Callose Deposition Is Required for Somatic Embryogenesis in Plasmolyzed Eleutherococcus senticosus Zygotic Embryos

Abstract
:
1. Introduction
2. Results and Discussion
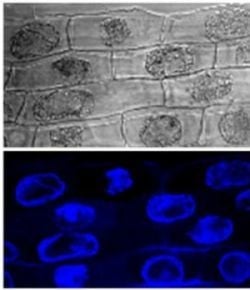
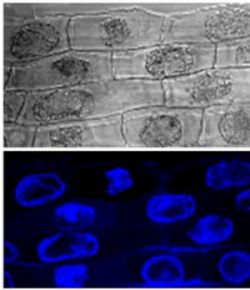
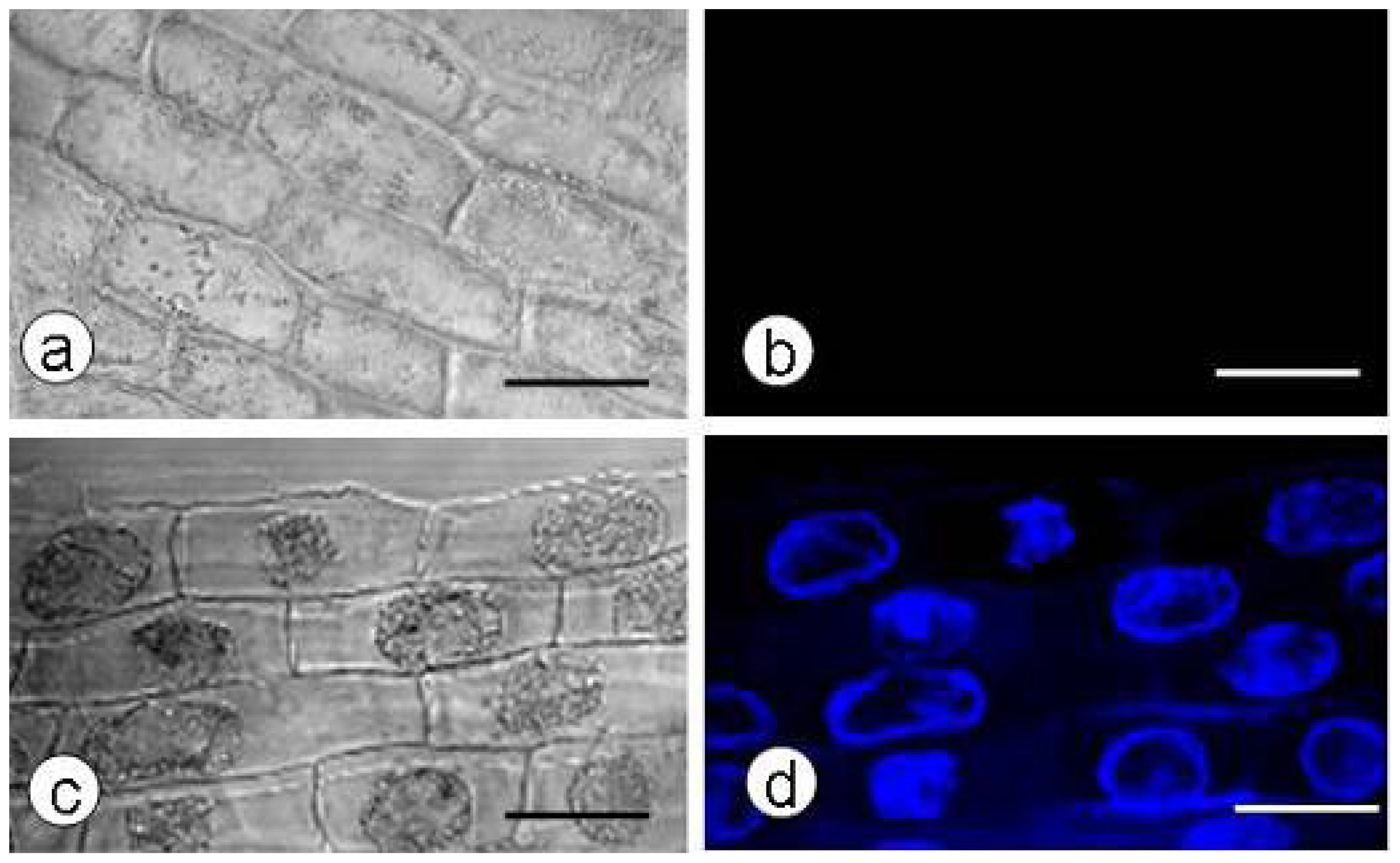
2.1. Callose Content in Zygotic Embryos Plasmolyzed with Mannitol
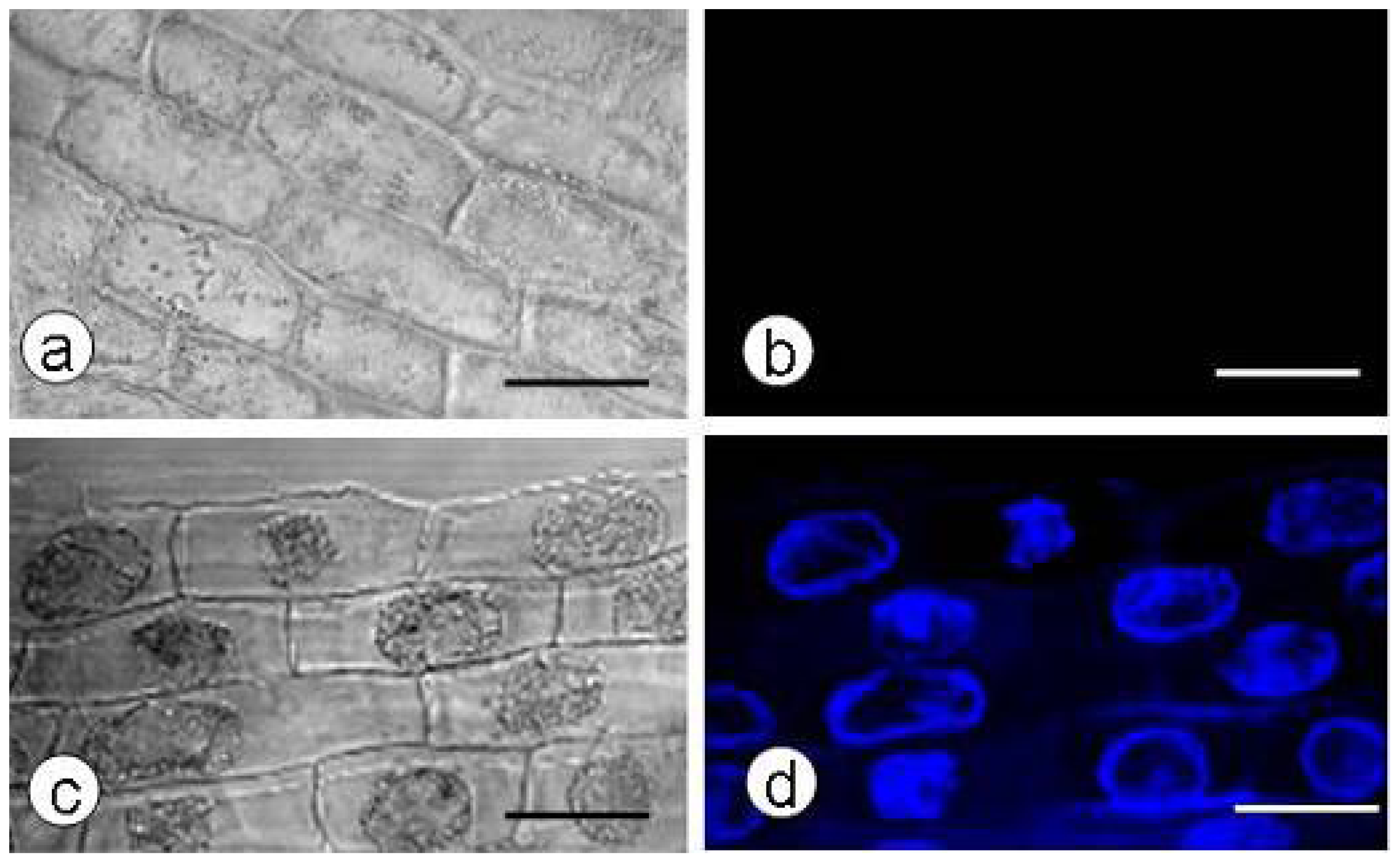
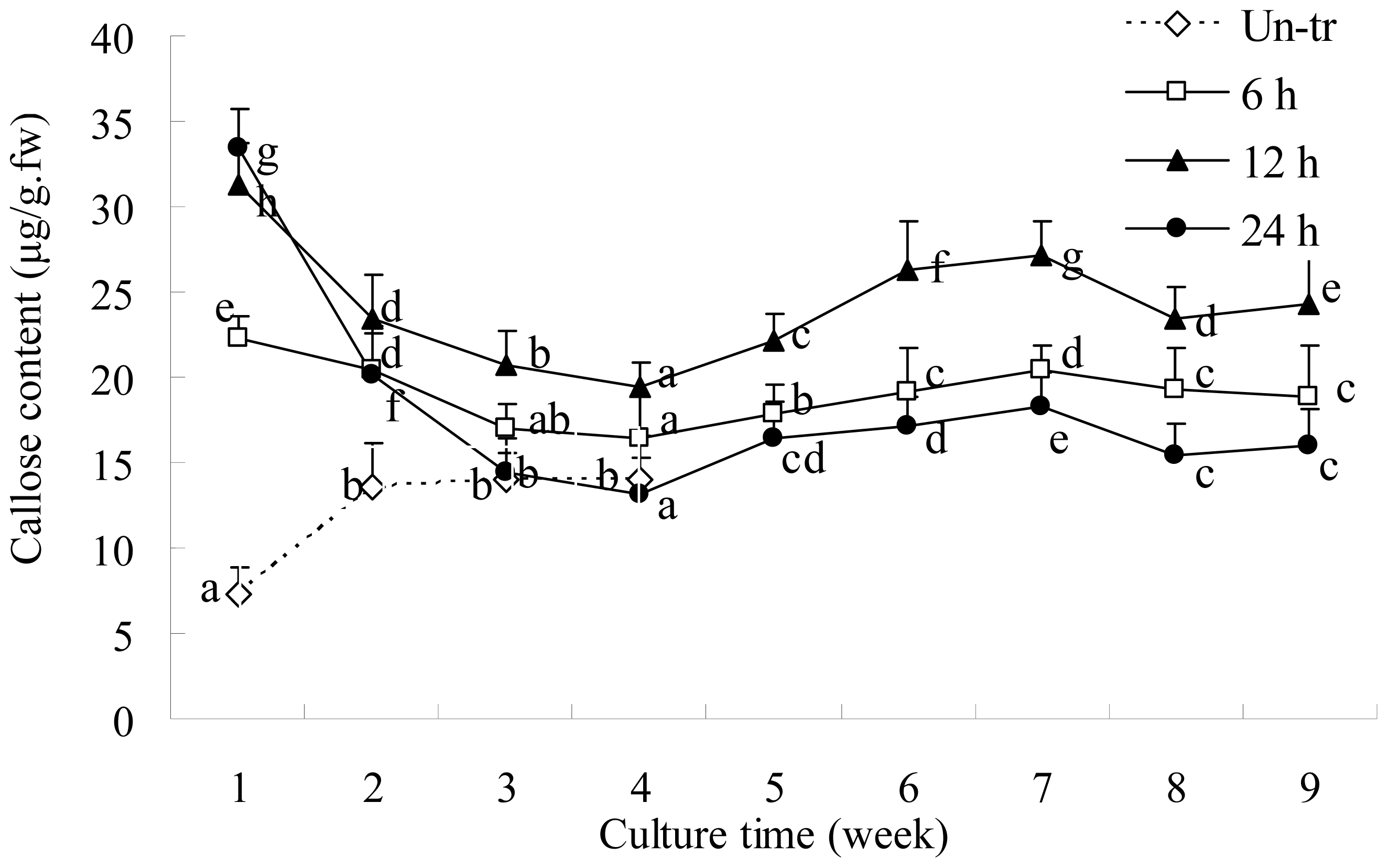
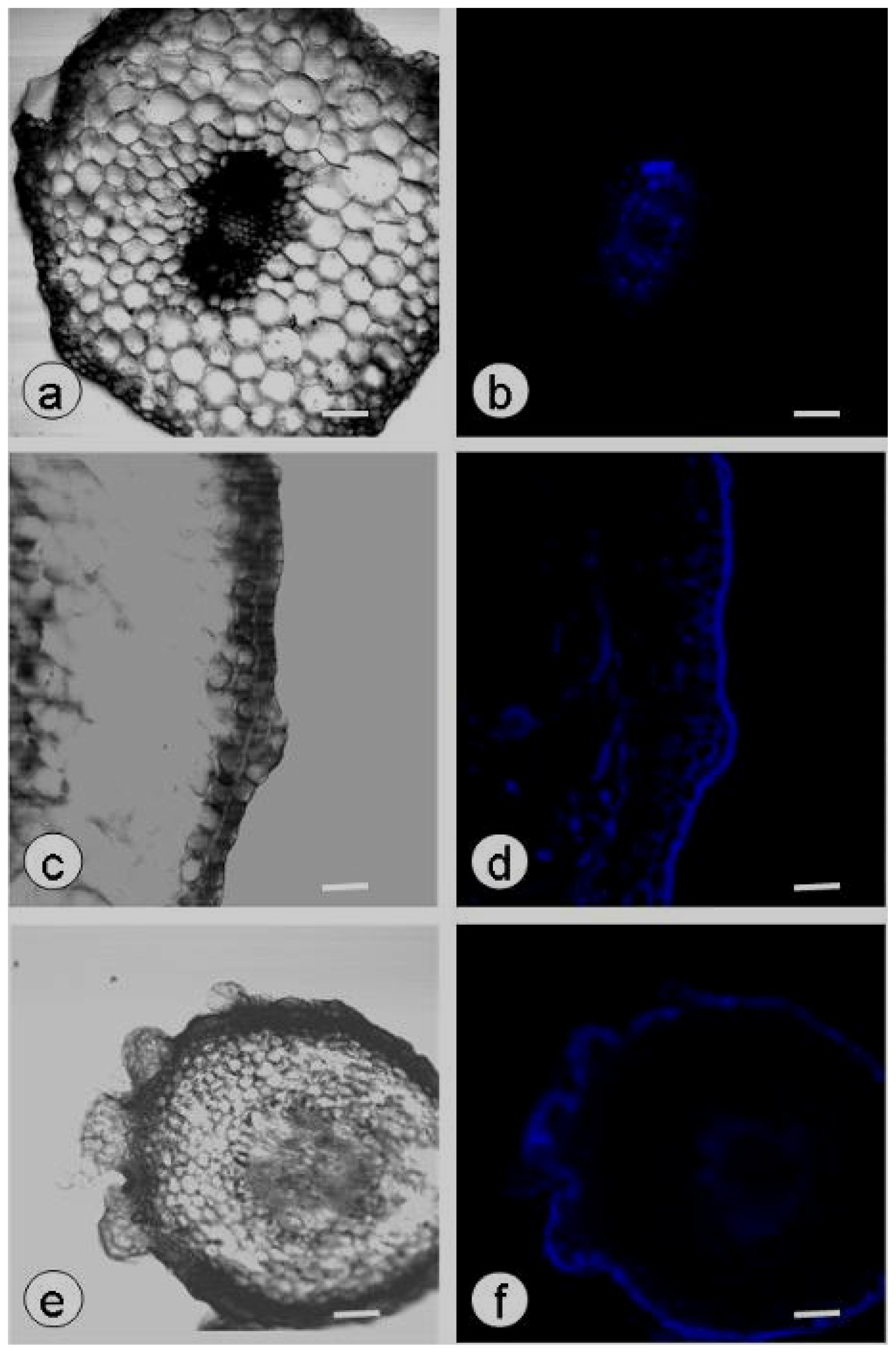
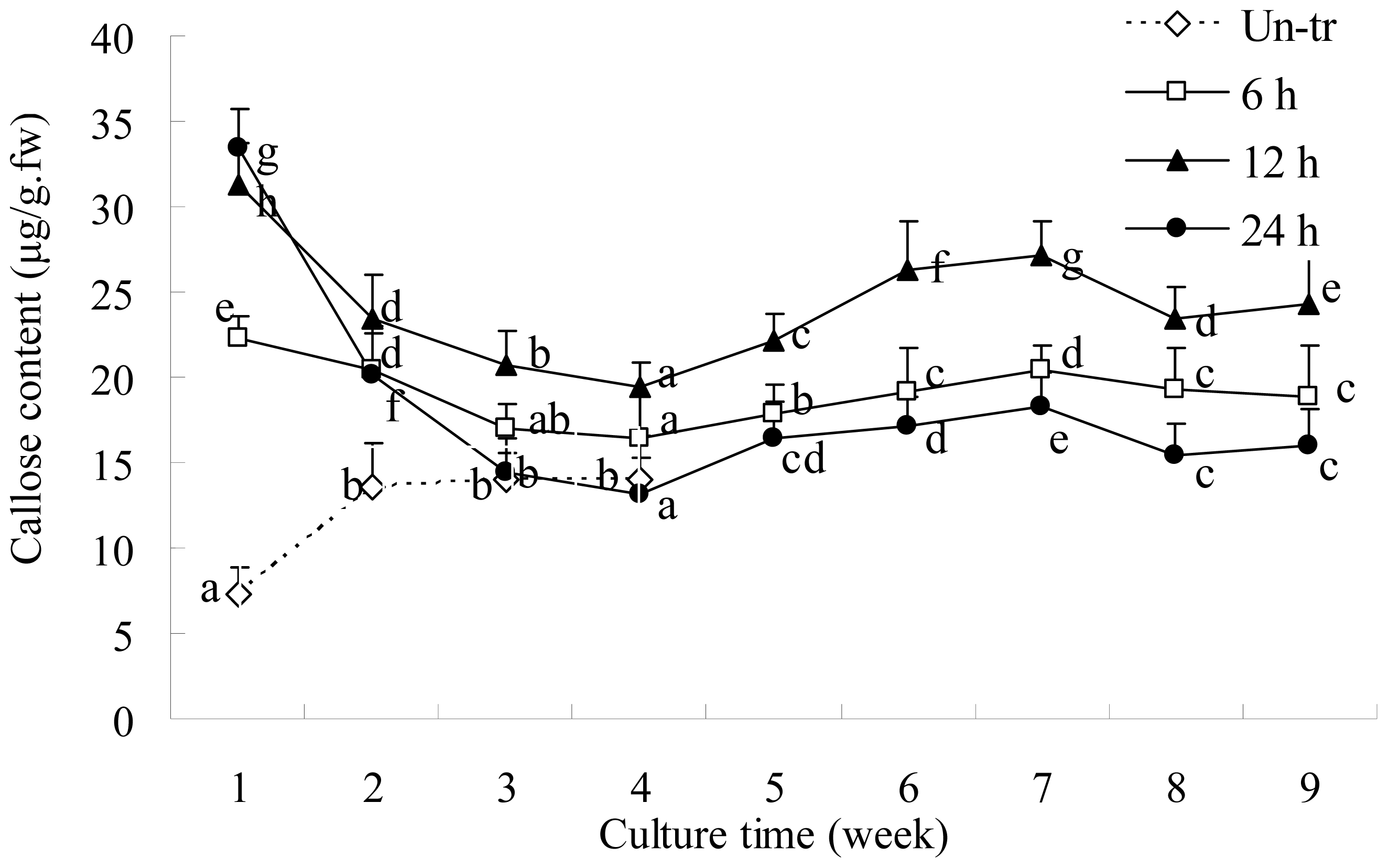
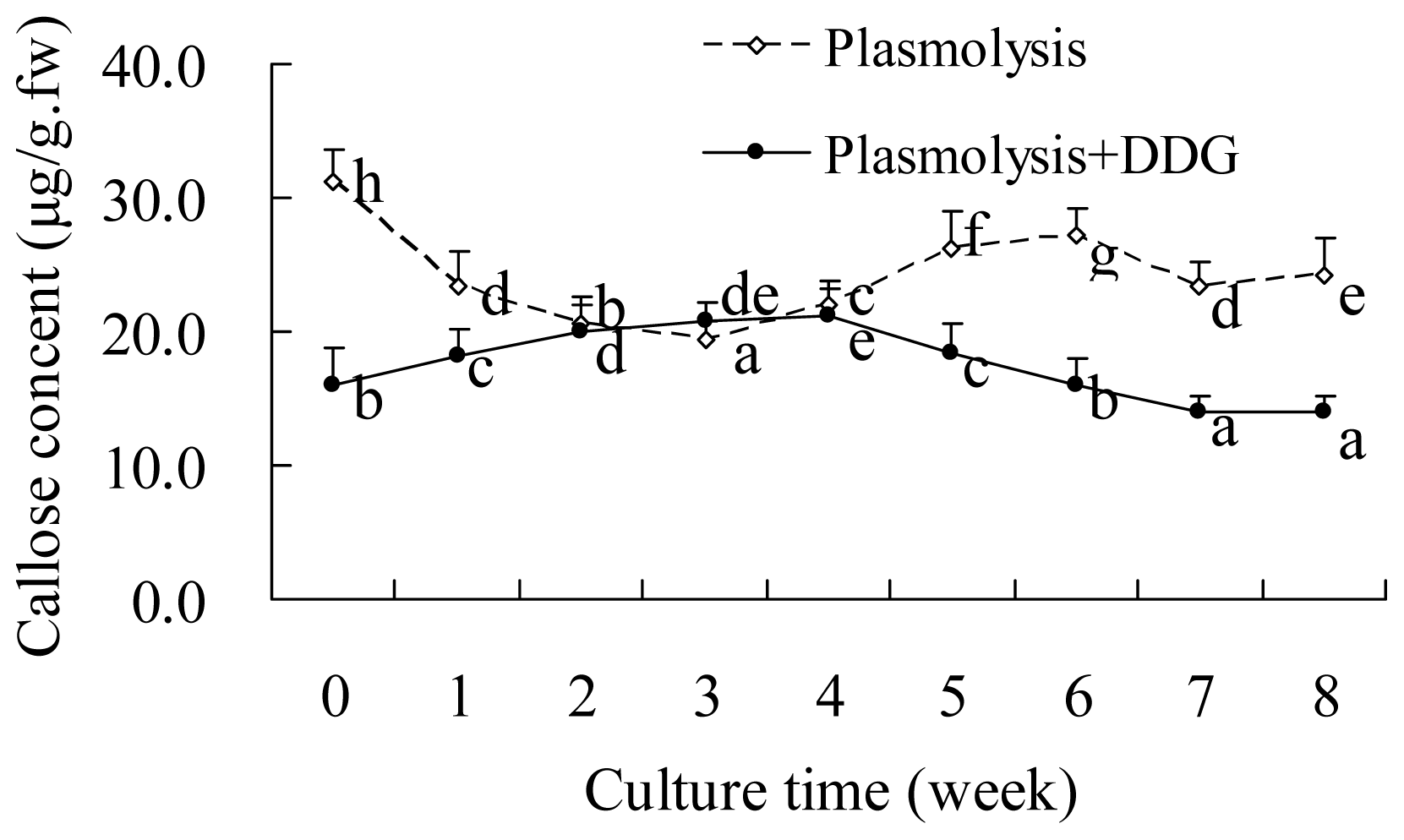
2.2. Callose Content in Plasmolyzed Zygotic Embryos during SE
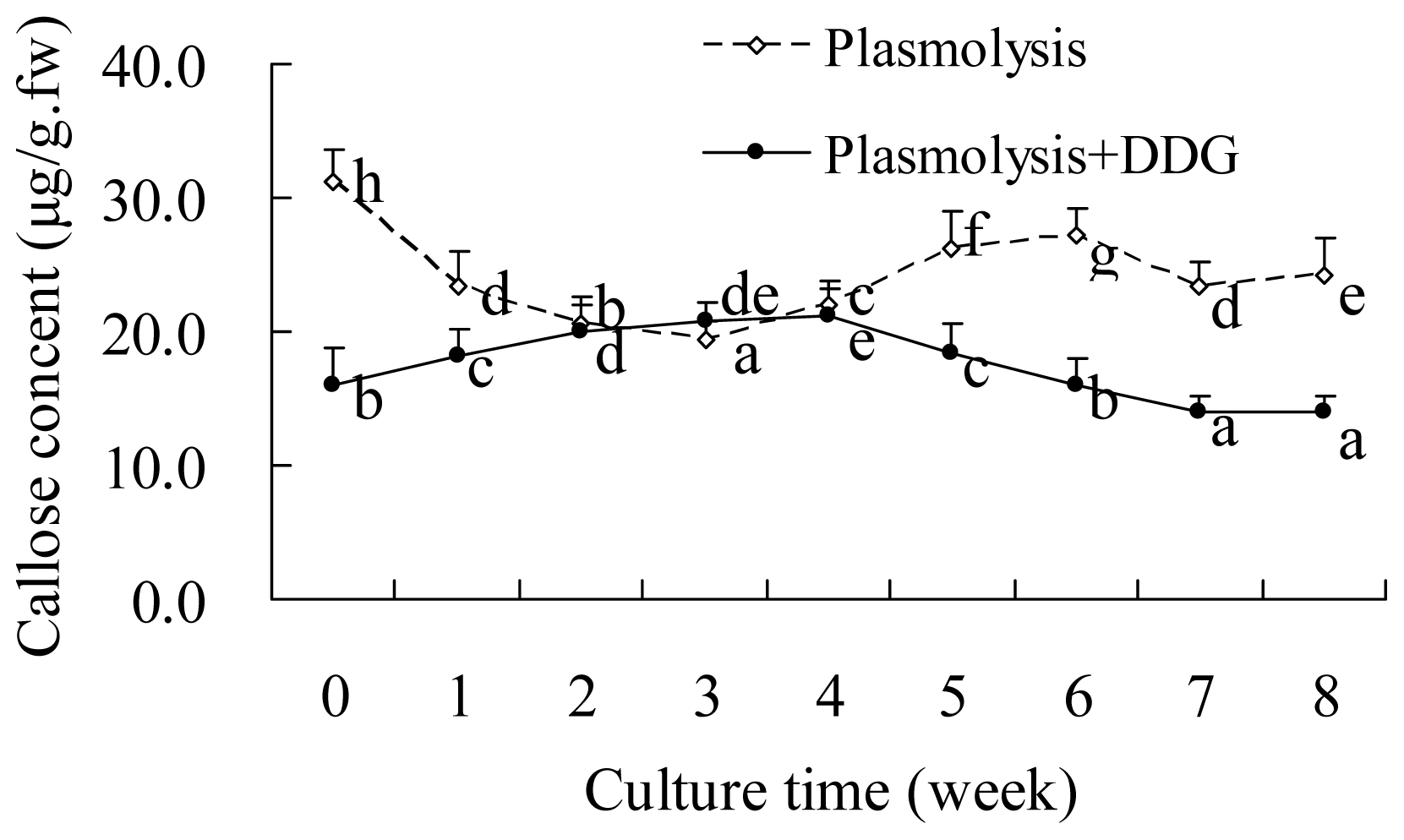
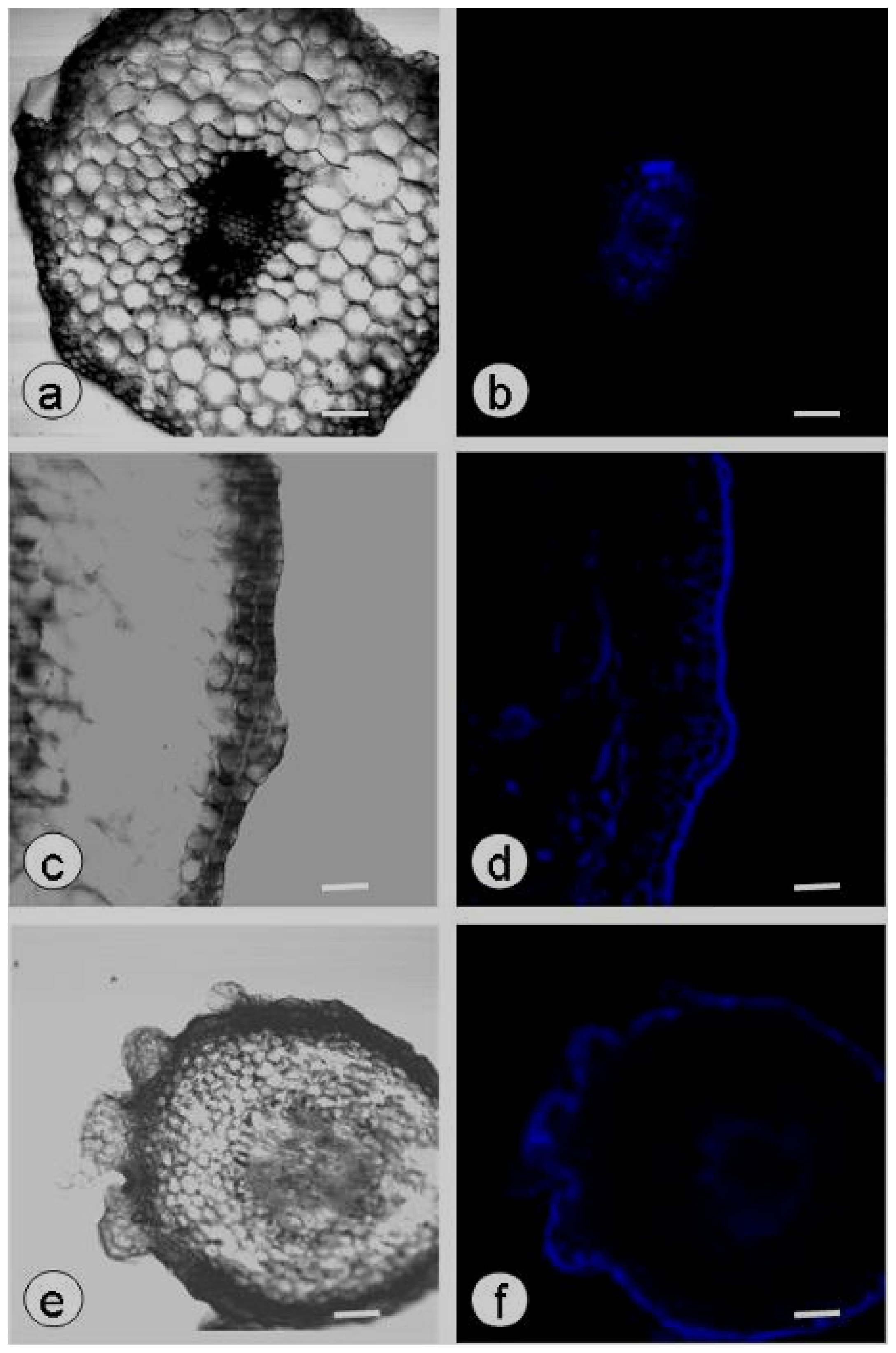
2.3. DDG-Inhibition of Callose Deposition in Zygotic Embryos
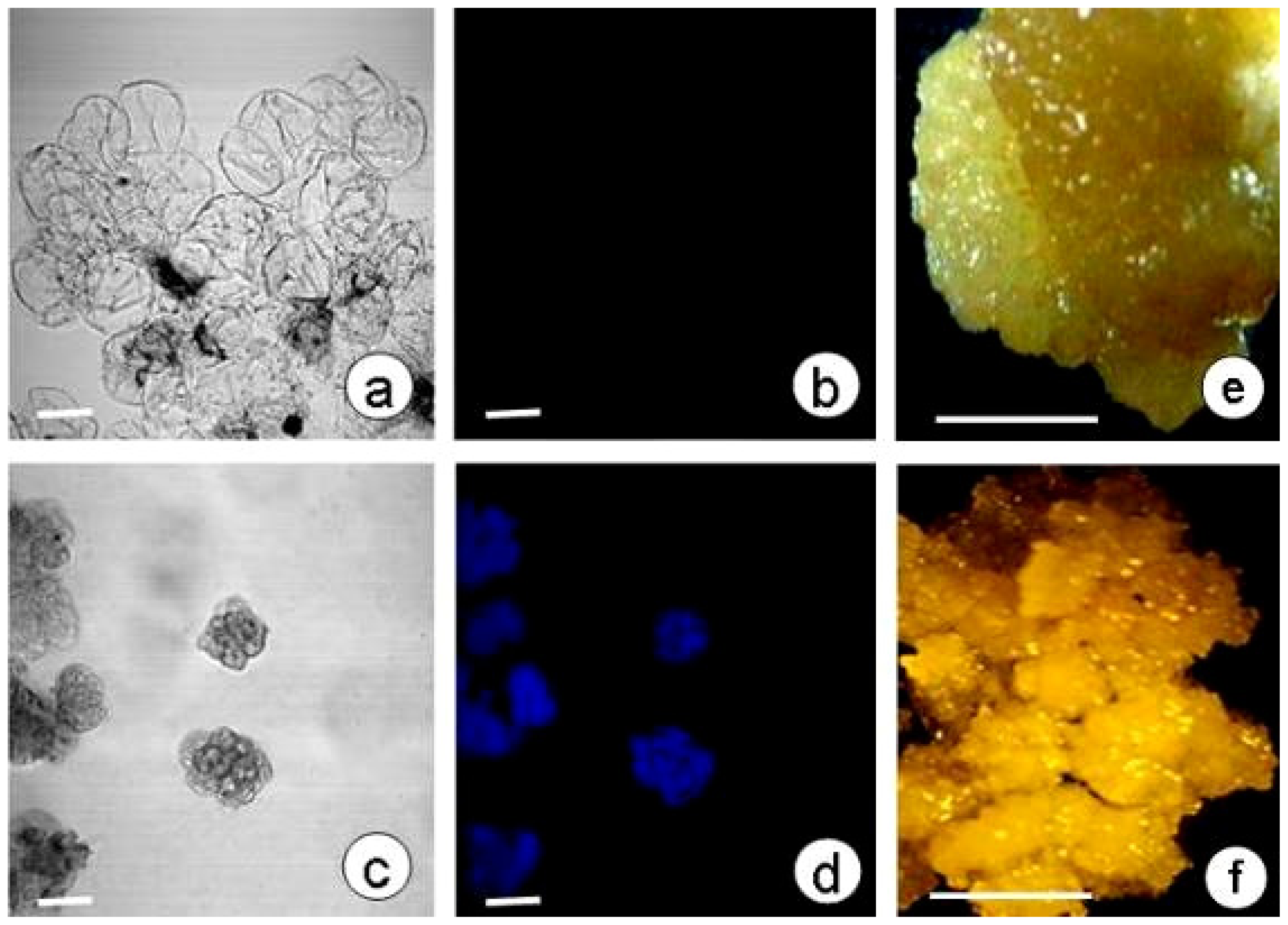
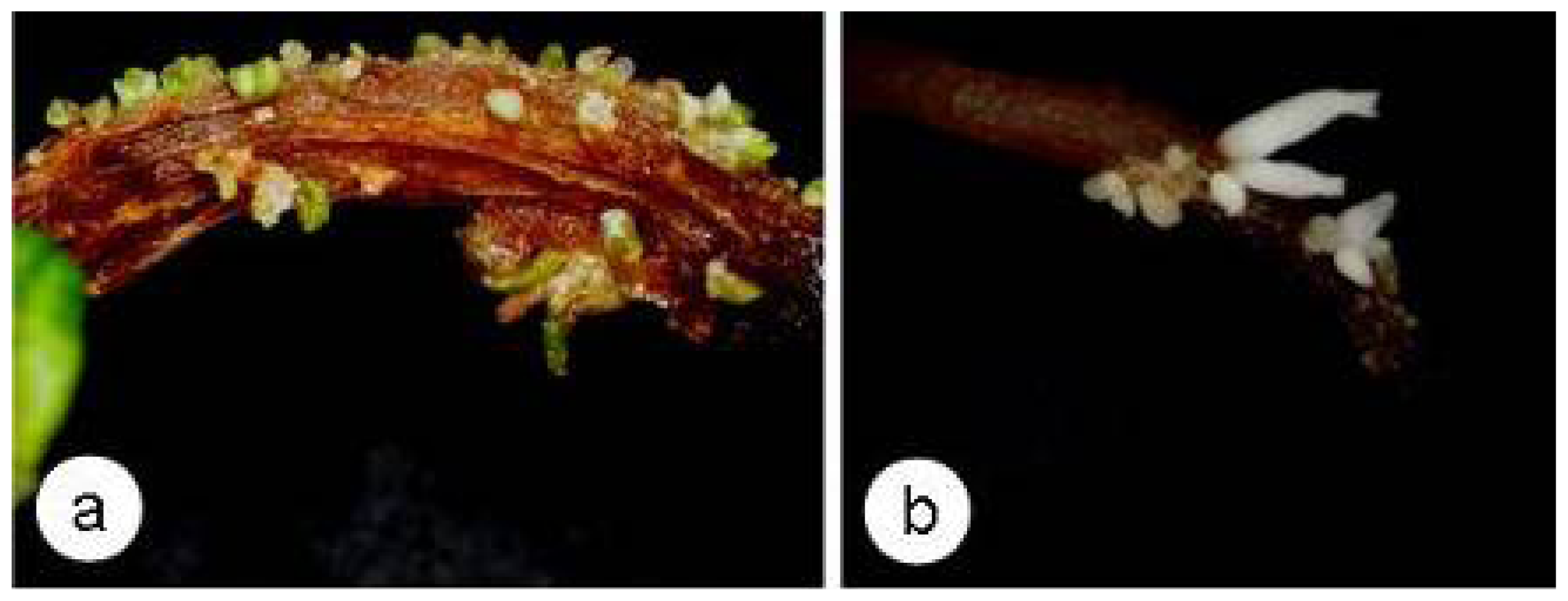
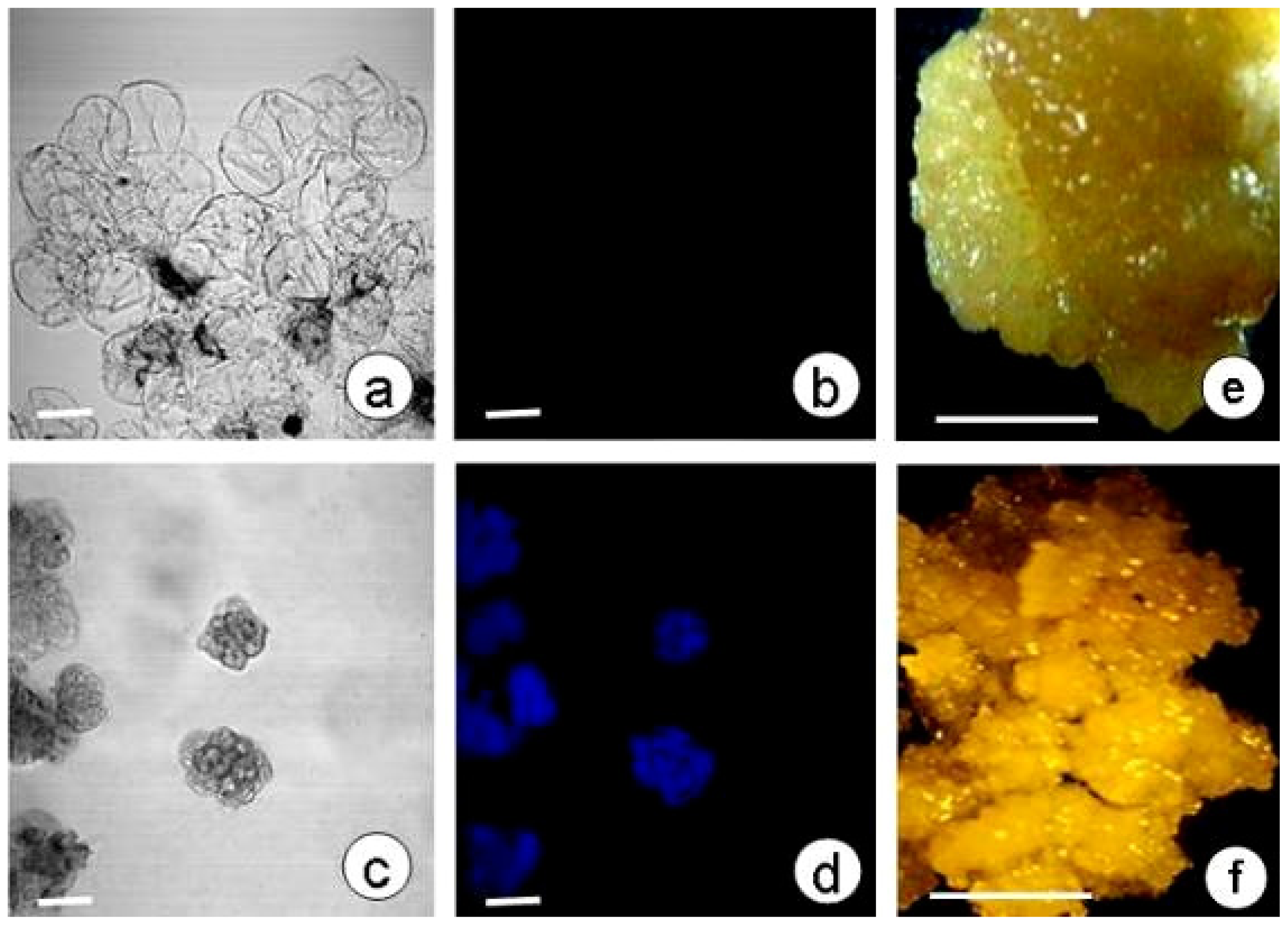
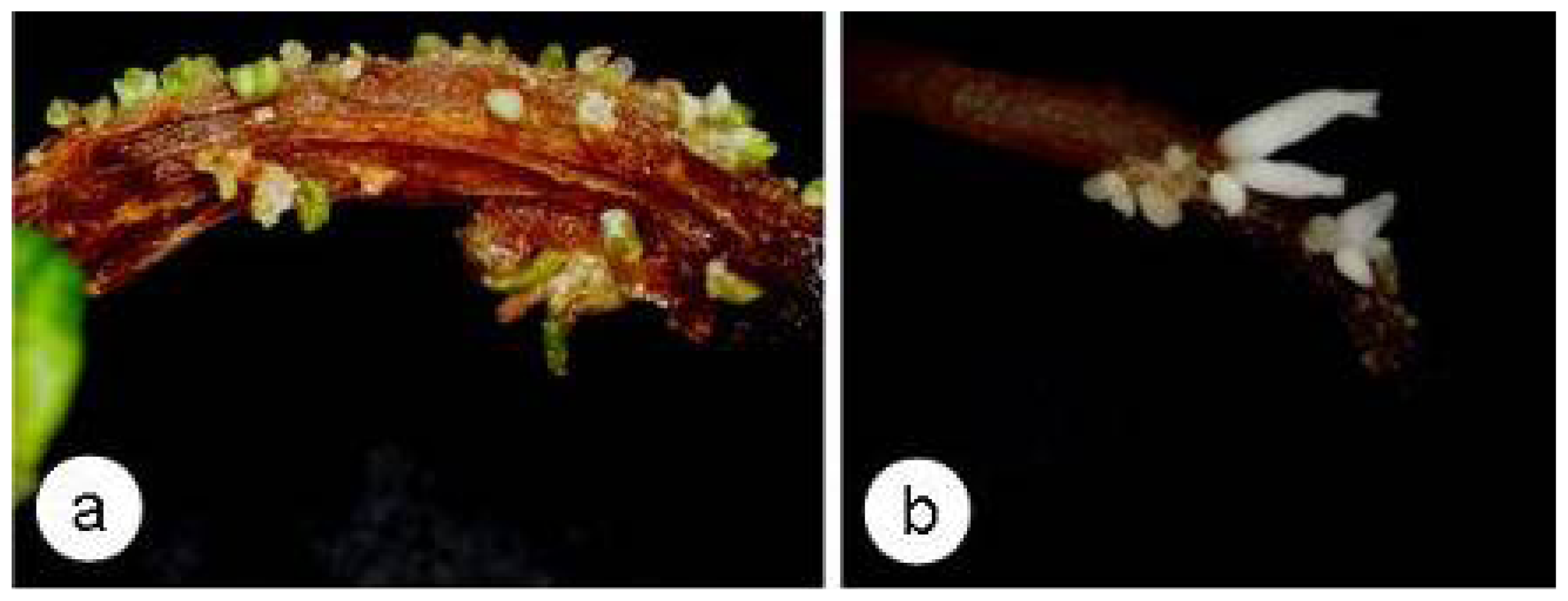
2.4. SE and Callose Deposition after DDG Treatment
3. Experimental Section
3.1. Callose Deposition in Zygotic Embryos after Plasmolysis and during SE
3.2. Induction of SE in Plasmolyzed Explants and Inhibition of Callose Deposition
3.3. Culture of Embryogenic and Non-Embryogenic Calli
3.4. Spectrofluorometric Assay of Callose Content
3.5. Localization of Callose by Confocal Laser Scanning Microscopy
3.6. Condition of Tissue Culture and Statistical Analysis
4. Conclusions
Acknowledgments
Abbreviations
| SE | somatic embryogenesis |
| 2,4-D | 2,4-dichlorophenoxy acetic acid |
| BA | 6-benzylaminopurine |
| CLSM | confocal laser scanning microscopy |
| DDG | 2-deoxy-d-glucose |
| NAA | naphthylacetic acid |
| PGR | plant growth regulator |
References
- Zimmerman, J.L. Somatic embryogenesis: a model for early development in higher plants. Plant Cell 1993, 5, 1411–1423. [Google Scholar]
- Fehér, A.; Pasternak, T.P.; Dudits, D. Transition of somatic plant cells to an embryogenic state. Plant Cell Tissue Org. Cult 2003, 74, 201–228. [Google Scholar]
- Nolan, K.E.; Saeed, N.A.; Rose, R.J. The stress kinase gene MtSK1 in Medicago. truncatula with particular reference to somatic embryogenesis. Plant Cell Rep 2006, 25, 711–722. [Google Scholar]
- Kiyosue, T.; Takano, K.; kamada, H.; Harad, H. Induction of somatic embryogenesis in carrot by heavy metal ions. Can. J. Bot 1990, 68, 2301–2303. [Google Scholar]
- Kiyosue, T.; Satoh, S.; Kamada, H.; Harada, H. Somatic embryogenesis in higher plants. J. Plant Res 1993, 3, 75–82. [Google Scholar]
- Kamada, H.; Tachikawa, Y.; Saitou, T.; Harada, H. Heat stresses induction of carrot somatic embryogenesis. Plant Tissue Cult. Lett 1994, 11, 229–232. [Google Scholar]
- Iwai, M.I.; Umehara, M.; Satoh, S.; Kamada, H. Stress-induced somatic embryogenesis in vegetative tissues of Arabidopsis thaliana. Plant J 2003, 34, 107–114. [Google Scholar]
- Namasivayam, P. Acquisition of embryogenic competence during somatic embryogenesis. Plant Cell Tissue Org. Cult 2007, 90, 1–8. [Google Scholar]
- Karami, O.; Saidi, A. The molecular basis for stress-induced acquisition of somatic embryogesis. Mol. Biol. Rep 2010, 37, 2493–2507. [Google Scholar]
- Ma, J.; He, Y.H.; Hu, Z.Y.; Xe, W.T.; Xia, J.X.; Guo, C.H.; Lin, S.Q.; Cao, L.; Chen, C.J.; Wu, C.H.; et al. Characterization and expression analysis of AcSERK2, a somatic embryogenesis and stress resistance related gene in pineapple. Gene 2012. 101016/j.gene.2012.03.013. [Google Scholar]
- Jacobs, A.K.; Lipka, V.; Burton, R.A.; Panstruga, R.; Strizhov, N.; Schulze-lefert, P.; Fincher, G.B. An Arabidopsis callose synthase, GSL5, is required for wound and papillary callose formation. Plant Cell 2003, 15, 2503–2513. [Google Scholar]
- Dubois, T.; Guedira, M.; Dubois, J.; Vasseur, J. Direct somatic embryogenesis in leaves of Cichorium. A histological and S.E.M study of early stages. Protoplasma 1991, 162, 120–127. [Google Scholar]
- Verdeil, J.L.; Hocher, V.; Huet, C.; Grosdemange, F.; Escoute, J.; Ferriere, N.; Nicole, M. Ultrastructural changes in cocunut calli-associated with the acquisition of embryogenic competence. Ann. Bot 2001, 88, 9–21. [Google Scholar]
- Grimault, V.; Helleboid, S.; Vasseur, J.; Hilbert, J.L. Co-Localization of β-1,3-Glucanases and callose during somatic embryogenesis in cichorium. Plant Signal. Behav 2007, 2, 455–461. [Google Scholar]
- You, X.L.; Yi, J.S.; Choi, Y.E. Cellular change and callose accumulation in zygotic embryos of Eleutherococcus senticosus caused by plasmolyzing pretreatment result in high frequency of single-cell-derived somatic embryogenesis. Protoplasma 2006, 227, 105–112. [Google Scholar]
- You, X.L. In Vitro Mass-Propagation of Eleutherococcus senticosus Maxim. Ph.D. Thesis, Kangwon National University, Chuncheon, Korea, August 2005. [Google Scholar]
- Hirano, Y.; Pannatier, E.G.; Zimmermann, S.; Brunner, I. Induction of callose in roots of Norway spruce seedlings after short-term exposure to aluminum. Tree Physiol 2004, 24, 1279–1283. [Google Scholar]
- Dong, X.; Hong, Z.; Sivaramakrishnan, M.; Mahfouz, M.; Verma, D.P. Callose synthase (CalS5) is required for exine formation during microgametogenesis and for pollen viability in Arabidopsis. Plant J 2005, 42, 315–328. [Google Scholar]
- Abercrombie, J.M.; O’Meara, B.C.; Moffett, A.R.; Williams, J.H. Developmental evolution of flowering plant pollen tube cell walls: Callose synthase (CalS) gene expression patterns. EvoDevo 2011, 2, 1–14. [Google Scholar]
- Hofmann, J.; Youssef-Banora, M.; de Almeida-Engler, J.; Grundler, F.M. The role of callose deposition along plasmodesmata in nematode feeding sites. Mol. Plant Microb. Interact 2010, 23, 549–557. [Google Scholar]
- Hu, X.L.; An, Y.; Xia, D.A.; You, X.L. Plasmolysis treatment enhances the expression of callose synthase gene in zygotic embryos of Eleutherococcus senticosus. J. Forest Res 2010, 21, 189–192. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmolysis time (h) | Callose concentration (μg/g.fw) |
|---|---|
| Control | 6.2 d ± 1.42 |
| 3.0 | 17.3 c ± 2.21 |
| 6.0 | 22.2 b ± 3.33 |
| 12.0 | 30.5 a ± 3.54 |
| 24.0 | 31.3 a ± 2.55 |
| DDG concentration (μM) | Callose concentration (μg/g.fw) |
|---|---|
| Untreated | 7.1 a ± 1.32 |
| control | 31.3 e ± 2.13 |
| 50 | 19.5 c ± 1.51 |
| 100 | 20.3 cd ± 1.22 |
| 200 | 20.6 d ± 1.85 |
| 500 | 16.1 b ± 1.05 |
| 1000 | 15.5 b ± 1.62 |
| 2000 | 15.8 b ± 1.44 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tao, L.; Yang, Y.; Wang, Q.; You, X. Callose Deposition Is Required for Somatic Embryogenesis in Plasmolyzed Eleutherococcus senticosus Zygotic Embryos. Int. J. Mol. Sci. 2012, 13, 14115-14126. https://doi.org/10.3390/ijms131114115
Tao L, Yang Y, Wang Q, You X. Callose Deposition Is Required for Somatic Embryogenesis in Plasmolyzed Eleutherococcus senticosus Zygotic Embryos. International Journal of Molecular Sciences. 2012; 13(11):14115-14126. https://doi.org/10.3390/ijms131114115
Chicago/Turabian StyleTao, Lei, Yang Yang, Qiuyu Wang, and Xiangling You. 2012. "Callose Deposition Is Required for Somatic Embryogenesis in Plasmolyzed Eleutherococcus senticosus Zygotic Embryos" International Journal of Molecular Sciences 13, no. 11: 14115-14126. https://doi.org/10.3390/ijms131114115