Microsatellite Mutation Rate during Allohexaploidization of Newly Resynthesized Wheat

 and
and
Abstract
:1. Introduction
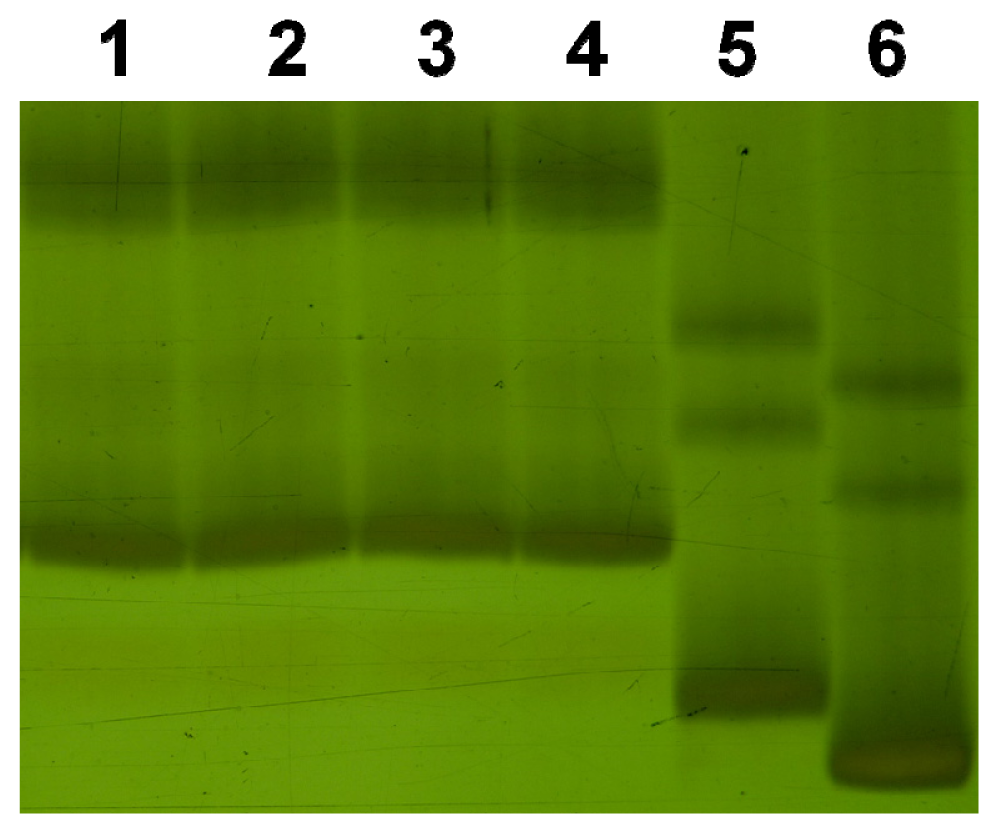
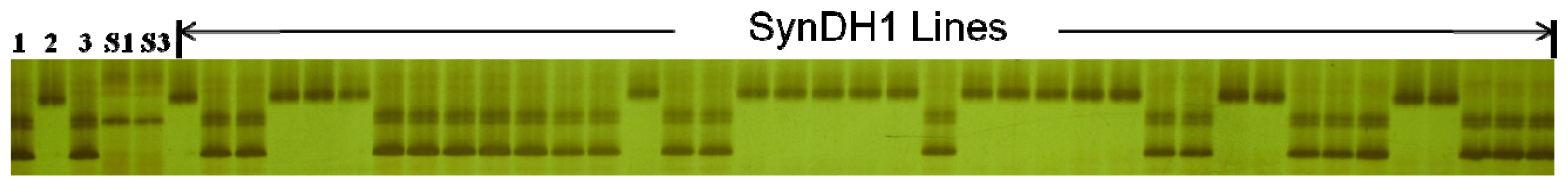
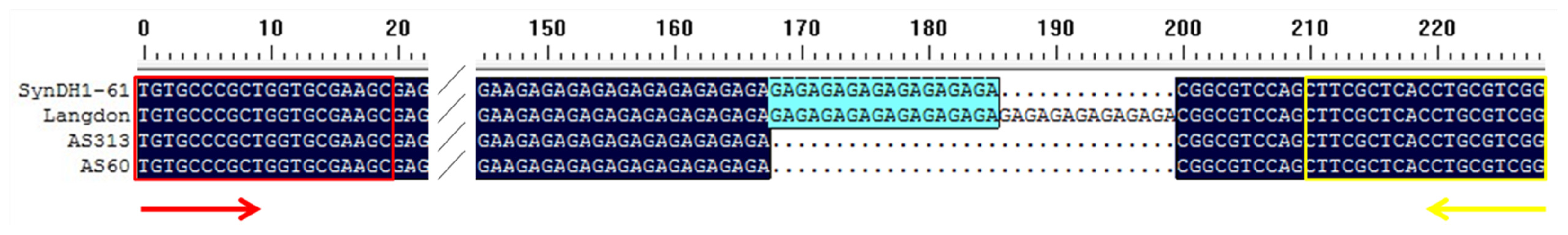
2. Results and Discussion
3. Experimental Section
3.1. Plant Materials
3.2. Simple Sequence Repeats (SSRs) Analysis
3.3. Cloning and Sequencing
3.4. Estimation of SSR Mutation Rate
4. Conclusions
Supplementary Materials
ijms-13-12533-s001.pdfAcknowledgments
- Conflict of InterestThe authors declare no conflict of interest.
References
- Ellegren, H. Microsatellites: Simple sequences with complex evolution. Natl. Rev. Genet 2004, 5, 435–445. [Google Scholar]
- Li, Y.C.; Korol, A.B.; Fahima, T.; Nevo, E. Microsatellites within genes: Structure, function, and evolution. Mol. Biol. Evol 2004, 21, 991–1007. [Google Scholar]
- Buschiazzo, E.; Gemmell, N.J. The rise, fall and renaissance of microsatellites in eukaryotic genomes. Bioessays 2006, 28, 1040–1050. [Google Scholar]
- Raquin, A.L.; Depaulis, F.; Lambert, A.; Galic, N.; Brabant, P.; Goldringer, I. Experimental estimation of mutation rates in a wheat population with a gene genealogy approach. Genetics 2008, 179, 2195–2211. [Google Scholar]
- Schug, M.D.; Mackay, T.F.; Aquadro, C.F. Low mutation rates of microsatellite loci in Drosophila melanogaster. Natl. Genet 1997, 15, 99–102. [Google Scholar]
- Weber, J.L.; Wong, C. Mutation of human short tandem repeats. Hum. Mol. Genet 1993, 2, 1123–1128. [Google Scholar]
- Thuillet, A.C.; Bru, D.; David, J.; Roumet, P.; Santoni, S.; Sourdille, P.; Bataillon, T. Direct estimation of mutation rate for 10 microsatellite loci in durum wheat, Triticum turgidum (L.) Thell. ssp durum desf. Mol. Biol. Evol 2002, 19, 122–125. [Google Scholar]
- Vigouroux, Y.; Jaqueth, J.S.; Matsuoka, Y.; Smith, O.S.; Beavis, W.D.; Smith, J.S.C.; Doebley, J. Rate and pattern of mutation at microsatellite loci in maize. Mol. Biol. Evol 2002, 19, 1251–1260. [Google Scholar]
- McClintock, B. The significance of responses of the genome to challenge. Science 1984, 226, 792–801. [Google Scholar]
- Khasdan, V.; Yaakov, B.; Kraitshtein, Z.; Kashkush, K. Developmental timing of DNA elimination following allopolyploidization in wheat. Genetics 2010, 185, 387–390. [Google Scholar]
- Liu, B.; Vega, J.M.; Feldman, M. Rapid genomic changes in newly synthesized amphiploids of Triticum and Aegilops. II. Changes in low copy coding DNA sequences. Genome 1998, 41, 535–542. [Google Scholar]
- Ozkan, H.; Levy, A.A.; Feldman, M. Allopolyploidy-induced rapid genome evolution in the wheat (Aegilops-Triticum) group. Plant Cell 2001, 13, 1735–1747. [Google Scholar]
- Shaked, H.; Kashkush, K.; Ozkan, H.; Feldman, M.; Levy, A.A. Sequence elimination and cytosine methylation are rapid and reproducible responses of the genome to wide hybridization and allopolyploidy in wheat. Plant Cell 2001, 13, 1749–1759. [Google Scholar]
- Gaeta, R.T.; Pires, J.C.; Iniguez-Luy, F.; Leon, E.; Osborn, T.C. Genomic changes in resynthesized Brassica napus and their effect on gene expression and phenotype. Plant Cell 2007, 19, 3403–3417. [Google Scholar]
- Lukens, L.N.; Pires, J.C.; Leon, E.; Vogelzang, R.; Oslach, L.; Osborn, T. Patterns of sequence loss and cytosine methylation within a population of newly resynthesized Brassica napus allopolyploids. Plant Physiol 2006, 140, 336–348. [Google Scholar]
- Song, K.; Lu, P.; Tang, K.; Osborn, T.C. Rapid genome change in synthetic polyploids of Brassica and its implications for polyploid evolution. Proc. Natl. Acad. Sci. USA 1995, 92, 7719–7723. [Google Scholar]
- Szadkowski, E.; Eber, F.; Huteau, V.; Lodé, M.; Huneau, C.; Belcram, H.; Coriton, O.; Manzanares-Dauleux, M.J.; Delourme, R.; King, G.J.; et al. The first meiosis of resynthesized Brassica napus, a genome blender. New Phytol 2010, 186, 102–112. [Google Scholar]
- Zou, J.; Fu, D.H.; Gong, H.H.; Qian, W.; Xia, W.; Pires, J.C.; Li, R.; Long, Y.; Mason, A.S.; Yang, T.J.; et al. De novo genetic variation associated with retrotransposon activation, genomic rearrangements and trait variation in a recombinant inbred line population of Brassica napus derived from interspecific hybridization with Brassica rapa. Plant J 2011, 68, 212–224. [Google Scholar]
- Bento, M.; Gustafson, J.P.; Viegas, W.; Silva, M. Size matters in Triticeae polyploids: Larger genomes have higher remodeling. Genome 2011, 54, 175–183. [Google Scholar]
- Ma, X.F.; Gustafson, J.P. Allopolyploidization-accommodated genomic sequence changes in Triticale. Ann. Bot 2008, 101, 825–832. [Google Scholar]
- Yuan, Z.; Liu, D.; Zhang, L.; Zhang, L.; Chen, W.; Yan, Z.; Zheng, Y.; Zhang, H.; Yen, Y. Mitotic illegitimate recombination is a mechanism for novel changes in high-molecular-weight glutenin subunits in wheat-rye hybrids. PLoS One 2011, 6, e23511. [Google Scholar]
- Comai, L.; Tyagi, A.P.; Winter, K.; Holmes-Davis, R.; Reynolds, S.H.; Stevens, Y.; Byers, B. Phenotypic instability and rapid gene silencing in newly formed Arabidopsis allotetraploids. Plant Cell 2000, 12, 1551–1568. [Google Scholar]
- Madlung, A.; Tyagi, A.P.; Watson, B.; Jiang, H.; Kagochi, T.; Doerge, R.W.; Martienssen, R.; Comai, L. Genomic changes in synthetic Arabidopsis polyploids. Plant J 2005, 41, 221–230. [Google Scholar]
- Zhang, L.; Zhang, L.; Luo, J.; Chen, W.; Hao, M.; Liu, B.; Yan, Z.; Zhang, B.; Zhang, H.; Zheng, Y.; et al. Synthesizing double haploid hexaploid wheat populations based on a spontaneous alloploidization process. J. Genet. Genomics 2011, 38, 89–94. [Google Scholar]
- Mestiri, I.; Chagué, V.; Tanguy, A.M.; Huneau, C.; Huteau, V.; Belcram, H.; Coriton, O.; Chalhoub, B.; Jahier, J. Newly synthesized wheat allohexaploids display progenitor-dependant meiotic stability and aneuploidy but structural genomic additivity. New Phytol 2010, 186, 86–101. [Google Scholar]
- Liu, B.; Brubaker, C.L.; Mergeai, G.; Cronn, R.C.; Wendel, J.F. Polyploid formation in cotton is not accompanied by rapid genomic changes. Genome 2001, 44, 321–330. [Google Scholar]
- Bottley, A.M.; Xia, G.; Koebner, R.M.D. Homoeologous gene silencing in hexaploid wheat. Plant J 2006, 47, 897–906. [Google Scholar]
- He, P.; Friebe, B.R.; Gill, B.S.; Zhou, J.M. Allopolyploidy alters gene expression in the highly stable hexaploid wheat. Plant Mol. Biol 2003, 52, 401–414. [Google Scholar]
- Wierdl, M.; Dominska, M.; Petes, T.D. Microsatellite instability in yeast: Dependence on the length of the microsatellite. Genetics 1997, 146, 769–779. [Google Scholar]
- Zhang, L.Q.; Liu, D.C.; Zheng, Y.L.; Yan, Z.H.; Dai, S.F.; Li, Y.F.; Jiang, Q.; Ye, Y.Q.; Yen, Y. Frequent occurrence of unreduced gametes in Triticum turgidum-Aegilops tauschii hybrids. Euphytica 2010, 172, 285–294. [Google Scholar]
- Zhang, L.Q.; Liu, D.C.; Yan, Z.H.; Lan, X.J.; Zheng, Y.L.; Zhou, Y. Rapid changes of microsatellite flanking sequence in the allopolyploidization of new synthesized hexaploid wheat. Sci. China C Life Sci 2004, 47, 553–561. [Google Scholar]
- Gupta, P.G.; Balyan, H.B.; Edwards, K.E.; Isaac, P.I.; Korzun, V.K.; Röder, M.; Gautier, M.F.; Joudrier, P.; Schlatter, R.; Dubcovsky, J.; et al. Genetic mapping of 66 new microsatellite (SSR) loci in bread wheat. Theor. Appl. Genet 2002, 105, 413–422. [Google Scholar]
- Guyomarc’h, H.; Sourdille, P.; Charmet, G.; Edwards, J.; Bernard, M. Characterisation of polymorphic microsatellite markers from Aegilops tauschii and transferability to the D-genome of bread wheat. Theor. Appl. Genet 2002, 104, 1164–1172. [Google Scholar]
- Pestsova, E.; Ganal, M.W.; Röder, M.S. Isolation and mapping of microsatellite markers specific for the D genome of bread wheat. Genome 2000, 43, 689–697. [Google Scholar]
- Röder, M.S.; Korzun, V.; Wendehake, K.; Plaschke, J.; Tixier, M.H.; Leroy, P.; Ganal, M.W. A microsatellite map of wheat. Genetics 1998, 149, 2007–2023. [Google Scholar]
- Song, Q.J.; Shi, J.R.; Singh, S.; Fickus, E.W.; Costa, J.M.; Lewis, J.; Gill, B.S.; Ward, R.; Cregan, P.B. Development and mapping of microsatellite (SSR) markers in wheat. Theor. Appl. Genet 2005, 110, 550–560. [Google Scholar]
- Sourdille, P.; Singh, S.; Cadalen, T.; Brown-Guedira, G.L.; Gay, G.; Qi, L.; Gill, B.S.; Dufour, P.; Murigneux, A.; Bernard, M. Microsatellite-based deletion bin system for the establishment of genetic-physical map relationships in wheat (Triticum aestivum L.). Funct. Integr. Genomics 2004, 4, 12–25. [Google Scholar]
- Chen, Q.; Zhang, L.; Yuan, Z.; Yan, Z.; Zheng, Y.; Sun, G.; Liu, D. Empirical verification of heterogeneous DNA fragments generated from wheat genome-specific SSR primers. Can. J. Plant Sci 2008, 88, 1065–1071. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Double haploid populations (hybrid combinations) | Hybrids of haploid (triploid) F1 hybrids | No. S1 lines analyzed * | No. of scored markers ** | No. of estimated S1 alleles | |
|---|---|---|---|---|---|
| Female | Male | ||||
| SynDH1 (LDN/AS313//AS60) | F1 hybrids between T. turgidum LDN and AS313 | Ae. tauschii AS60 | 113 | 81 | 93790 |
| SynDH2 (LDN/AS2255//AS60) | F1 hybrids between T. turgidum LDN and AS2255 | Ae. tauschii AS60 | 89 | 12 | 11570 |
| SynDH3 (PI377655//AS66/AS87) | T. turgidum PI377655 | hybrids between Ae. tauschii AS66 and AS87 | 39 | 79 | 42900 |
| Total | - | - | 241 | 160 | 148260 |
| Materials | No. of sequenced clones | Number of clones with different sequence length | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 193 (bp) | 195 (bp) | 197 (bp) | 199 (bp) | 213 (bp) | 215 (bp) | 217 (bp) | 219 (bp) | 223 (bp) | 227 (bp) | 229 (bp) | 231 (bp) | 233 (bp) | ||
| AS313 | 8 | - | 1 | 7 | - | - | - | - | - | - | - | - | - | - |
| Langdon | 9 | - | - | - | - | - | - | - | - | 1 | 2 | 4 | 1 | 1 |
| AS60 | 9 | 1 | 1 | 6 | 1 | - | - | - | - | - | - | - | - | - |
| SynDH1-61 (S1) | 13 | - | - | - | - | - | 9 | 3 | 1 | - | - | - | - | - |
| SynDH1-61 (S2) | 11 | - | - | 1 | - | 3 | 4 | 3 | - | - | - | - | - | - |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Luo, J.; Hao, M.; Zhang, L.; Chen, J.; Zhang, L.; Yuan, Z.; Yan, Z.; Zheng, Y.; Zhang, H.; Yen, Y.; et al. Microsatellite Mutation Rate during Allohexaploidization of Newly Resynthesized Wheat. Int. J. Mol. Sci. 2012, 13, 12533-12543. https://doi.org/10.3390/ijms131012533
Luo J, Hao M, Zhang L, Chen J, Zhang L, Yuan Z, Yan Z, Zheng Y, Zhang H, Yen Y, et al. Microsatellite Mutation Rate during Allohexaploidization of Newly Resynthesized Wheat. International Journal of Molecular Sciences. 2012; 13(10):12533-12543. https://doi.org/10.3390/ijms131012533
Chicago/Turabian StyleLuo, Jiangtao, Ming Hao, Li Zhang, Jixiang Chen, Lianquan Zhang, Zhongwei Yuan, Zehong Yan, Youliang Zheng, Huaigang Zhang, Yang Yen, and et al. 2012. "Microsatellite Mutation Rate during Allohexaploidization of Newly Resynthesized Wheat" International Journal of Molecular Sciences 13, no. 10: 12533-12543. https://doi.org/10.3390/ijms131012533
APA StyleLuo, J., Hao, M., Zhang, L., Chen, J., Zhang, L., Yuan, Z., Yan, Z., Zheng, Y., Zhang, H., Yen, Y., & Liu, D. (2012). Microsatellite Mutation Rate during Allohexaploidization of Newly Resynthesized Wheat. International Journal of Molecular Sciences, 13(10), 12533-12543. https://doi.org/10.3390/ijms131012533