Lutein Protects RGC-5 Cells Against Hypoxia and Oxidative Stress

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
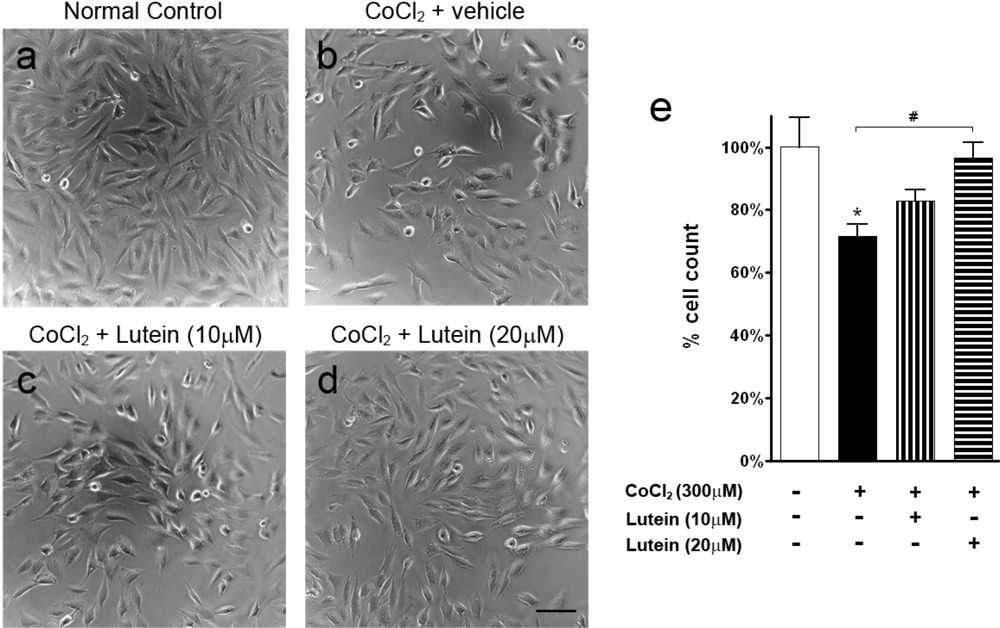
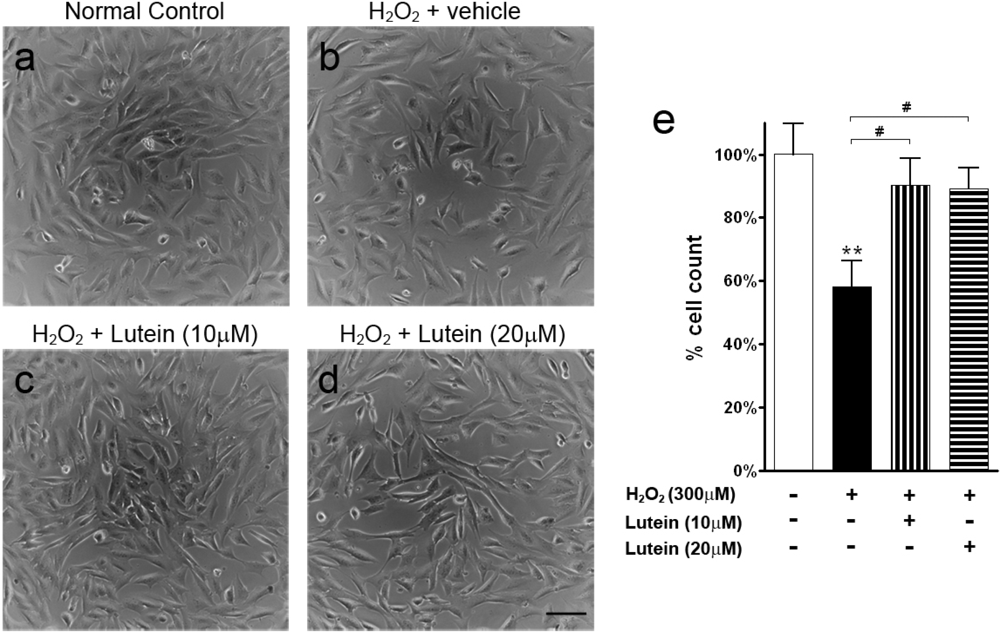
2.1. Results
2.2. Discussion
3. Experimental Section
4. Conclusions
Acknowledgments
References and Notes
- Osborne, NN; Casson, RJ; Wood, JPM; Chidlow, G; Graham, M; Melena, J. Retinal ischemia: mechanisms of damage and potential therapeutic strategies. Prog. Retin. Eye Res 2004, 23, 91–147. [Google Scholar]
- McKinnon, SJ. Glaucoma, apoptosis, and neuroprotection. Curr. Opin. Ophthalmol 1997, 8, 28–37. [Google Scholar]
- Tezel, G. Oxidative stress in glaucomatous neurodegeneration: mechanisms and consequences. Prog. Retin. Eye Res 2006, 25, 490–513. [Google Scholar]
- Nakajima, Y; Inokuchi, Y; Nishi, M; Shimazawa, M; Otsubo, K; Hara, H. Coenzyme Q10 protects retinal cells against oxidative stress in vitro and in vivo. Brain Res 2008, 1226, 226–233. [Google Scholar]
- Nakajima, Y; Inokuchi, Y; Shimazawa, M; Otsubo, K; Ishibashi, T; Hara, H. Astaxanthin, a dietary carotenoid, protects retinal cells against oxidative stress in vitro and in mice in vivo. J. Pharm. Pharmacol 2008, 60, 1365–1374. [Google Scholar]
- Aydemir, O; Celebi, S; Yilmaz, T; Yekeler, H; Kukner, AS. Protective effects of vitamin E forms (alpha-tocopherol, gamma-tocopherol and d-alpha-tocopherol polyethylene glycol 1000 succinate) on retinal edema during ischemia-reperfusion injury in the guinea pig retina. Int. Ophthalmol 2004, 25, 283–289. [Google Scholar]
- Calvo, MM. Lutein: A valuable ingredient of fruit and vegetables. Crit. Rev. Food Sci. Nutr 2005, 45, 671–696. [Google Scholar]
- Carpentier, S; Knaus, M; Suh, M. Associations between lutein, zeaxanthin, and age-related macular degeneration: an overview. Crit. Rev. Food Sci. Nutr 2009, 49, 313–326. [Google Scholar]
- Ojima, F; Sakamoto, H; Ishiguro, Y; Terao, J. Consumption of carotenoids in photosensitized oxidation of human plasma and plasma low-density lipoprotein. Free Radic. Biol. Med 1993, 15, 377–384. [Google Scholar]
- Alves-Rodrigues, A; Shao, A. The science behind lutein. Toxicol. Lett 2004, 150, 57–83. [Google Scholar]
- Sommerburg, O; Keunen, JE; Bird, AC; van Kuijk, FJ. Fruits and vegetables that are sources for lutein and zeaxanthin: the macular pigment in human eyes. Br. J. Ophthalmol 1998, 82, 907–910. [Google Scholar]
- Li, SY; Fu, ZJ; Ma, H; Jang, WC; So, KF; Wong, D; Lo, AC. Effect of lutein on retinal neurons and oxidative stress in a model of acute retinal ischemia/reperfusion. Invest. Ophthalmol. Vis. Sci 2009, 50, 836–843. [Google Scholar]
- Krishnamoorthy, RR; Agarwal, P; Prasanna, G; Vopat, K; Lambert, W; Sheedlo, HJ; Pang, IH; Shade, D; Wordinger, RJ; Yorio, T; Clark, AF; Agarwal, N. Characterization of a transformed rat retinal ganglion cell line. Brain Res. Mol. Brain Res 2001, 86, 1–12. [Google Scholar]
- Das, S; Lin, D; Jena, S; Shi, A; Battina, S; Hua, DH; Allbaugh, R; Takemoto, DJ. Protection of retinal cells from ischemia by a novel gap junction inhibitor. Biochem. Biophys. Res. Commun 2008, 373, 504–508. [Google Scholar]
- Guo, M; Song, LP; Jiang, Y; Liu, W; Yu, Y; Chen, GQ. Hypoxia-mimetic agents desferrioxamine and cobalt chloride induce leukemic cell apoptosis through different hypoxia-inducible factor-1alpha independent mechanisms. Apoptosis 2006, 11, 67–77. [Google Scholar]
- Whitlock, NA; Agarwal, N; Ma, JX; Crosson, CE. Hsp27 upregulation by HIF-1 signaling offers protection against retinal ischemia in rats. Invest. Ophthalmol. Vis. Sci 2005, 46, 1092–1098. [Google Scholar]
- Yang, SJ; Pyen, J; Lee, I; Lee, H; Kim, Y; Kim, T. Cobalt chloride-induced apoptosis and extracellular signal-regulated protein kinase 1/2 activation in rat C6 glioma cells. J. Biochem. Mol. Biol 2004, 37, 480–486. [Google Scholar]
- Zhu, X; Zhou, W; Cui, Y; Zhu, L; Li, J; Feng, X; Shao, B; Qi, H; Zheng, J; Wang, H; Chen, H. Pilocarpine protects cobalt chloride-induced apoptosis of RGC-5 cells: Involvement of muscarinic receptors and HIF-1alpha pathway. Cell Mol. Neurobiol 2009, 30, 427–435. [Google Scholar]
- Goldberg, MA; Schneider, TJ. Similarities between the oxygen-sensing mechanisms regulating the expression of vascular endothelial growth factor and erythropoietin. J. Biol. Chem 1994, 269, 4355–4359. [Google Scholar]
- An, WG; Kanekal, M; Simon, MC; Maltepe, E; Blagosklonny, MV; Neckers, LM. Stabilization of wild-type p53 by hypoxia-inducible factor 1alpha. Nature 1998, 392, 405–408. [Google Scholar]
- Zou, W; Yan, M; Xu, W; Huo, H; Sun, L; Zheng, Z; Liu, X. Cobalt chloride induces PC12 cells apoptosis through reactive oxygen species and accompanied by AP-1 activation. J. Neurosci. Res 2001, 64, 646–653. [Google Scholar]
- Hara, A; Niwa, M; Aoki, H; Kumada, M; Kunisada, T; Oyama, T; Yamamoto, T; Kozawa, O; Mori, H. A new model of retinal photoreceptor cell degeneration induced by a chemical hypoxia-mimicking agent, cobalt chloride. Brain Res 2006, 1109, 192–200. [Google Scholar]
- Zhu, X; Zhou, W; Cui, Y; Zhu, L; Li, J; Xia, Z; Shao, B; Wang, H; Chen, H. Muscarinic activation attenuates abnormal processing of beta-amyloid precursor protein induced by cobalt chloride-mimetic hypoxia in retinal ganglion cells. Biochem. Biophys. Res. Commun 2009, 384, 110–113. [Google Scholar]
- Bazan, NG. The metabolism of omega-3 polyunsaturated fatty acids in the eye: the possible role of docosahexaenoic acid and docosanoids in retinal physiology and ocular pathology. Prog. Clin. Biol. Res 1989, 312, 95–112. [Google Scholar]
- Li, GY; Osborne, NN. Oxidative-induced apoptosis to an immortalized ganglion cell line is caspase independent but involves the activation of poly(ADP-ribose)polymerase and apoptosis-inducing factor. Brain Res 2008, 1188, 35–43. [Google Scholar]
- Shimazawa, M; Nakajima, Y; Mashima, Y; Hara, H. Docosahexaenoic acid (DHA) has neuroprotective effects against oxidative stress in retinal ganglion cells. Brain Res 2009, 1251, 269–275. [Google Scholar]
- Goldberg, J; Flowerdew, G; Smith, E; Brody, JA; Tso, MO. Factors associated with age-related macular degeneration. An analysis of data from the first National Health and Nutrition Examination Survey. Am. J. Epidemiol 1988, 128, 700–710. [Google Scholar]
- Richer, S; Devenport, J; Lang, JC. LAST II: Differential temporal responses of macular pigment optical density in patients with atrophic age-related macular degeneration to dietary supplementation with xanthophylls. Optometry 2007, 78, 213–219. [Google Scholar]
- Richer, S; Stiles, W; Statkute, L; Pulido, J; Frankowski, J; Rudy, D; Pei, K; Tsipursky, M; Nyland, J. Double-masked, placebo-controlled, randomized trial of lutein and antioxidant supplementation in the intervention of atrophic age-related macular degeneration: the Veterans LAST study (Lutein Antioxidant Supplementation Trial). Optometry 2004, 75, 216–230. [Google Scholar]
- Kruger, CL; Murphy, M; DeFreitas, Z; Pfannkuch, F; Heimbach, J. An innovative approach to the determination of safety for a dietary ingredient derived from a new source: case study using a crystalline lutein product. Food Chem. Toxicol 2002, 40, 1535–1549. [Google Scholar]
- Muriach, M; Bosch-Morell, F; Alexander, G; Blomhoff, R; Barcia, J; Arnal, E; Almansa, I; Romero, FJ; Miranda, M. Lutein effect on retina and hippocampus of diabetic mice. Free Radic. Biol. Med 2006, 41, 979–984. [Google Scholar]
- Sasaki, M; Ozawa, Y; Kurihara, T; Noda, K; Imamura, Y; Kobayashi, S; Ishida, S; Tsubota, K. Neuroprotective effect of an antioxidant, lutein, during retinal inflammation. Invest. Ophthalmol. Vis. Sci 2009, 50, 1433–1439. [Google Scholar]
- Nakajima, Y; Shimazawa, M; Otsubo, K; Ishibashi, T; Hara, H. Zeaxanthin, a retinal carotenoid, protects retinal cells against oxidative stress. Curr. Eye Res 2009, 34, 311–318. [Google Scholar]
- Kim, JH; Na, HJ; Kim, CK; Kim, JY; Ha, KS; Lee, H; Chung, HT; Kwon, HJ; Kwon, YG; Kim, YM. The non-provitamin A carotenoid, lutein, inhibits NF-kappaB-dependent gene expression through redox-based regulation of the phosphatidylinositol 3-kinase/PTEN/Akt and NF-kappaB-inducing kinase pathways: role of H2O2 in NF-kappaB activation. Free Radic. Biol. Med 2008, 45, 885–896. [Google Scholar]
- Bhosale, P; Li, B; Sharifzadeh, M; Gellermann, W; Frederick, JM; Tsuchida, K; Bernstein, PS. Purification and partial characterization of a lutein-binding protein from human retina. Biochemistry 2009, 48, 4798–4807. [Google Scholar]
- Bernstein, PS; Balashov, NA; Tsong, ED; Rando, RR. Retinal tubulin binds macular carotenoids. Invest. Ophthalmol. Vis. Sci 1997, 38, 167–175. [Google Scholar]
- Moorhouse, AJ; Li, S; Vickery, RM; Hill, MA; Morley, JW. A patch-clamp investigation of membrane currents in a novel mammalian retinal ganglion cell line. Brain Res 2004, 1003, 205–208. [Google Scholar]
- van Bergen, NJ; Wood, JP; Chidlow, G; Trounce, IA; Casson, RJ; Ju, WK; Weinreb, RN; Crowston, JG. Recharacterization of the RGC-5 retinal ganglion cell line. Invest. Ophthalmol. Vis. Sci 2009, 50, 4267–4272. [Google Scholar]
- Wood, JP; Chidlow, G; Tran, T; Crowston, J; Casson, RJ. A comparison of differentiation protocols for Rgc-5 cells. Invest Ophthalmol Vis Sci 2010. [Google Scholar]
- Rafi, MM; Shafaie, Y. Dietary lutein modulates inducible nitric oxide synthase (iNOS) gene and protein expression in mouse macrophage cells (RAW 264.7). Mol. Nutr. Food Res 2007, 51, 333–340. [Google Scholar]


© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, S.-Y.; Lo, A.C.Y. Lutein Protects RGC-5 Cells Against Hypoxia and Oxidative Stress. Int. J. Mol. Sci. 2010, 11, 2109-2117. https://doi.org/10.3390/ijms11052109
Li S-Y, Lo ACY. Lutein Protects RGC-5 Cells Against Hypoxia and Oxidative Stress. International Journal of Molecular Sciences. 2010; 11(5):2109-2117. https://doi.org/10.3390/ijms11052109
Chicago/Turabian StyleLi, Suk-Yee, and Amy C. Y. Lo. 2010. "Lutein Protects RGC-5 Cells Against Hypoxia and Oxidative Stress" International Journal of Molecular Sciences 11, no. 5: 2109-2117. https://doi.org/10.3390/ijms11052109
APA StyleLi, S.-Y., & Lo, A. C. Y. (2010). Lutein Protects RGC-5 Cells Against Hypoxia and Oxidative Stress. International Journal of Molecular Sciences, 11(5), 2109-2117. https://doi.org/10.3390/ijms11052109