ATP-Dependent Chromatin Remodeling Factors and Their Roles in Affecting Nucleosome Fiber Composition


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Chromatin Remodeling Machines and Their Impact on Nucleosome Structure
2.1. The Role of ATP-Dependent Chromatin Remodeling Factors in Nucleosome Positioning
2.2. Chromatin Remodeling Factorsin Replication-Coupled Nucleosome Assembly
2.3. Incorporation of Linker Histone H1
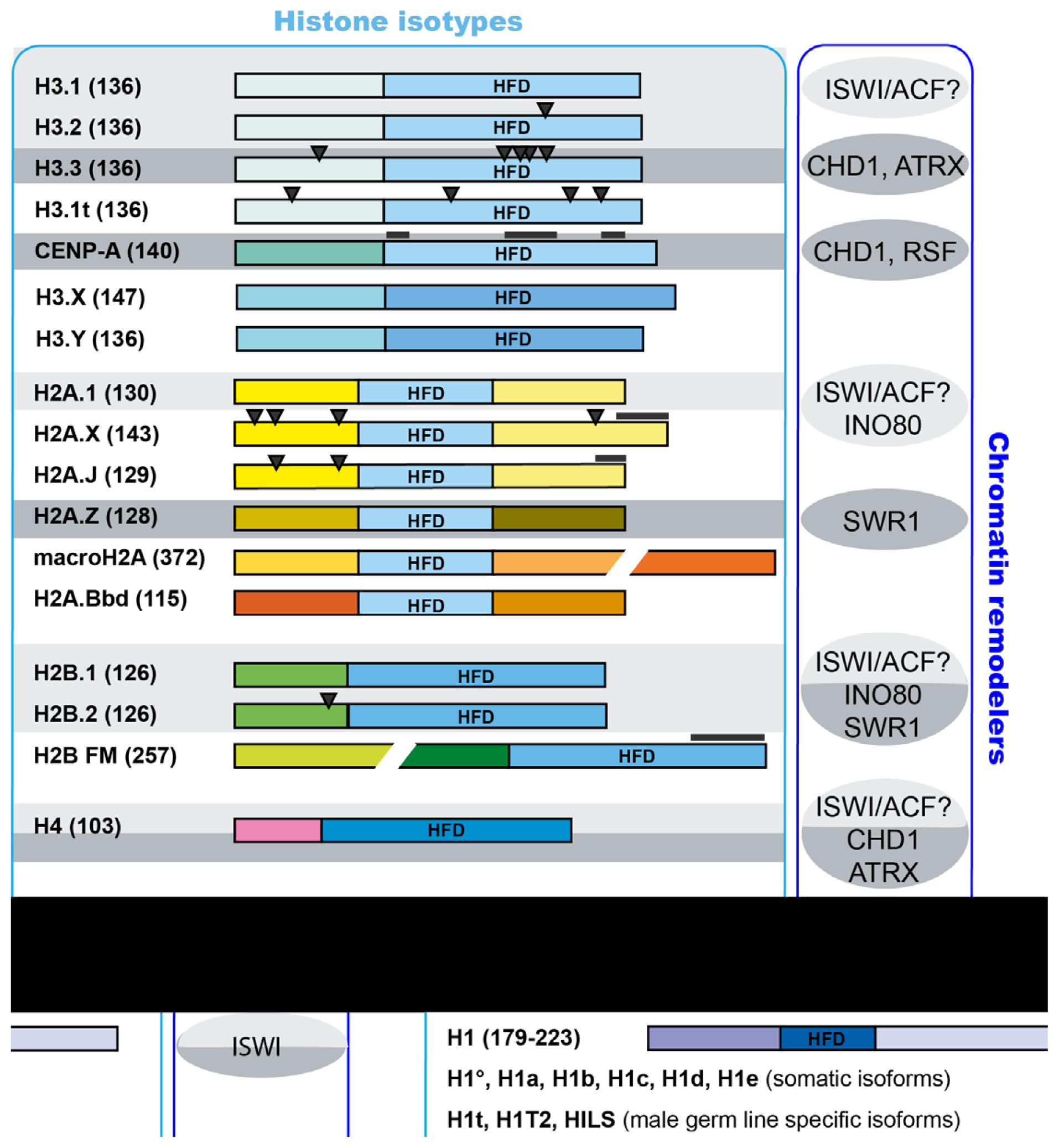
2.4. Incorporation of Variant Histones
2.5. Chromatin Remodelers and H3.3
2.6. Chromatin Remodelers and the Assembly of Centromeric Chromatin
2.7. Chromatin Remodelers and H2A.Z Exchange
3. Do Chromatin Remodeling Factors Incorporate Non-Histone Chromosomal Proteins?
4. Conclusion
Acknowledgments
References
- Luger, K; Mader, AW; Richmond, RK; Sargent, DF; Richmond, TJ. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature 1997, 389, 251–260. [Google Scholar]
- Olins, AL; Olins, DE. Spheroid chromatin units (v bodies). Science 1974, 183, 330–332. [Google Scholar]
- Woodcock, CL; Safer, JP; Stanchfield, JE. Structural repeating units in chromatin. I. Evidence for their general occurrence. Exp. Cell Res 1976, 97, 101–110. [Google Scholar]
- Jiang, C; Pugh, BF. Nucleosome positioning and gene regulation: advances through genomics. Nat. Rev. Genet 2009, 10, 161–172. [Google Scholar]
- Radman-Livaja, M; Rando, OJ. Nucleosome positioning: how is it established, and why does it matter? Dev. Biol 2010, 339, 258–266. [Google Scholar]
- Lavelle, C. Transcription elongation through a chromatin template. Biochimie 2007, 89, 516–527. [Google Scholar]
- Kouzarides, T. Chromatin modifications and their function. Cell 2007, 128, 693–705. [Google Scholar]
- Jin, J; Cai, Y; Li, B; Conaway, RC; Workman, JL; Conaway, JW; Kusch, T. In and out: histone variant exchange in chromatin. Trends Biochem. Sci 2005, 30, 680–687. [Google Scholar]
- Hock, R; Furusawa, T; Ueda, T; Bustin, M. HMG chromosomal proteins in development and disease. Trends Cell Biol 2007, 17, 72–79. [Google Scholar]
- Clapier, CR; Cairns, BR. The biology of chromatin remodeling complexes. Annu. Rev. Biochem 2009, 78, 273–304. [Google Scholar]
- Flaus, A; Martin, DM; Barton, GJ; Owen-Hughes, T. Identification of multiple distinct Snf2 subfamilies with conserved structural motifs. Nucleic Acids Res 2006, 34, 2887–2905. [Google Scholar]
- Gorbalenya, AE. Helicases: amino acid sequence comparisons and structure-function relationships. Curr. Opin. Struct. Biol 1993, 3, 419–429. [Google Scholar]
- Eberharter, A; Becker, PB. ATP-dependent nucleosome remodelling: factors and functions. J. Cell Sci 2004, 117, 3707–3711. [Google Scholar]
- Lusser, A; Kadonaga, JT. Chromatin remodeling by ATP-dependent molecular machines. Bioessays 2003, 25, 1192–1200. [Google Scholar]
- Bao, Y; Shen, X. INO80 subfamily of chromatin remodeling complexes. Mutat. Res 2007, 618, 18–29. [Google Scholar]
- Marfella, CG; Imbalzano, AN. The Chd family of chromatin remodelers. Mutat. Res 2007, 618, 30–40. [Google Scholar]
- Hargreaves, DC; Crabtree, GR. ATP-dependent chromatin remodeling: genetics, genomics and mechanisms. Cell Res 2011, 21, 396–420. [Google Scholar]
- Yuan, GC; Liu, YJ; Dion, MF; Slack, MD; Wu, LF; Altschuler, SJ; Rando, OJ. Genome-scale identification of nucleosome positions in S. cerevisiae. Science 2005, 309, 626–630. [Google Scholar]
- Mavrich, TN; Ioshikhes, IP; Venters, BJ; Jiang, C; Tomsho, LP; Qi, J; Schuster, SC; Albert, I; Pugh, BF. A barrier nucleosome model for statistical positioning of nucleosomes throughout the yeast genome. Genome Res 2008, 18, 1073–1083. [Google Scholar]
- Mavrich, TN; Jiang, C; Ioshikhes, IP; Li, X; Venters, BJ; Zanton, SJ; Tomsho, LP; Qi, J; Glaser, RL; Schuster, SC; et al. Nucleosome organization in the Drosophila genome. Nature 2008, 453, 358–362. [Google Scholar]
- Valouev, A; Ichikawa, J; Tonthat, T; Stuart, J; Ranade, S; Peckham, H; Zeng, K; Malek, JA; Costa, G; McKernan, K; et al. A high-resolution, nucleosome position map of C. elegans reveals a lack of universal sequence-dictated positioning. Genome Res 2008, 18, 1051–1063. [Google Scholar]
- Schones, DE; Cui, K; Cuddapah, S; Roh, TY; Barski, A; Wang, Z; Wei, G; Zhao, K. Dynamic regulation of nucleosome positioning in the human genome. Cell 2008, 132, 887–898. [Google Scholar]
- Lantermann, AB; Straub, T; Stralfors, A; Yuan, GC; Ekwall, K; Korber, P. Schizosaccharomyces pombe genome-wide nucleosome mapping reveals positioning mechanisms distinct from those of Saccharomyces cerevisiae. Nat. Struct. Mol. Biol 2010, 17, 251–257. [Google Scholar]
- Zhang, Z; Wippo, CJ; Wal, M; Ward, E; Korber, P; Pugh, BF. A packing mechanism for nucleosome organization reconstituted across a eukaryotic genome. Science 2011, 332, 977–980. [Google Scholar]
- Flaus, A; Owen-Hughes, T. Mechanisms for ATP-dependent chromatin remodelling: the means to the end. FEBS J 2011, 278, 3579–3595. [Google Scholar]
- Langst, G; Becker, PB. Nucleosome remodeling: one mechanism, many phenomena? Biochim. Biophys. Acta 2004, 1677, 58–63. [Google Scholar]
- Morettini, S; Podhraski, V; Lusser, A. ATP-dependent chromatin remodeling enzymes and their various roles in cell cycle control. Front. Biosci 2008, 13, 5522–5532. [Google Scholar]
- Moshkin, YM; Mohrmann, L; van Ijcken, WF; Verrijzer, CP. Functional differentiation of SWI/SNF remodelers in transcription and cell cycle control. Mol. Cell. Biol 2007, 27, 651–661. [Google Scholar]
- Guschin, D; Wade, PA; Kikyo, N; Wolffe, AP. ATP-Dependent histone octamer mobilization and histone deacetylation mediated by the Mi-2 chromatin remodeling complex. Biochemistry (Mosc. ) 2000, 39, 5238–5245. [Google Scholar]
- Brehm, A; Langst, G; Kehle, J; Clapier, CR; Imhof, A; Eberharter, A; Muller, J; Becker, PB. dMi-2 and ISWI chromatin remodelling factors have distinct nucleosome binding and mobilization properties. EMBO J 2000, 19, 4332–4341. [Google Scholar]
- Stockdale, C; Flaus, A; Ferreira, H; Owen-Hughes, T. Analysis of nucleosome repositioning by yeast ISWI and Chd1 chromatin remodeling complexes. J. Biol. Chem 2006, 281, 16279–16288. [Google Scholar]
- Langst, G; Bonte, EJ; Corona, DF; Becker, PB. Nucleosome movement by CHRAC and ISWI without disruption or trans- displacement of the histone octamer. Cell 1999, 97, 843–852. [Google Scholar]
- Hamiche, A; Sandaltzopoulos, R; Gdula, DA; Wu, C. ATP-dependent histone octamer sliding mediated by the chromatin remodeling complex NURF. Cell 1999, 97, 833–842. [Google Scholar]
- Rippe, K; Schrader, A; Riede, P; Strohner, R; Lehmann, E; Langst, G. DNA sequence- and conformation-directed positioning of nucleosomes by chromatin-remodeling complexes. Proc. Natl. Acad. Sci. USA 2007, 104, 15635–15640. [Google Scholar]
- Stokes, DG; Tartof, KD; Perry, RP. CHD1 is concentrated in interbands and puffed regions of Drosophila polytene chromosomes. Proc. Natl. Acad. Sci. USA 1996, 93, 7137–7142. [Google Scholar]
- Srinivasan, S; Armstrong, JA; Deuring, R; Dahlsveen, IK; McNeill, H; Tamkun, JW. The Drosophila trithorax group protein Kismet facilitates an early step in transcriptional elongation by RNA Polymerase II. Development 2005, 132, 1623–1635. [Google Scholar]
- Whitehouse, I; Tsukiyama, T. Antagonistic forces that position nucleosomes in vivo. Nat. Struct. Mol. Biol 2006, 13, 633–640. [Google Scholar]
- Sala, A; Toto, M; Pinello, L; Gabriele, A; Di Benedetto, V; Ingrassia, AM; Lo Bosco, G; Di Gesu, V; Giancarlo, R; Corona, DF. Genome-wide characterization of chromatin binding and nucleosome spacing activity of the nucleosome remodelling ATPase ISWI. EMBO J 2011, 30, 1766–1777. [Google Scholar]
- Petesch, SJ; Lis, JT. Rapid, transcription-independent loss of nucleosomes over a large chromatin domain at Hsp70 loci. Cell 2008, 134, 74–84. [Google Scholar]
- Morettini, S; Tribus, M; Zeilner, A; Sebald, J; Campo-Fernandez, B; Scheran, G; Worle, H; Podhraski, V; Fyodorov, DV; Lusser, A. The chromodomains of CHD1 are critical for enzymatic activity but less important for chromatin localization. Nucleic Acids Res 2011, 39, 3103–3115. [Google Scholar]
- Alen, C; Kent, NA; Jones, HS; O’Sullivan, J; Aranda, A; Proudfoot, NJ. A role for chromatin remodeling in transcriptional termination by RNA polymerase II. Mol. Cell 2002, 10, 1441–1452. [Google Scholar]
- Sugiyama, T; Cam, HP; Sugiyama, R; Noma, K; Zofall, M; Kobayashi, R; Grewal, SI. SHREC, an effector complex for heterochromatic transcriptional silencing. Cell 2007, 128, 491–504. [Google Scholar]
- Kwon, H; Imbalzano, AN; Khavari, PA; Kingston, RE; Green, MR. Nucleosome disruption and enhancement of activator binding by a human SW1/SNF complex. Nature 1994, 370, 477–481. [Google Scholar]
- Cairns, BR; Lorch, Y; Li, Y; Zhang, M; Lacomis, L; Erdjument-Bromage, H; Tempst, P; Du, J; Laurent, B; Kornberg, RD. RSC, an essential, abundant chromatin-remodeling complex. Cell 1996, 87, 1249–1260. [Google Scholar]
- Cote, J; Quinn, J; Workman, JL; Peterson, CL. Stimulation of GAL4 derivative binding to nucleosomal DNA by the yeast SWI/SNF complex. Science 1994, 265, 53–60. [Google Scholar]
- Imbalzano, AN; Schnitzler, GR; Kingston, RE. Nucleosome disruption by human SWI/SNF is maintained in the absence of continued ATP hydrolysis. J. Biol. Chem 1996, 271, 20726–20733. [Google Scholar]
- Lorch, Y; Zhang, M; Kornberg, RD. RSC unravels the nucleosome. Mol. Cell 2001, 7, 89–95. [Google Scholar]
- Owen-Hughes, T; Utley, RT; Cote, J; Peterson, CL; Workman, JL. Persistent site-specific remodeling of a nucleosome array by transient action of the SWI/SNF complex. Science 1996, 273, 513–516. [Google Scholar]
- Dechassa, ML; Sabri, A; Pondugula, S; Kassabov, SR; Chatterjee, N; Kladde, MP; Bartholomew, B. SWI/SNF has intrinsic nucleosome disassembly activity that is dependent on adjacent nucleosomes. Mol. Cell 2010, 38, 590–602. [Google Scholar]
- Whitehouse, I; Flaus, A; Cairns, BR; White, MF; Workman, JL; Owen-Hughes, T. Nucleosome mobilization catalysed by the yeast SWI/SNF complex. Nature 1999, 400, 784–787. [Google Scholar]
- Hartley, PD; Madhani, HD. Mechanisms that specify promoter nucleosome location and identity. Cell 2009, 137, 445–458. [Google Scholar]
- Ito, T; Levenstein, ME; Fyodorov, DV; Kutach, AK; Kobayashi, R; Kadonaga, JT. ACF consists of two subunits, Acf1 and ISWI, that function cooperatively in the ATP-dependent catalysis of chromatin assembly. Genes Dev 1999, 13, 1529–1539. [Google Scholar]
- Phelan, ML; Sif, S; Narlikar, GJ; Kingston, RE. Reconstitution of a core chromatin remodeling complex from SWI/SNF subunits. Mol. Cell 1999, 3, 247–253. [Google Scholar]
- Groth, A; Rocha, W; Verreault, A; Almouzni, G. Chromatin challenges during DNA replication and repair. Cell 2007, 128, 721–733. [Google Scholar]
- Ransom, M; Dennehey, BK; Tyler, JK. Chaperoning histones during DNA replication and repair. Cell 2010, 140, 183–195. [Google Scholar]
- Lusser, A; Kadonaga, JT. Strategies for the reconstitution of chromatin. Nat. Methods 2004, 1, 19–26. [Google Scholar]
- LeRoy, G; Loyola, A; Lane, WS; Reinberg, D. Purification and characterization of a human factor that assembles and remodels chromatin. J. Biol. Chem 2000, 275, 14787–14790. [Google Scholar]
- Lusser, A; Urwin, DL; Kadonaga, JT. Distinct activities of CHD1 and ACF in ATP-dependent chromatin assembly. Nat. Struct. Mol. Biol 2005, 12, 160–166. [Google Scholar]
- Torigoe, SE; Urwin, DL; Ishii, H; Smith, DE; Kadonaga, JT. Identification of a Rapidly Formed Nonnucleosomal Histone-DNA Intermediate that Is Converted into Chromatin by ACF. Mol. Cell 2011, 43, 638–648. [Google Scholar]
- Robinson, KM; Schultz, MC. Replication-independent assembly of nucleosome arrays in a novel yeast chromatin reconstitution system involves antisilencing factor Asf1p and chromodomain protein Chd1p. Mol. Cell. Biol 2003, 23, 7937–7946. [Google Scholar]
- Fyodorov, DV; Blower, MD; Karpen, GH; Kadonaga, JT. Acf1 confers unique activities to ACF/CHRAC and promotes the formation rather than disruption of chromatin in vivo. Genes Dev 2004, 18, 170–183. [Google Scholar]
- Poot, RA; Bozhenok, L; van den Berg, DL; Steffensen, S; Ferreira, F; Grimaldi, M; Gilbert, N; Ferreira, J; Varga-Weisz, PD. The Williams syndrome transcription factor interacts with PCNA to target chromatin remodelling by ISWI to replication foci. Nat. Cell Biol 2004, 6, 1236–1244. [Google Scholar]
- Collins, N; Poot, RA; Kukimoto, I; Garcia-Jimenez, C; Dellaire, G; Varga-Weisz, PD. An ACF1-ISWI chromatin-remodeling complex is required for DNA replication through heterochromatin. Nat. Genet 2002, 32, 627–632. [Google Scholar]
- Bozhenok, L; Wade, PA; Varga-Weisz, P. WSTF-ISWI chromatin remodeling complex targets heterochromatic replication foci. EMBO J 2002, 21, 2231–2241. [Google Scholar]
- Rowbotham, SP; Barki, L; Neves-Costa, A; Santos, F; Dean, W; Hawkes, N; Choudhary, P; Will, WR; Webster, J; Oxley, D; et al. Maintenance of silent chromatin through replication requires SWI/SNF-like chromatin remodeler SMARCAD1. Mol. Cell 2011, 42, 285–296. [Google Scholar]
- Thoma, F; Koller, T; Klug, A. Involvement of histone H1 in the organization of the nucleosome and of the salt-dependent superstructures of chromatin. J. Cell Biol 1979, 83, 403–427. [Google Scholar]
- Marsden, MP; Laemmli, UK. Metaphase chromosome structure: evidence for a radial loop model. Cell 1979, 17, 849–858. [Google Scholar]
- Routh, A; Sandin, S; Rhodes, D. Nucleosome repeat length and linker histone stoichiometry determine chromatin fiber structure. Proc. Natl. Acad. Sci. USA 2008, 105, 8872–8877. [Google Scholar]
- Robinson, PJ; Rhodes, D. Structure of the “30 nm” chromatin fibre: a key role for the linker histone. Curr. Opin. Struct. Biol 2006, 16, 336–343. [Google Scholar]
- Robinson, PJ; Fairall, L; Huynh, VA; Rhodes, D. EM measurements define the dimensions of the “30-nm” chromatin fiber: evidence for a compact, interdigitated structure. Proc. Natl. Acad. Sci. USA 2006, 103, 6506–6511. [Google Scholar]
- Schalch, T; Duda, S; Sargent, DF; Richmond, TJ. X-ray structure of a tetranucleosome and its implications for the chromatin fibre. Nature 2005, 436, 138–141. [Google Scholar]
- Woodcock, CL; Horowitz, RA. Chromatin organization re-viewed. Trends Cell Biol 1995, 5, 272–277. [Google Scholar]
- Horowitz, RA; Agard, DA; Sedat, JW; Woodcock, CL. The three-dimensional architecture of chromatin in situ: electron tomography reveals fibers composed of a continuously variable zig-zag nucleosomal ribbon. J. Cell Biol 1994, 125, 1–10. [Google Scholar]
- Kimura, H; Cook, PR. Kinetics of core histones in living human cells: little exchange of H3 and H4 and some rapid exchange of H2B. J. Cell Biol 2001, 153, 1341–1353. [Google Scholar]
- Misteli, T; Gunjan, A; Hock, R; Bustin, M; Brown, DT. Dynamic binding of histone H1 to chromatin in living cells. Nature 2000, 408, 877–881. [Google Scholar]
- Lever, MA; Th’ng, JP; Sun, X; Hendzel, MJ. Rapid exchange of histone H1.1 on chromatin in living human cells. Nature 2000, 408, 873–876. [Google Scholar]
- Siriaco, G; Deuring, R; Chioda, M; Becker, PB; Tamkun, JW. Drosophila ISWI regulates the association of histone H1 with interphase chromosomes in vivo. Genetics 2009, 182, 661–669. [Google Scholar]
- Fyodorov, DV; Kadonaga, JT. Chromatin assembly in vitro with purified recombinant ACF and NAP-1. Methods Enzymol 2003, 371, 499–515. [Google Scholar]
- Maier, VK; Chioda, M; Rhodes, D; Becker, PB. ACF catalyses chromatosome movements in chromatin fibres. EMBO J 2008, 27, 817–826. [Google Scholar]
- Deuring, R; Fanti, L; Armstrong, JA; Sarte, M; Papoulas, O; Prestel, M; Daubresse, G; Verardo, M; Moseley, SL; Berloco, M; et al. The ISWI chromatin-remodeling protein is required for gene expression and the maintenance of higher order chromatin structure in vivo. Mol. Cell 2000, 5, 355–365. [Google Scholar]
- Corona, DF; Siriaco, G; Armstrong, JA; Snarskaya, N; McClymont, SA; Scott, MP; Tamkun, JW. ISWI regulates higher-order chromatin structure and histone H1 assembly in vivo. PLoS Biol 2007, 5, e232. [Google Scholar]
- Badenhorst, P; Voas, M; Rebay, I; Wu, C. Biological functions of the ISWI chromatin remodeling complex NURF. Genes Dev 2002, 16, 3186–3198. [Google Scholar]
- Bai, X; Larschan, E; Kwon, SY; Badenhorst, P; Kuroda, MI. Regional control of chromatin organization by noncoding roX RNAs and the NURF remodeling complex in Drosophila melanogaster. Genetics 2007, 176, 1491–1499. [Google Scholar]
- Fan, Y; Nikitina, T; Morin-Kensicki, EM; Zhao, J; Magnuson, TR; Woodcock, CL; Skoultchi, AI. H1 linker histones are essential for mouse development and affect nucleosome spacing in vivo. Mol. Cell. Biol 2003, 23, 4559–4572. [Google Scholar]
- Lu, X; Wontakal, SN; Emelyanov, AV; Morcillo, P; Konev, AY; Fyodorov, DV; Skoultchi, AI. Linker histone H1 is essential for Drosophila development, the establishment of pericentric heterochromatin, and a normal polytene chromosome structure. Genes Dev 2009, 23, 452–465. [Google Scholar]
- Gunjan, A; Paik, J; Verreault, A. Regulation of histone synthesis and nucleosome assembly. Biochimie 2005, 87, 625–635. [Google Scholar]
- Henikoff, S; Ahmad, K. Assembly of variant histones into chromatin. Annu. Rev. Cell Dev. Biol 2005, 21, 133–153. [Google Scholar]
- Pina, B; Suau, P. Changes in histones H2A and H3 variant composition in differentiating and mature rat brain cortical neurons. Dev. Biol 1987, 123, 51–58. [Google Scholar]
- Ahmad, K; Henikoff, S. Histone H3 variants specify modes of chromatin assembly. Proc. Natl. Acad. Sci. USA 2002, 99, 16477–16484. [Google Scholar]
- Ahmad, K; Henikoff, S. The histone variant H3.3 marks active chromatin by replication-independent nucleosome assembly. Mol. Cell 2002, 9, 1191–1200. [Google Scholar]
- Bernstein, E; Hake, SB. The nucleosome: a little variation goes a long way. Biochem. Cell Biol 2006, 84, 505–517. [Google Scholar]
- Talbert, PB; Henikoff, S. Histone variants–ancient wrap artists of the epigenome. Nat. Rev. Mol. Cell Biol 2010, 11, 264–275. [Google Scholar]
- Wiedemann, SM; Mildner, SN; Bonisch, C; Israel, L; Maiser, A; Matheisl, S; Straub, T; Merkl, R; Leonhardt, H; Kremmer, E; et al. Identification and characterization of two novel primate-specific histone H3 variants, H3.X and H3.Y. J. Cell Biol 2010, 190, 777–791. [Google Scholar]
- Haushalter, KA; Kadonaga, JT. Chromatin assembly by DNA-translocating motors. Nat. Rev. Mol. Cell Biol 2003, 4, 613–620. [Google Scholar]
- Mellone, BG; Zhang, W; Karpen, GH. Frodos found: Behold the CENP-a “Ring” bearers. Cell 2009, 137, 409–412. [Google Scholar]
- Polo, SE; Almouzni, G. Chromatin assembly: a basic recipe with various flavours. Curr. Opin. Genet. Dev 2006, 16, 104–111. [Google Scholar]
- Loyola, A; Almouzni, G. Marking histone H3 variants: how, when and why? Trends Biochem. Sci 2007, 32, 425–433. [Google Scholar]
- Tachiwana, H; Osakabe, A; Shiga, T; Miya, Y; Kimura, H; Kagawa, W; Kurumizaka, H. Structures of human nucleosomes containing major histone H3 variants. Acta Crystallogr. D Biol.Crystallogr 2011, 67, 578–583. [Google Scholar]
- Jin, C; Felsenfeld, G. Nucleosome stability mediated by histone variants H3.3 and H2A.Z. Genes Dev 2007, 21, 1519–1529. [Google Scholar]
- Goldberg, AD; Banaszynski, LA; Noh, KM; Lewis, PW; Elsaesser, SJ; Stadler, S; Dewell, S; Law, M; Guo, X; Li, X; et al. Distinct factors control histone variant H3.3 localization at specific genomic regions. Cell 2010, 140, 678–691. [Google Scholar]
- Jin, C; Zang, C; Wei, G; Cui, K; Peng, W; Zhao, K; Felsenfeld, G. H3.3/H2A.Z double variant-containing nucleosomes mark “nucleosome-free regions” of active promoters and other regulatory regions. Nat. Genet 2009, 41, 941–945. [Google Scholar]
- Mito, Y; Henikoff, JG; Henikoff, S. Genome-scale profiling of histone H3.3 replacement patterns. Nat. Genet 2005, 37, 1090–1097. [Google Scholar]
- Sakai, A; Schwartz, BE; Goldstein, S; Ahmad, K. Transcriptional and developmental functions of the H3.3 histone variant in Drosophila. Curr. Biol 2009, 19, 1816–1820. [Google Scholar]
- Hodl, M; Basler, K. Transcription in the absence of histone H3.3. Curr. Biol 2009, 19, 1221–1226. [Google Scholar]
- Loppin, B; Bonnefoy, E; Anselme, C; Laurencon, A; Karr, TL; Couble, P. The histone H3.3 chaperone HIRA is essential for chromatin assembly in the male pronucleus. Nature 2005, 437, 1386–1390. [Google Scholar]
- van der Heijden, GW; Dieker, JW; Derijck, AA; Muller, S; Berden, JH; Braat, DD; van der Vlag, J; de Boer, P. Asymmetry in histone H3 variants and lysine methylation between paternal and maternal chromatin of the early mouse zygote. Mech. Dev 2005, 122, 1008–1022. [Google Scholar]
- Bonnefoy, E; Orsi, GA; Couble, P; Loppin, B. The essential role of Drosophila HIRA for de novo assembly of paternal chromatin at fertilization. PLoS Genet 2007, 3, 1991–2006. [Google Scholar]
- Konev, AY; Tribus, M; Park, SY; Podhraski, V; Lim, CY; Emelyanov, AV; Vershilova, E; Pirrotta, V; Kadonaga, JT; Lusser, A; et al. CHD1 motor protein is required for deposition of histone variant H3.3 into chromatin in vivo. Science 2007, 317, 1087–1090. [Google Scholar]
- Gaspar-Maia, A; Alajem, A; Polesso, F; Sridharan, R; Mason, MJ; Heidersbach, A; Ramalho-Santos, J; McManus, MT; Plath, K; Meshorer, E; et al. Chd1 regulates open chromatin and pluripotency of embryonic stem cells. Nature 2009, 460, 863–868. [Google Scholar]
- Wong, LH; McGhie, JD; Sim, M; Anderson, MA; Ahn, S; Hannan, RD; George, AJ; Morgan, KA; Mann, JR; Choo, KH. ATRX interacts with H3.3 in maintaining telomere structural integrity in pluripotent embryonic stem cells. Genome Res 2010, 20, 351–360. [Google Scholar]
- Lewis, PW; Elsaesser, SJ; Noh, KM; Stadler, SC; Allis, CD. Daxx is an H3.3-specific histone chaperone and cooperates with ATRX in replication-independent chromatin assembly at telomeres. Proc. Natl. Acad. Sci. USA 2010, 107, 14075–14080. [Google Scholar]
- Drane, P; Ouararhni, K; Depaux, A; Shuaib, M; Hamiche, A. The death-associated protein DAXX is a novel histone chaperone involved in the replication-independent deposition of H3.3. Genes Dev 2010, 24, 1253–1265. [Google Scholar]
- Torras-Llort, M; Moreno-Moreno, O; Azorin, F. Focus on the centre: the role of chromatin on the regulation of centromere identity and function. EMBO J 2009, 28, 2337–2348. [Google Scholar]
- Dalal, Y; Furuyama, T; Vermaak, D; Henikoff, S. Structure, dynamics, and evolution of centromeric nucleosomes. Proc. Natl. Acad. Sci. USA 2007, 104, 15974–15981. [Google Scholar]
- Sullivan, BA; Blower, MD; Karpen, GH. Determining centromere identity: cyclical stories and forking paths. Nat. Rev. Genet 2001, 2, 584–596. [Google Scholar]
- Allshire, RC; Karpen, GH. Epigenetic regulation of centromeric chromatin: old dogs, new tricks? Nat. Rev. Genet 2008, 9, 923–937. [Google Scholar]
- Gkikopoulos, T; Singh, V; Tsui, K; Awad, S; Renshaw, MJ; Scholfield, P; Barton, GJ; Nislow, C; Tanaka, TU; Owen-Hughes, T. The SWI/SNF complex acts to constrain distribution of the centromeric histone variant Cse4. EMBO J 2011, 30, 1919–1927. [Google Scholar]
- Dunleavy, EM; Roche, D; Tagami, H; Lacoste, N; Ray-Gallet, D; Nakamura, Y; Daigo, Y; Nakatani, Y; Almouzni-Pettinotti, G. HJURP is a cell-cycle-dependent maintenance and deposition factor of CENP-A at centromeres. Cell 2009, 137, 485–497. [Google Scholar]
- Foltz, DR; Jansen, LE; Bailey, AO; Yates, JR, 3rd; Bassett, EA; Wood, S; Black, BE; Cleveland, DW. Centromere-specific assembly of CENP-a nucleosomes is mediated by HJURP. Cell 2009, 137, 472–484. [Google Scholar]
- Williams, JS; Hayashi, T; Yanagida, M; Russell, P. Fission yeast Scm3 mediates stable assembly of Cnp1/CENP-A into centromeric chromatin. Mol. Cell 2009, 33, 287–298. [Google Scholar]
- Pidoux, AL; Choi, ES; Abbott, JK; Liu, X; Kagansky, A; Castillo, AG; Hamilton, GL; Richardson, W; Rappsilber, J; He, X; et al. Fission yeast Scm3: A CENP-A receptor required for integrity of subkinetochore chromatin. Mol. Cell 2009, 33, 299–311. [Google Scholar]
- Sanchez-Pulido, L; Pidoux, AL; Ponting, CP; Allshire, RC. Common ancestry of the CENP-A chaperones Scm3 and HJURP. Cell 2009, 137, 1173–1174. [Google Scholar]
- Perpelescu, M; Nozaki, N; Obuse, C; Yang, H; Yoda, K. Active establishment of centromeric CENP-A chromatin by RSF complex. J. Cell Biol 2009, 185, 397–407. [Google Scholar]
- Okada, M; Okawa, K; Isobe, T; Fukagawa, T. CENP-H-containing complex facilitates centromere deposition of CENP-A in cooperation with FACT and CHD1. Mol. Biol. Cell 2009, 20, 3986–3995. [Google Scholar]
- Walfridsson, J; Bjerling, P; Thalen, M; Yoo, EJ; Park, SD; Ekwall, K. The CHD remodeling factor Hrp1 stimulates CENP-A loading to centromeres. Nucleic Acids Res 2005, 33, 2868–2879. [Google Scholar]
- Podhraski, V; Campo-Fernandez, B; Worle, H; Piatti, P; Niederegger, H; Bock, G; Fyodorov, DV; Lusser, A. CenH3/CID incorporation is not dependent on the chromatin assembly factor CHD1 in Drosophila. PLoS One 2010, 5, e10120. [Google Scholar]
- Hanai, K; Furuhashi, H; Yamamoto, T; Akasaka, K; Hirose, S. RSF governs silent chromatin formation via histone H2Av replacement. PLoS Genet 2008, 4, e1000011. [Google Scholar]
- Erhardt, S; Mellone, BG; Betts, CM; Zhang, W; Karpen, GH; Straight, AF. Genome-wide analysis reveals a cell cycle-dependent mechanism controlling centromere propagation. J. Cell Biol 2008, 183, 805–818. [Google Scholar]
- Liu, X; Bowen, J; Gorovsky, MA. Either of the major H2A genes but not an evolutionarily conserved H2A.F/Z variant of Tetrahymena thermophila can function as the sole H2A gene in the yeast Saccharomyces cerevisiae. Mol. Cell. Biol 1996, 16, 2878–2887. [Google Scholar]
- Faast, R; Thonglairoam, V; Schulz, TC; Beall, J; Wells, JR; Taylor, H; Matthaei, K; Rathjen, PD; Tremethick, DJ; Lyons, I. Histone variant H2A.Z is required for early mammalian development. Curr. Biol 2001, 11, 1183–1187. [Google Scholar]
- van Daal, A; Elgin, SC. A histone variant, H2AvD, is essential in Drosophila melanogaster. Mol. Biol. Cell 1992, 3, 593–602. [Google Scholar]
- Suto, RK; Clarkson, MJ; Tremethick, DJ; Luger, K. Crystal structure of a nucleosome core particle containing the variant histone H2A.Z. Nat. Struct. Biol 2000, 7, 1121–1124. [Google Scholar]
- Fan, JY; Rangasamy, D; Luger, K; Tremethick, DJ. H2A.Z alters the nucleosome surface to promote HP1alpha-mediated chromatin fiber folding. Mol. Cell 2004, 16, 655–661. [Google Scholar]
- Zhang, Z; Pugh, BF. Genomic Organization of H2Av Containing Nucleosomes in Drosophila Heterochromatin. PLoS One 2011, 6, e20511. [Google Scholar]
- Hardy, S; Jacques, PE; Gevry, N; Forest, A; Fortin, ME; Laflamme, L; Gaudreau, L; Robert, F. The euchromatic and heterochromatic landscapes are shaped by antagonizing effects of transcription on H2A.Z deposition. PLoS Genet 2009, 5, e1000687. [Google Scholar]
- Henikoff, S; Henikoff, JG; Sakai, A; Loeb, GB; Ahmad, K. Genome-wide profiling of salt fractions maps physical properties of chromatin. Genome Res 2009, 19, 460–469. [Google Scholar]
- Mizuguchi, G; Shen, X; Landry, J; Wu, WH; Sen, S; Wu, C. ATP-driven exchange of histone H2AZ variant catalyzed by SWR1 chromatin remodeling complex. Science 2004, 303, 343–348. [Google Scholar]
- Kobor, MS; Venkatasubrahmanyam, S; Meneghini, MD; Gin, JW; Jennings, JL; Link, AJ; Madhani, HD; Rine, J. A Protein Complex Containing the Conserved Swi2/Snf2-Related ATPase Swr1p Deposits Histone Variant H2A.Z into Euchromatin. PLoS Biol 2004, 2, e131. [Google Scholar] [Green Version]
- Krogan, NJ; Keogh, MC; Datta, N; Sawa, C; Ryan, OW; Ding, H; Haw, RA; Pootoolal, J; Tong, A; Canadien, V; et al. A Snf2 family ATPase complex required for recruitment of the histone H2A variant Htz1. Mol. Cell 2003, 12, 1565–1576. [Google Scholar]
- Ruhl, DD; Jin, J; Cai, Y; Swanson, S; Florens, L; Washburn, MP; Conaway, RC; Conaway, JW; Chrivia, JC. Purification of a human SRCAP complex that remodels chromatin by incorporating the histone variant H2A.Z into nucleosomes. Biochemistry (Mosc. ) 2006, 45, 5671–5677. [Google Scholar]
- Luk, E; Ranjan, A; Fitzgerald, PC; Mizuguchi, G; Huang, Y; Wei, D; Wu, C. Stepwise histone replacement by SWR1 requires dual activation with histone H2A.Z and canonical nucleosome. Cell 2010, 143, 725–736. [Google Scholar]
- Wu, WH; Alami, S; Luk, E; Wu, CH; Sen, S; Mizuguchi, G; Wei, D; Wu, C. Swc2 is a widely conserved H2AZ-binding module essential for ATP-dependent histone exchange. Nat. Struct. Mol. Biol 2005, 12, 1064–1071. [Google Scholar]
- Papamichos-Chronakis, M; Watanabe, S; Rando, OJ; Peterson, CL. Global regulation of H2A.Z localization by the INO80 chromatin-remodeling enzyme is essential for genome integrity. Cell 2011, 144, 200–213. [Google Scholar]
- Park, YJ; Chodaparambil, JV; Bao, Y; McBryant, SJ; Luger, K. Nucleosome assembly protein 1 exchanges histone H2A-H2B dimers and assists nucleosome sliding. J. Biol. Chem 2005, 280, 1817–1825. [Google Scholar]
- Luk, E; Vu, ND; Patteson, K; Mizuguchi, G; Wu, WH; Ranjan, A; Backus, J; Sen, S; Lewis, M; Bai, Y; et al. Chz1, a nuclear chaperone for histone H2AZ. Mol. Cell 2007, 25, 357–368. [Google Scholar]
- Straube, K; Blackwell, JS, Jr; Pemberton, LF. Nap1 and Chz1 have separate Htz1 nuclear import and assembly functions. Traffic 2010, 11, 185–197. [Google Scholar]
- Reeves, R. Nuclear functions of the HMG proteins. Biochim. Biophys. Acta 2010, 1799, 3–14. [Google Scholar]
- Bianchi, ME; Agresti, A. HMG proteins: dynamic players in gene regulation and differentiation. Curr. Opin. Genet. Dev 2005, 15, 496–506. [Google Scholar]
- Rochman, M; Malicet, C; Bustin, M. HMGN5/NSBP1: a new member of the HMGN protein family that affects chromatin structure and function. Biochim. Biophys. Acta 2010, 1799, 86–92. [Google Scholar]
- Catez, F; Yang, H; Tracey, KJ; Reeves, R; Misteli, T; Bustin, M. Network of dynamic interactions between histone H1 and high-mobility-group proteins in chromatin. Mol. Cell. Biol 2004, 24, 4321–4328. [Google Scholar]
- Rochman, M; Postnikov, Y; Correll, S; Malicet, C; Wincovitch, S; Karpova, TS; McNally, JG; Wu, X; Bubunenko, NA; Grigoryev, S; et al. The interaction of NSBP1/HMGN5 with nucleosomes in euchromatin counteracts linker histone-mediated chromatin compaction and modulates transcription. Mol. Cell 2009, 35, 642–656. [Google Scholar]
- Phair, RD; Scaffidi, P; Elbi, C; Vecerova, J; Dey, A; Ozato, K; Brown, DT; Hager, G; Bustin, M; Misteli, T. Global nature of dynamic protein-chromatin interactions in vivo: three-dimensional genome scanning and dynamic interaction networks of chromatin proteins. Mol. Cell. Biol 2004, 24, 6393–6402. [Google Scholar]
- Rattner, BP; Yusufzai, T; Kadonaga, JT. HMGN proteins act in opposition to ATP-dependent chromatin remodeling factors to restrict nucleosome mobility. Mol. Cell 2009, 34, 620–626. [Google Scholar]
- Berube, NG; Smeenk, CA; Picketts, DJ. Cell cycle-dependent phosphorylation of the ATRX protein correlates with changes in nuclear matrix and chromatin association. Hum. Mol. Genet 2000, 9, 539–547. [Google Scholar]
- Lechner, MS; Schultz, DC; Negorev, D; Maul, GG; Rauscher, FJ, 3rd. The mammalian heterochromatin protein 1 binds diverse nuclear proteins through a common motif that targets the chromoshadow domain. Biochem. Biophys. Res. Commun 2005, 331, 929–937. [Google Scholar]
- Kourmouli, N; Sun, YM; van der Sar, S; Singh, PB; Brown, JP. Epigenetic regulation of mammalian pericentric heterochromatin in vivo by HP1. Biochem. Biophys. Res. Commun 2005, 337, 901–907. [Google Scholar]
- Emelyanov, AV; Konev, AY; Vershilova, E; Fyodorov, DV. Protein complex of Drosophila ATRX/XNP and HP1a is required for the formation of pericentric beta-heterochromatin in vivo. J. Biol. Chem 2010, 285, 15027–15037. [Google Scholar]
- Eskeland, R; Eberharter, A; Imhof, A. HP1 binding to chromatin methylated at H3K9 is enhanced by auxiliary factors. Mol. Cell. Biol 2007, 27, 453–465. [Google Scholar]
- Hake, SB; Allis, CD. Histone H3 variants and their potential role in indexing mammalian genomes: the “H3 barcode hypothesis”. Proc. Natl. Acad. Sci. USA 2006, 103, 6428–6435. [Google Scholar]
- Henikoff, S; Furuyama, T; Ahmad, K. Histone variants, nucleosome assembly and epigenetic inheritance. Trends Genet 2004, 20, 320–326. [Google Scholar]


© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Piatti, P.; Zeilner, A.; Lusser, A. ATP-Dependent Chromatin Remodeling Factors and Their Roles in Affecting Nucleosome Fiber Composition. Int. J. Mol. Sci. 2011, 12, 6544-6565. https://doi.org/10.3390/ijms12106544
Piatti P, Zeilner A, Lusser A. ATP-Dependent Chromatin Remodeling Factors and Their Roles in Affecting Nucleosome Fiber Composition. International Journal of Molecular Sciences. 2011; 12(10):6544-6565. https://doi.org/10.3390/ijms12106544
Chicago/Turabian StylePiatti, Paolo, Anette Zeilner, and Alexandra Lusser. 2011. "ATP-Dependent Chromatin Remodeling Factors and Their Roles in Affecting Nucleosome Fiber Composition" International Journal of Molecular Sciences 12, no. 10: 6544-6565. https://doi.org/10.3390/ijms12106544