In Vitro Antioxidant, Anticoagulant and Antimicrobial Activity and in Inhibition of Cancer Cell Proliferation by Xylan Extracted from Corn Cobs



Abstract
:1. Introduction
2. Results and Discussion
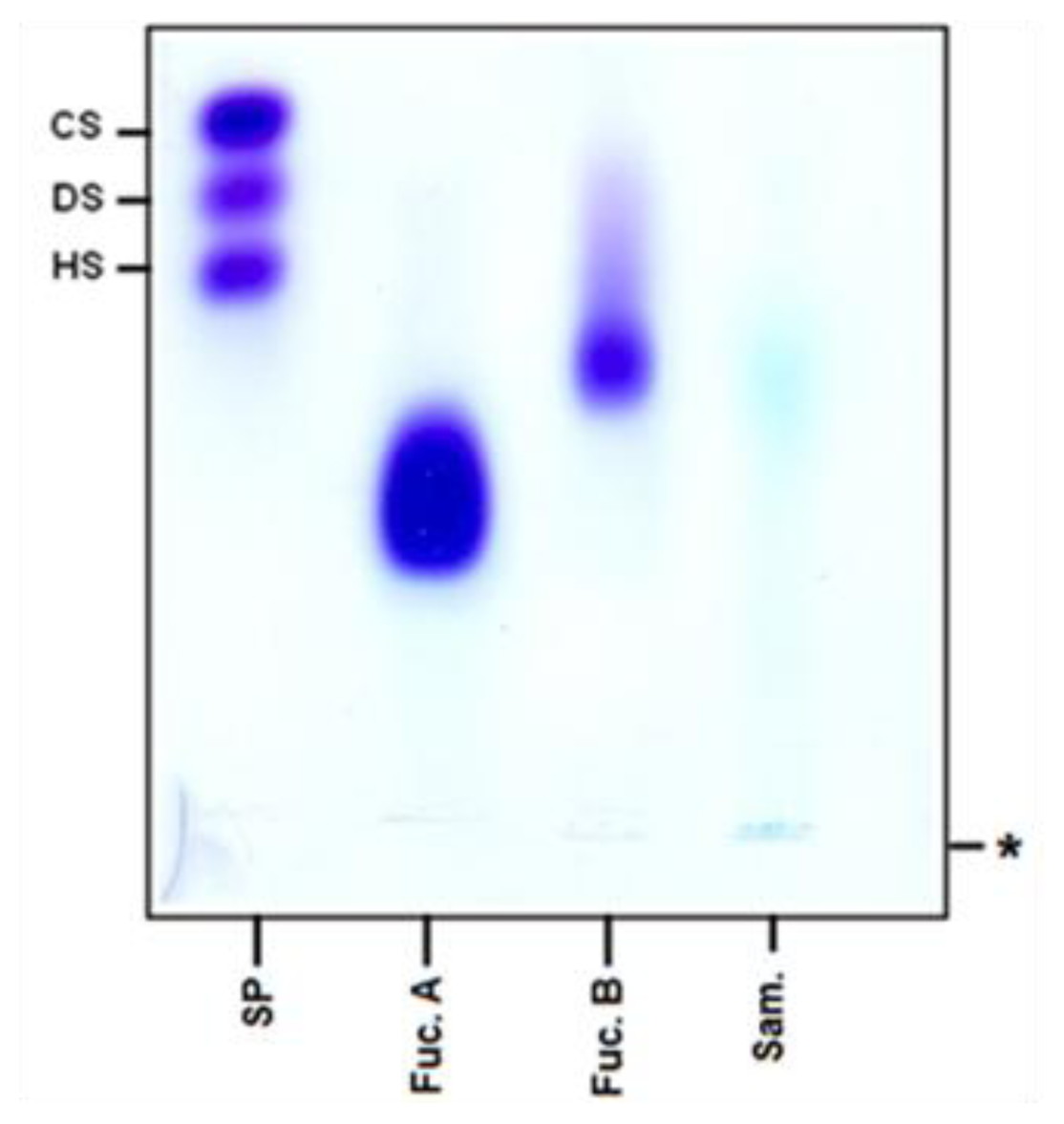
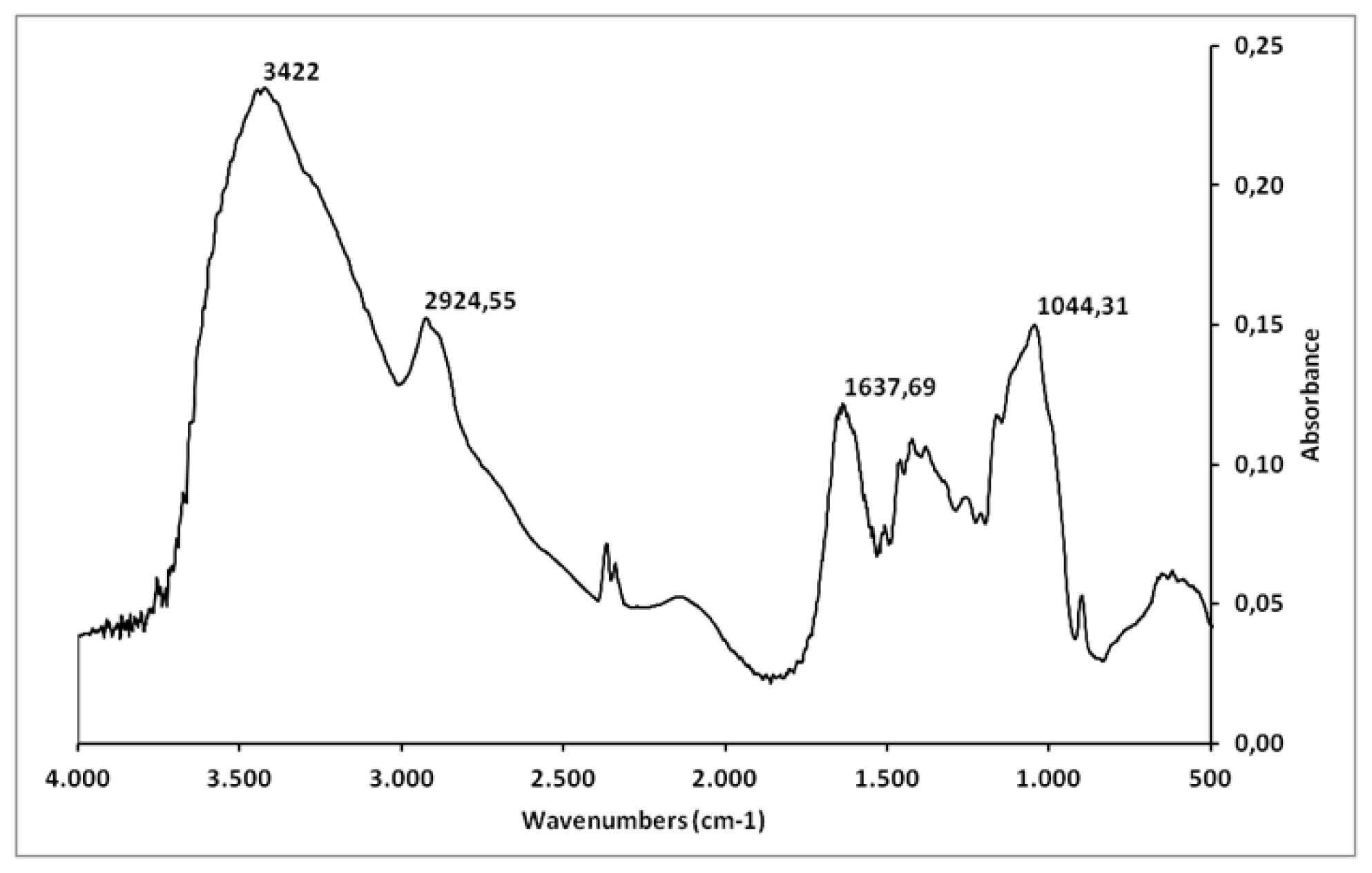
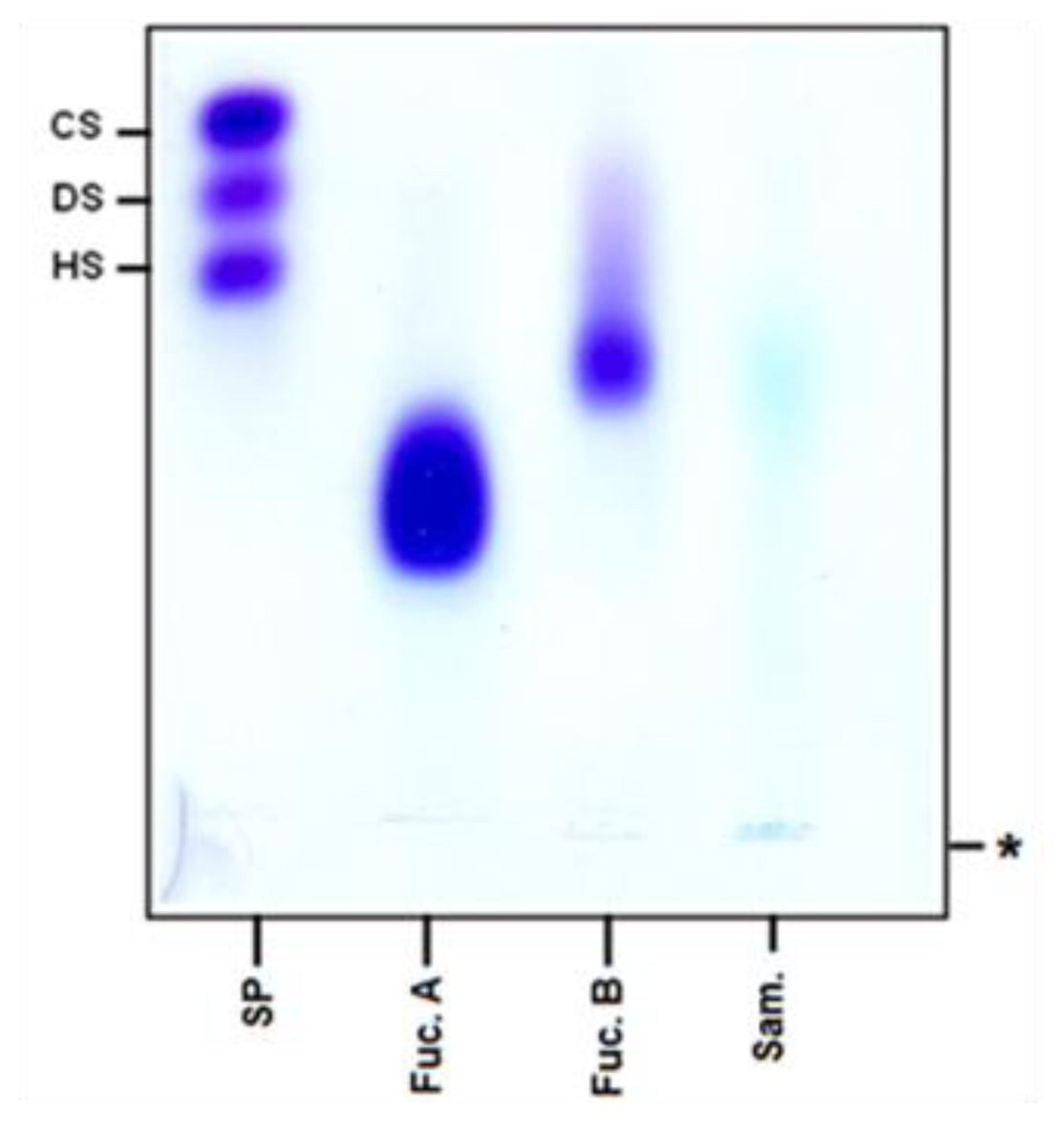
2.1. Chemical Analysis
2.2. Antioxidant Activity
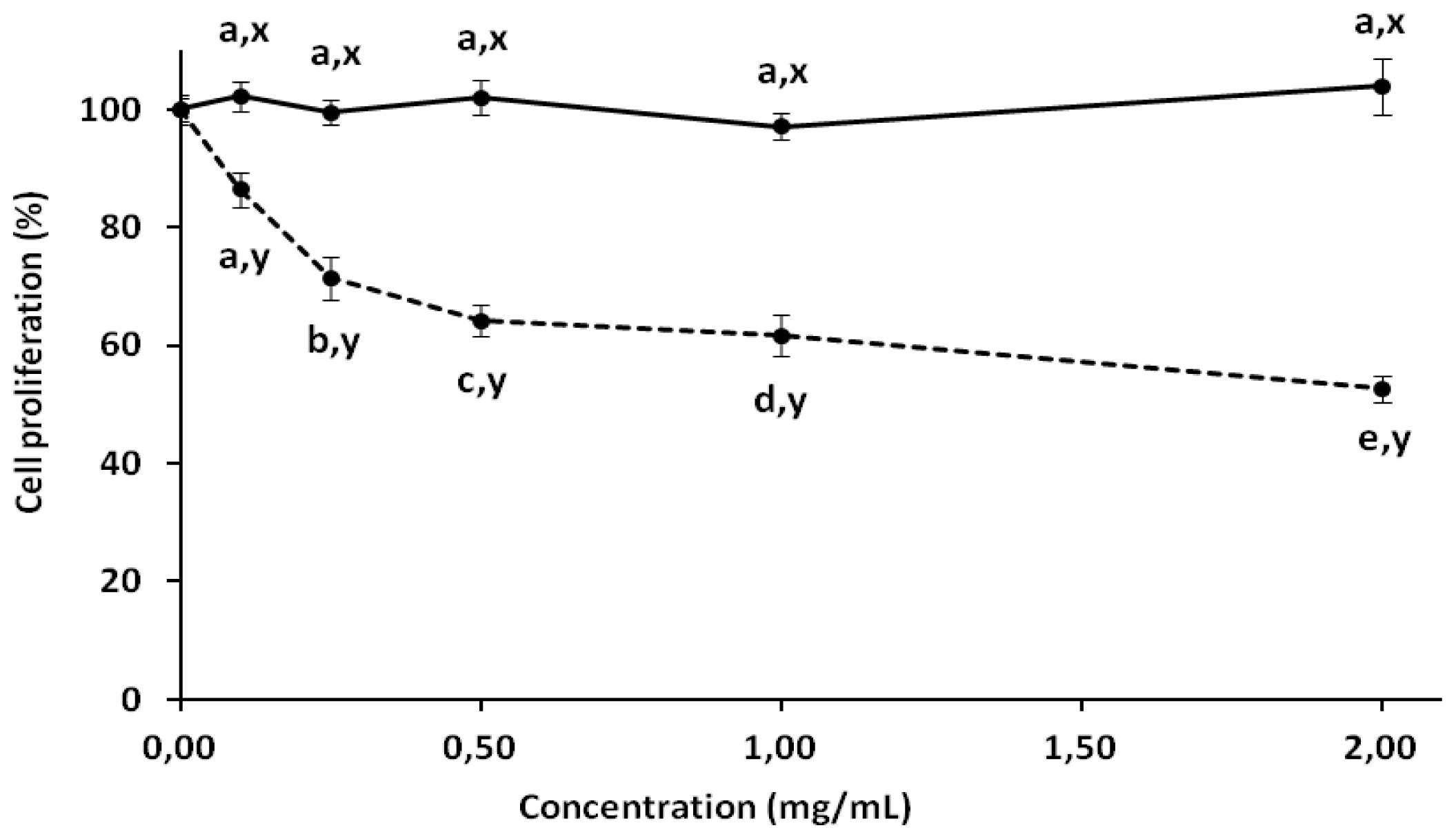
2.3. Antiproliferative Activity
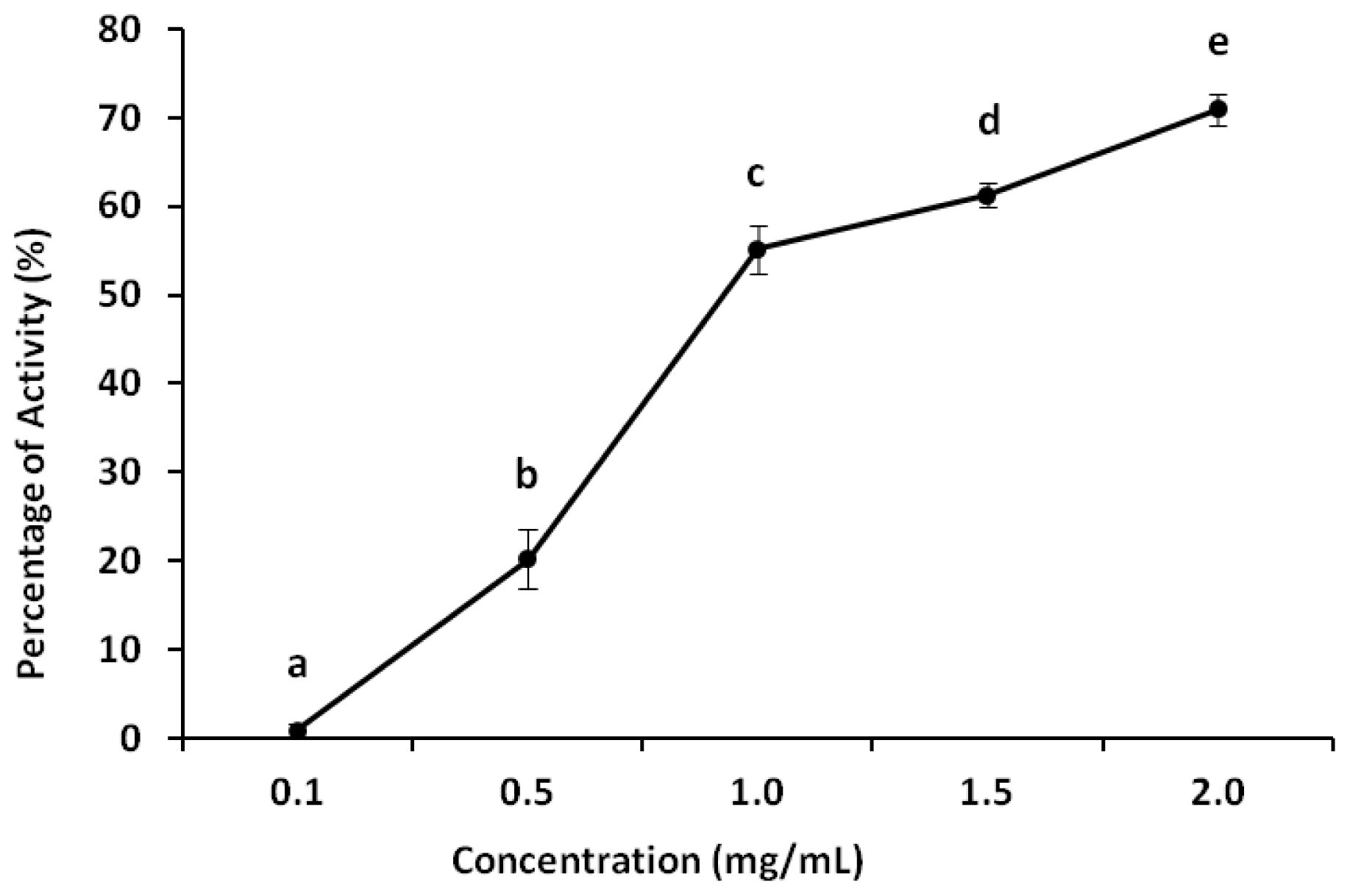
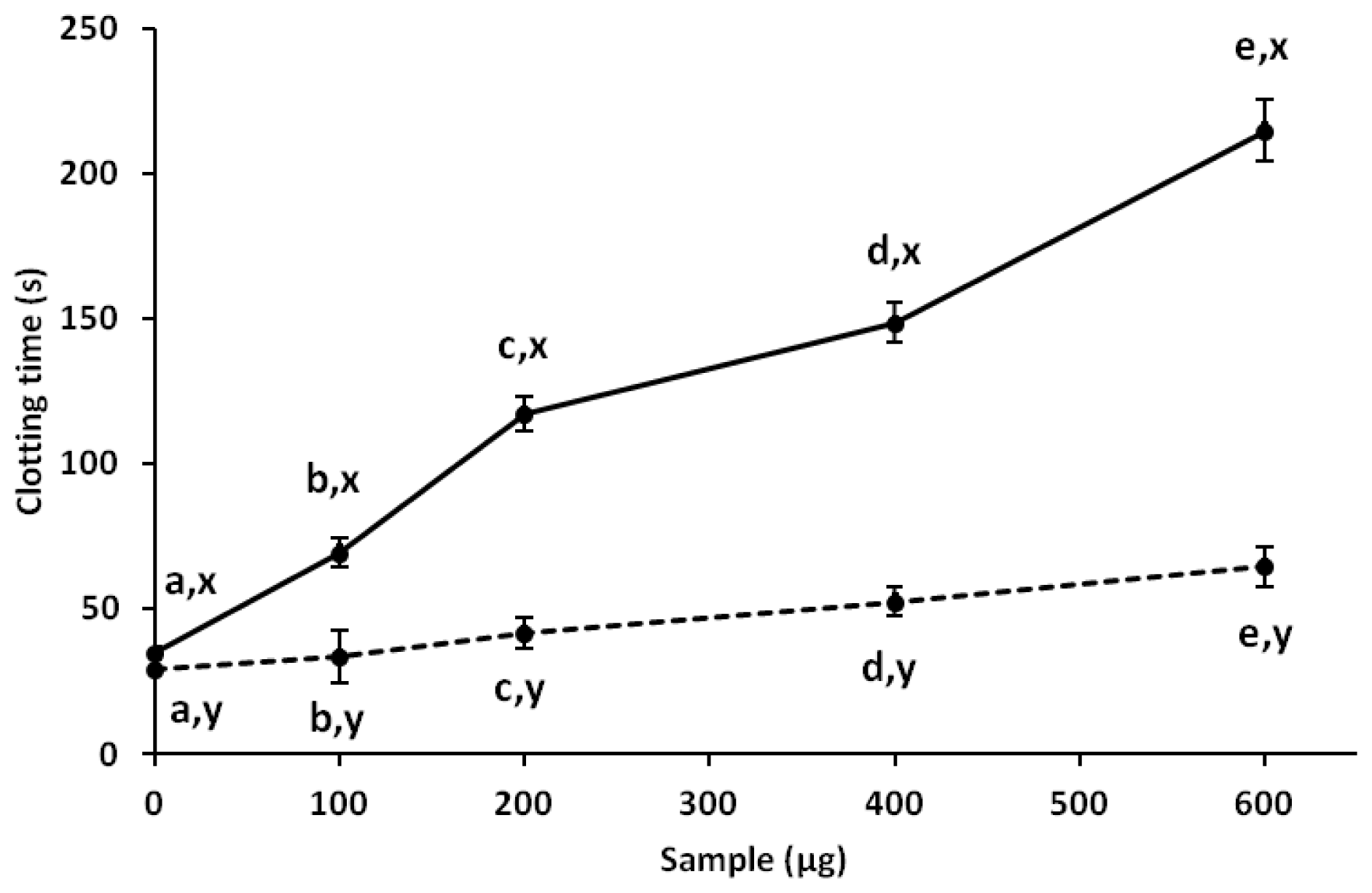
2.4. Anticoagulant Activity
2.5. Antimicrobial Activity
3. Experimental Section
3.1. Materials
3.2. Extraction of Xylan from Corn Cobs
3.2. Chemical Analysis and Monosaccharide Composition
3.3. Molecular Weight Determination
3.4. Fourier Transformed Infrared (FT-IR) Spectroscopy Analysis
3.5. Antioxidant Activities
3.5.1. Determination of Total Antioxidant Capacity
3.5.2. Hydroxyl Radical Scavenging Activity Assay
3.5.3. Superoxide Radical Scavenging Activity Assay
3.5.4. Ferric Chelating
3.5.5. Reducing Power
3.6. Antiproliferative Activity
3.7. Anticoagulant Activity
3.8. Antimicrobial Activity
3.8.1. Antibiofilm Assay
3.8.2. Bacterial Inhibition Assay
3.9. Statistical Analysis
4. Conclusion
Acknowledgments
References
- Winterbourn, C.C. Reconciling the chemistry and biology of reactive oxygen species. Nat. Chem. Biol 2008, 4, 278–286. [Google Scholar]
- Koppula, S.B.; Ammani, K. Antioxidant findings of araku environment medicinal plants using different assays. Drug Invent. Today 2011, 3, 203–205. [Google Scholar]
- Soory, M. Relevance of nutritional antioxidants in metabolic syndrome, ageing and cancer: Potential for therapeutic targeting. Infect. Disord. Drug Targets 2009, 9, 400–414. [Google Scholar]
- Balsano, C.; Alisi, A. Antioxidant effects of natural bioactive compounds. Curr. Pharm. Des 2009, 15, 3063–3073. [Google Scholar]
- Sun, J.; Chu, Y.F.; Wu, X.; Liu, R.H. Antioxidant and antiproliferative activities of common fruits. J. Agric. Food Chem 2002, 50, 7449–7454. [Google Scholar]
- Camara, R.B.; Costa, L.S.; Fidelis, G.P.; Nobre, L.T.; Dantas-Santos, N.; Cordeiro, S.L.; Costa, M.S.; Alves, L.G.; Rocha, H.A. Heterofucans from the brown seaweed Canistrocarpus cervicornis with anticoagulant and antioxidant activities. Mar. Drugs 2011, 9, 124–138. [Google Scholar]
- de Sousa, A.P.A.; Torres, M.R.; Pessoa, C.; de Moraes, M.O.; Filho, F.D.R.; Alves, A.P.N.N. In vivo growth-inhibition of Sarcoma 180 tumor by alginates from brown seaweed Sargassum vulgare. Carbohydr. Polym 2007, 69, 7–13. [Google Scholar]
- Costa, L.S.; Fidelis, G.P.; Cordeiro, S.L.; Oliveira, R.M.; Sabry, D.A.; Camara, R.B.; Nobre, L.T.; Costa, M.S.; Almeida-Lima, J.; Farias, E.H.; et al. Biological activities of sulfated polysaccharides from tropical seaweeds. Biomed. Pharmacother 2010, 64, 21–28. [Google Scholar]
- da Silva, T.D.; Giordani, R.B.; Zimmer, K.R.; da Silva, A.G.; da Silva, M.V.; Correia, M.T.; Baumvol, I.J.R.; Macedo, A.J. Potential of medicinal plants from the Brazilian semi-arid region (Caatinga) against Staphylococcus epidermidis planktonic and biofilm lifestyles. J. Ethnopharmacol 2011, 137, 327–335. [Google Scholar]
- Mehrotra, V.; Mehrotra, S.; Kirar, V.; Shyam, R.; Misra, K.; Srivastava, A.K.; Nandi, S.P. Antioxidant and antimicrobial activities of aqueous extract of Withania somnifera against methicillin-resistant Staphylococcus aureus. J. Microbiol. Biotechnol. Res 2011, 1, 40–45. [Google Scholar]
- Ebringerová, A.; Kardosová, A.; Hromádková, Z.; Malovíková, A.; Hríbalova, V. Immunomodulatory activity of acidic xylans in relation to their structural and molecular properties. Int. J. Biol. Macromol 2002, 30, 1–6. [Google Scholar]
- Anwar, F.; Jami, A.; Iqbal, S.; Sheikh, M.A. Antioxidant activity of various plant extracts under ambient and accelerated storage of sunflower oil. Grasas y Aceites 2006, 57, 189–197. [Google Scholar]
- Garcia, R.B.; Ganterb, J.L.M.S.; Carvalho, R.R. Solution properties of d-xylans from corn cobs. Eur. Polym. J 2000, 36, 783–787. [Google Scholar]
- Wang, Y.; Zhang, J. A novel hybrid process, enhanced by ultrasonication, for xylan extraction from corncobs and hydrolysis of xylan to xylose by xylanase. J. Food Eng 2006, 77, 140–145. [Google Scholar]
- Hromadková, Z.; Kovaciková, J.; Ebringerová, A. Study of the classical and ultrasound-assisted extraction of the corn cob xylan. Ind. Crops Prod 1999, 9, 101–109. [Google Scholar]
- Ebringerová, A.; Hromadková, Z.; Alfoldi, J.; Berth, G. Structural and solution properties of corn cob heteroxylans. Carbohydr. Polym 1992, 19, 99–105. [Google Scholar]
- Ebringerova, A.; Hromadkova, Z.; Alfoldi, J.; Hribalova, V. The immunologically active xylan from ultrasound-treated corn cobs: Extractability, structure and properties. Carbohydr. Polym 1998, 37, 231–239. [Google Scholar]
- Ebringerová, A.; Hromádková, Z. The effect of ultrasound on the structure and properties of the watersoluble corn hull heteroxylan. Ultrason. Sonochem 1997, 4, 305–309. [Google Scholar]
- Garcia, R.B.; Nagashima, T., Jr; Praxedes, A.K.C.; Raffin, F.N.; Moura, T.F.A.L.; do Egito, E.S.T. Preparation of micro and nanoparticles from corn cobs xylan. Polym. Bull. 2001, 46, 371–379. [Google Scholar]
- Barroso, E.M.A.; Costa, L.S.; Medeiros, V.P.; Cordeiro, L.S.; Costa, M.S.S.P.; Franco, C.R.C.; Nader, H.B.; Leite, E.L.; Rocha, H.A.O. A non-anticoagulant heterofucan has antithrombotic activity in vivo. Planta Med 2008, 74, 712–718. [Google Scholar]
- Kacuráková, M.; Wellner, N.; Ebringerová, A.; Hromádková, Z.; Wilson, R.H.; Belton, P.S. Characterisation of xylan-type polysaccharides and associated cell wall components by FT-IR and FT-Raman spectroscopies. Food Hydrocoll 1999, 13, 35–41. [Google Scholar]
- Oliveira, E.E.; Silva, A.E.; Nagashima, T.; Gomes, M.C.S.; Aguiar, L.M.; Marcelino, H.R.; Araujo, I.B.; Bayer, M.P.; Ricardo, N.M.P.S.; Oliveira, A.G.; et al. Xylan from corn cobs, a promising polymer for drug delivery production and characterization. Bioresour. Technol 2010, 101, 5402–5406. [Google Scholar]
- Prieto, P.; Pineda, M.; Aguilar, M. Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex: Specific application to the determination of vitamin E. Anal. Biochem 1999, 269, 337–341. [Google Scholar]
- Choi, J.-I.; Kim, J.-K.; Srinivasan, P.; Kim, J.-H.; Park, H.-J.; Byun, M.-W.; Lee, J.-W. Comparison of gamma ray and electron beam irradiation on extraction yield, morphological and antioxidant properties of polysaccharides from tamarind seed. Radiat. Phys. Chem 2009, 78, 605–609. [Google Scholar]
- Singh, S.; Singh, R.P. In Vitro methods of assay of antioxidants: An overview. Food Rev. Int 2008, 24, 392–415. [Google Scholar]
- Cao, Y.; Ikeda, I. Antioxidant activity and antitumor activity (in vitro) of xyloglucan selenious ester and surfated xyloglucan. Int. J. Biol. Macromol 2009, 45, 231–235. [Google Scholar]
- Wang, J.; Zhang, Q.; Zhang, Z.; Li, Z. Antioxidant activity of sulfated polysaccharide fractions extracted from Laminaria japonica. Int. J. Biol. Macromol 2008, 42, 127–132. [Google Scholar]
- Qi, H.; Zhao, T.; Zhang, Q.; Li, Z.; Zhao, Z.; Xing, R. Antioxidant activity of different molecular weight sulfated polysaccharides from Ulva pertusa Kjellm (Chlorophyta). J. Appl. Phycol 2005, 17, 527–534. [Google Scholar]
- Zhou, Y.C.; Zheng, R.L. Phenolic compounds and an analog as superoxide anion scavengers and anti oxidants. Biochem. Pharmacol 1991, 42, 1177–1179. [Google Scholar]
- Liu, W.; Wang, H.; Yao, W.; Gao, X.; Yu, L. Effects of sulfation on the physicochemical and functional properties of a water insoluble polysaccharide preparation fromGanoderma lucidum. J. Agric. Food Chem 2010, 58, 3336–3334. [Google Scholar]
- Changa, S.C.; Hsua, B.Y.; Chen, B.H. Structural characterization of polysaccharides from Zizyphus jujube and evaluation of antioxidant activity. Int. J. Biol. Macromol 2010, 47, 445–453. [Google Scholar]
- Telles, C.B.S.; Sabry, D.A.; Almeida-Lima, J.; Costa, M.S.S.P.; Melo-Silveira, R.F.; Trindade, E.S.; Sassaki, G.L.; Wisbeck, E.; Furlan, S.A.; Leite, E.L.; et al. Sulfation of the extracellular polysaccharide produced by the edible mushroom Pleurotus sajor caju alters its antioxidant, anticoagulant and antiproliferative properties in vitro. Carbohydr. Pol 2011, 85, 514–521. [Google Scholar]
- Yang, X.B.; Gao, X.D.; Han, F.; Tan, R.X. Sulfation of a polysaccharide produced by marine filamentous fungus Phoma herbarum YS4108 alters its antioxidant properties in vitro. Biochim. Biophys. Acta 2005, 1725, 120–127. [Google Scholar]
- Ryu, D.S.; Kim, S.H.; Lee, D.S. Anti-proliferative effect of polysaccharides from Salicornia herbacea on induction of G2/M arrest and apoptosis in human colon cancer cells. J. Microbiol. Biotechnol 2009, 19, 1482–1489. [Google Scholar]
- Ando, H.; Ohba, H.; Sakak, I.T.; Takamine, K.; Kamino, Y.; Moriwaki, S.; Bakalova, R.; Uemura, Y.; Hatate, Y. Hot-compressed-water decomposed products from bamboo manifest a selective cytotoxicity against acute lymphoblastic leukemia cells. Toxicol. in Vitro 2004, 18, 765–771. [Google Scholar]
- Meyers, K.J.; Watkins, C.B.; Pritts, M.P.; Rui, H.L. Antioxidant and antiproliferative activities of strawberries. J. Agric. Food Chem 2003, 51, 6887–6892. [Google Scholar]
- Yoon, S.; Pereira, M.S.; Pavao, M.S.G.; Hwang, J.; Pyun, Y.; Mourao, P.A.S. The medicinal plant Porana volubilis contains polysaccharides with anticoagulant activity mediated by heparin cofactor II. Thromb. Res 2002, 106, 51–58. [Google Scholar]
- Lee, C.K.; Kin, H.; Moon, K.H.; Shun, K.H. Screening and isolation of antibiotic resistance inhibitors from herb materials resistance inhibition of volatile components of Korean aromatic herbs. Arch. Pharm. Res 1998, 21, 62–66. [Google Scholar]
- Gul, M.Z.; Bhakshu, L.M.; Ahmad, F.; Kondapi, A.K.; Qureshi, I.A.; Ghazi, I.A. Evaluation of Abelmoschus moschatus extracts for antioxidant, free radical scavenging, antimicrobial and antiproliferative activities using in vitro assays. BMC Complement. Altern. Med 2011, 11, 64. [Google Scholar]
- Almeida-Lima, J.; Costa, L.S.; Silva, N.B.; Melo-Silveira, R.F.; Silva, F.V.; Felipe, M.B.; Medeiros, S.R.; Leite, E.L.; Rocha, H.A. Evaluating the possible genotoxic, mutagenic and tumor cell proliferation-inhibition effects of a non-anticoagulant, but antithrombotic algal heterofucan. J. Appl. Toxicol 2010, 7, 708–715. [Google Scholar]
- Rocha, H.A.; Bezerra, L.C.; de Albuquerque, I.R.; Costa, L.S.; Guerra, C.M.; de Abreu, L.D.; Nader, H.B.; Leite, E.L. A xylogalactofucan from the brown seaweed Spatoglossum schröederi stimulates the synthesis of an antithrombotic heparan sulfate from endothelial cells. Planta Med 2005, 71, 379–381. [Google Scholar]
- Magalhaes, K.D.; Costa, L.S.; Fidelis, G.P.; Oliveira, R.M.; Nobre, L.T.D.B.; Dantas-Santos, N.; Camara, R.B.G.; Albuquerque, I.R.L.; Cordeiro, S.L.; Sabry, D.A.; et al. Anticoagulant, antioxidant and antitumor activities of heterofucans from the seaweed Dictyopeteris delicatula. Int. J. Mol. Sci 2011, 12, 3352–3365. [Google Scholar]
- Dische, Z. A new specific color reactions of hexuronic acids. J. Biol. Chem 1974, 167, 189–198. [Google Scholar]
- Antunes, A.L.; Trentin, D.S.; Bonfanti, J.W.; Pinto, C.C.; Perez, L.R.; Macedo, A.J.; Barth, A.L. Application of a feasible method for determination of biofilm antimicrobial susceptibility in staphylococci. APMIS 2010, 118, 873–877. [Google Scholar]
- Antunes, A.L.; Bonfanti, J.W.; Perez, L.R.; Pinto, C.C.; Freitas, A.L.; Macedo, A.J.; Barth, A.L. High vancomycin resistance among biofilms produced by Staphylococcus species isolated from central venous catheters. Mem. Inst. Oswaldo Cruz 2011, 106, 51–55. [Google Scholar]
- Gupta, N.; Limbago, B.M.; Patel, J.B.; Kallen, A.J. Carbapenem-resistant enterobacteriaceae: Epidemiology and prevention. Clin. Infect. Dis 2011, 53, 60–67. [Google Scholar]
- Baumvol, I.J.; Macedo, A.J. Potential of medicinal plants from the Brazilian semi-arid region (Caatinga) against Staphylococcus epidermidis planktonic and biofilm lifestyles. J. Ethnopharmacol 2011, 137, 327–335. [Google Scholar]
- SigmaStat, version 2.01; Jandel Scientific Software: San Rafael, CA, USA, 1997.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Polysaccharides (%) | Phenolic Compounds (%) | Protein (%) | Molar Ratio a (%) | |||||
|---|---|---|---|---|---|---|---|---|---|
| Xyl | Ara | Glc | Gal | Man | Glu A | ||||
| Xylan | 70 | <0.01 | 0.4 | 50.0 | 15.0 | 20.0 | 10.0 | 2.5 | 2.5 |
| APTT (s) b | |||
|---|---|---|---|
| Clexane®c | Xylan | ||
| Mass Utilized (μg) | Clotting Time (s) | Mass Utilized (mg) | Clotting Time (s) |
| 0.1 | 35.2 ± 3.5 | 0.1 | 69.5 ± 5.2 |
| 0.4 | 195.6 ± 8.8 | 0.2 | 117.0 ± 3.3 |
| 1.0 | ≥240 | 0.4 | 148.5 ± 2.4 |
| 10.0 | ≥240 | 0.6 | 214.8 ± 4.0 |
| Microorganism | Xylan From Corn Cob | |
|---|---|---|
| % Antibacterial b,c | % Antibiofilm b,c | |
| KPC producing Klebsiella pneumoniae (clinical isolate) | 25.4 ± 0.02 | 30.0 ± 0.09 |
| Staphylococcus epidermidis (ATCC 35984) | 0 | 0 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Melo-Silveira, R.F.; Fidelis, G.P.; Costa, M.S.S.P.; Telles, C.B.S.; Dantas-Santos, N.; Elias, S.d.O.; Ribeiro, V.B.; Barth, A.L.; Macedo, A.J.; Leite, E.L.; et al. In Vitro Antioxidant, Anticoagulant and Antimicrobial Activity and in Inhibition of Cancer Cell Proliferation by Xylan Extracted from Corn Cobs. Int. J. Mol. Sci. 2012, 13, 409-426. https://doi.org/10.3390/ijms13010409
Melo-Silveira RF, Fidelis GP, Costa MSSP, Telles CBS, Dantas-Santos N, Elias SdO, Ribeiro VB, Barth AL, Macedo AJ, Leite EL, et al. In Vitro Antioxidant, Anticoagulant and Antimicrobial Activity and in Inhibition of Cancer Cell Proliferation by Xylan Extracted from Corn Cobs. International Journal of Molecular Sciences. 2012; 13(1):409-426. https://doi.org/10.3390/ijms13010409
Chicago/Turabian StyleMelo-Silveira, Raniere Fagundes, Gabriel Pereira Fidelis, Mariana Santana Santos Pereira Costa, Cinthia Beatrice Silva Telles, Nednaldo Dantas-Santos, Susana de Oliveira Elias, Vanessa Bley Ribeiro, Afonso Luis Barth, Alexandre José Macedo, Edda Lisboa Leite, and et al. 2012. "In Vitro Antioxidant, Anticoagulant and Antimicrobial Activity and in Inhibition of Cancer Cell Proliferation by Xylan Extracted from Corn Cobs" International Journal of Molecular Sciences 13, no. 1: 409-426. https://doi.org/10.3390/ijms13010409
APA StyleMelo-Silveira, R. F., Fidelis, G. P., Costa, M. S. S. P., Telles, C. B. S., Dantas-Santos, N., Elias, S. d. O., Ribeiro, V. B., Barth, A. L., Macedo, A. J., Leite, E. L., & Rocha, H. A. O. (2012). In Vitro Antioxidant, Anticoagulant and Antimicrobial Activity and in Inhibition of Cancer Cell Proliferation by Xylan Extracted from Corn Cobs. International Journal of Molecular Sciences, 13(1), 409-426. https://doi.org/10.3390/ijms13010409