- Weldon, S; Mitchell, S; Kelleher, D; Gibney, MJ; Roche, HM. Conjugated linoleic acid and atherosclerosis: No effect on molecular markers of cholesterol homeostasis in THP-1 macrophages. Atherosclerosis 2004, 174, 261–273. [Google Scholar]
- Havel, RJ; Eder, HA; Bragdon, JH. The distribution and chemical composition of ultracentrifugally separated lipoproteins in human serum. J. Clin. Invest 1955, 34, 1345–1353. [Google Scholar]
- Fraenkel-Conrat, H. Methods Forinvestigatingthe Essentialgroups for Enzymeactivity. Method in Enzymology; s.p. colowick & n. o. kaplan: New York, NY, USA, 1957; Volume 4; pp. 247–269. [Google Scholar]
- Via, DP; Plant, AL; Craig, IF; Gotto, AM; Smith, LC. Metabolism of normal and modified low-density lipoproteins by macrophage cell lines of murine and human origin. Biochim. Biophys. Acta 1985, 833, 417–428. [Google Scholar]
- Scudiero, DA; Shoemaker, RH; Paull, KD; Monks, A; Tierney, S; Nofziger, TH; Currens, MJ; Seniff, D; Boyd, MR. Evaluation of a soluble tetrazolium/formazan assay for cell growth and drug sensitivity in culture using human and other tumor cell lines. Cancer Res 1988, 48, 4827–4833. [Google Scholar]
- Iida, KT; Kawakami, Y; Suzuki, H; Sone, H; Shimano, H; Toyoshima, H; Okuda, Y; Yamada, N. PPAR gamma ligands, troglitazone and pioglitazone, up-regulate expression of HMG-CoA synthase and HMG-CoA reductase gene in THP-1 macrophages. FEBS Lett 2002, 520, 177–181. [Google Scholar]
- Kehrer, JP; Biswal, SS; La, E; Thuillier, P; Datta, K; Fischer, SM; Vanden Heuvel, JP. Inhibition of peroxisome-proliferator-activated receptor (PPAR)alpha by MK886. Biochem. J 2001, 356, 899–906. [Google Scholar]
- Bradford, MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem 1976, 72, 248–254. [Google Scholar]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Salehipour, M.; Javadi, E.; Reza, J.Z.; Doosti, M.; Rezaei, S.; Paknejad, M.; Nejadi, N.; Heidari, M. Polyunsaturated Fatty Acids and Modulation of Cholesterol Homeostasis in THP-1 Macrophage-Derived Foam Cells. Int. J. Mol. Sci. 2010, 11, 4660-4672. https://doi.org/10.3390/ijms11114660
Salehipour M, Javadi E, Reza JZ, Doosti M, Rezaei S, Paknejad M, Nejadi N, Heidari M. Polyunsaturated Fatty Acids and Modulation of Cholesterol Homeostasis in THP-1 Macrophage-Derived Foam Cells. International Journal of Molecular Sciences. 2010; 11(11):4660-4672. https://doi.org/10.3390/ijms11114660
Chicago/Turabian StyleSalehipour, Masoud, Ebrahim Javadi, Javad Zavvar Reza, Mahmoud Doosti, Shahla Rezaei, Malihe Paknejad, Naser Nejadi, and Mansour Heidari. 2010. "Polyunsaturated Fatty Acids and Modulation of Cholesterol Homeostasis in THP-1 Macrophage-Derived Foam Cells" International Journal of Molecular Sciences 11, no. 11: 4660-4672. https://doi.org/10.3390/ijms11114660