The -308G/A of Tumor Necrosis Factor (TNF)-α and 825C/T of Guanidine Nucleotide Binding Protein 3 (GNB3) are Associated with the Onset of Acute Myocardial Infarction and Obesity in Taiwan

Abstract
:1. Introduction
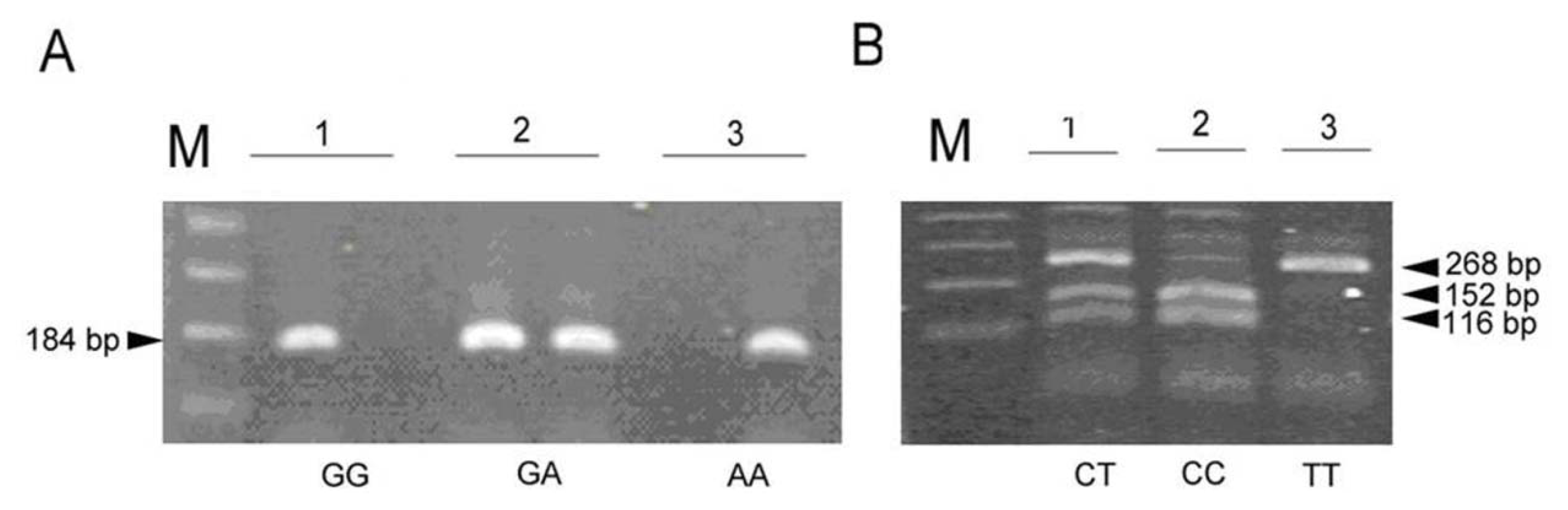
2. Results
3. Materials and methods
3.1. Patients
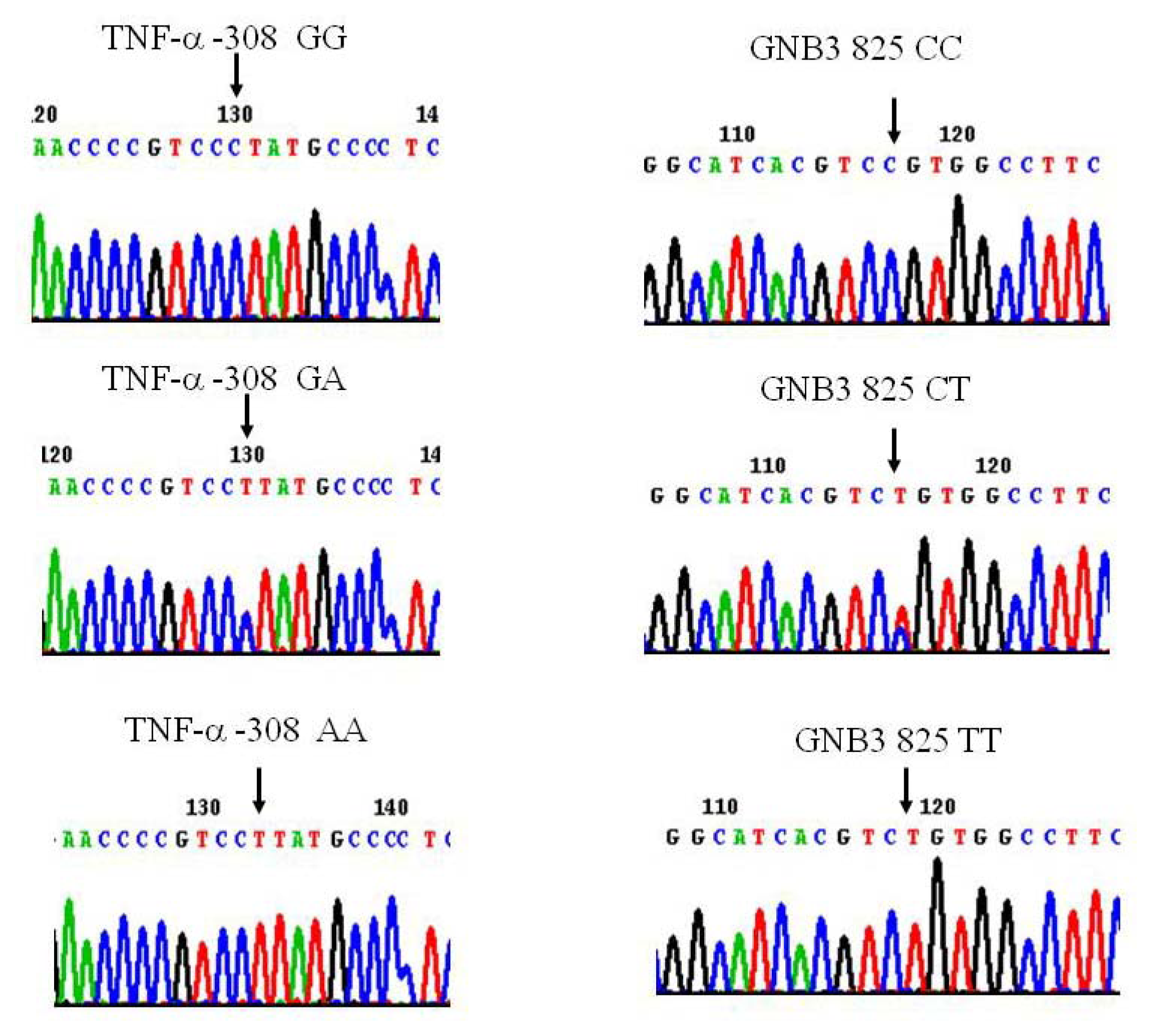
3.2. Genotyping
3.3. Statistical Analysis
4. Discussion
5. Conclusions
Acknowledgements
References
- Wang, Y.C.; Hwang, J.J.; Hung, C.S.; Kao, H.L.; Chiang, F.T.; Tseng, C.D. Outcome of primary percutaneous coronary intervention in octogenarians with acute myocardial infarction. J. Formos. Med. Assoc 2006, 105, 451–458. [Google Scholar]
- Liu, P.Y.; Chen, J.H.; Li, Y.H.; Wu, H.L.; Shi, G.Y. Synergistic effect of stromelysin-1 (matrix metallo-proteinase-3) promoter 5A/6A polymorphism with smoking on the onset of young acute myocardial infarction. Thromb. Haemost 2003, 90, 132–139. [Google Scholar]
- Vendrell, J.; Fernandez-Real, J.M.; Gutierrez, C.; Zamora, A.; Simon, I.; Bardaji, A.; Ricart, W.; Richart, C. A polymorphism in the promoter of the tumor necrosis factor-alpha gene (-308) is associated with coronary heart disease in type 2 diabetic patients. Atherosclerosis 2003, 167, 257–264. [Google Scholar]
- Bernard, V.; Pillois, X.; Dubus, I.; Benchimol, D.; Labouyrie, J.P.; Couffinhal, T.; Coste, P.; Bonnet, J. The -308 G/A tumor necrosis factor-alpha gene dimorphism: A risk factor for unstable angina. Clin. Chem. Lab. Med 2003, 41, 511–516. [Google Scholar]
- Dedoussis, G.V.; Panagiotakos, D.B.; Vidra, N.V.; Louizou, E.; Chrysohoou, C.; Germanos, A.; Mantas, Y.; Tokmakidis, S.; Pitsavos, C.; Stefanadis, C. Association between TNF-alpha -308G>A polymorphism and the development of acute coronary syndromes in Greek subjects: The CARDIO2000-GENE Study. Genet. Med 2005, 7, 411–416. [Google Scholar]
- Antonicelli, R.; Olivieri, F.; Cavallone, L.; Spazzafumo, L.; Bonafe, M.; Marchegiani, F.; Cardelli, M.; Galeazzi, R.; Giovagnetti, S.; Perna, G.P.; et al. Tumor necrosis factor-alpha gene -308G>A polymorphism is associated with ST-elevation myocardial infarction and with high plasma levels of biochemical ischemia markers. Coron. Artery Dis 2005, 16, 489–493. [Google Scholar]
- Kang, M.; Chung, K.Y.; Walker, J.W. G-protein coupled receptor signaling in myocardium: not for the faint of heart. Physiology (Bethesda) 2007, 22, 174–184. [Google Scholar]
- Schunkert, H.; Hense, H.W.; Doring, A.; Riegger, G.A.; Siffert, W. Association between a polymorphism in the G protein beta3 subunit gene and lower renin and elevated diastolic blood pressure levels. Hypertension 1998, 32, 510–513. [Google Scholar]
- Siffert, W.; Rosskopf, D.; Siffert, G.; Busch, S.; Moritz, A.; Erbel, R.; Sharma, A.M.; Ritz, E.; Wichmann, H.E.; Jakobs, K.H.; et al. Association of a human G-protein beta3 subunit variant with hypertension. Nat. Genet 1998, 18, 45–48. [Google Scholar]
- Siffert, W. G protein polymorphisms in hypertension, atherosclerosis, and diabetes. Annu. Rev. Med 2005, 56, 17–28. [Google Scholar]
- Wascher, T.C.; Paulweber, B.; Malaimare, L.; Stadlmayr, A.; Iglseder, B.; Schmoelzer, I.; Renner, W. Associations of a human G protein beta3 subunit dimorphism with insulin resistance and carotid atherosclerosis. Stroke 2003, 34, 605–609. [Google Scholar]
- Zhang, L.; Zhang, H.; Sun, K.; Song, Y.; Hui, R.; Huang, X. The 825C/T polymorphism of G-protein beta3 subunit gene and risk of ischaemic stroke. J. Hum. Hypertens 2005, 19, 709–714. [Google Scholar]
- Klintschar, M.; Stiller, D.; Schwaiger, P.; Kleiber, M. DNA polymorphisms in the tyrosine hydroxylase and GNB3 genes: Association with unexpected death from acute myocardial infarction and increased heart weight. Forensic. Sci. Int 2005, 153, 142–146. [Google Scholar]
- Beige, J.; Hohenbleicher, H.; Distler, A.; Sharma, A.M. G-Protein beta3 subunit C825T variant and ambulatory blood pressure in essential hypertension. Hypertension 1999, 33, 1049–1051. [Google Scholar]
- Dong, Y.; Zhu, H.; Sagnella, G.A.; Carter, N.D.; Cook, D.G.; Cappuccio, F.P. Association between the C825T polymorphism of the G protein beta3-subunit gene and hypertension in blacks. Hypertension 1999, 34, 1193–1196. [Google Scholar]
- Huang, X.; Ju, Z.; Song, Y.; Zhang, H.; Sun, K.; Lu, H.; Yang, Z.; Jose, P.A.; Zhou, G.; Wang, M.; et al. Lack of association between the G protein beta3 subunit gene and essential hypertension in Chinese: a case-control and a family-based study. J. Mol. Med. (Berl.) 2003, 81, 729–735. [Google Scholar]
- Li, B.; Ge, D.; Wang, Y.; Zhao, W.; Zhou, X.; Gu, D.; Chen, R. G protein beta 3 subunit gene variants and essential hypertension in the northern Chinese Han population. Ann. Hum. Genet 2005, 69, 468–473. [Google Scholar]
- Siffert, W.; Forster, P.; Jockel, K.H.; Mvere, D.A.; Brinkmann, B.; Naber, C.; Crookes, R.; Du, P.H.A.; Epplen, J.T.; Fridey, J.; et al. Worldwide ethnic distribution of the G protein beta3 subunit 825T allele and its association with obesity in Caucasian, Chinese, and Black African individuals. J. Am. Soc. Nephrol 1999, 10, 1921–1930. [Google Scholar]
- Wang, Y.C.; Chen, C.C.; Zhang, W.D.; Zhang, S.K.; Chang, F.H.; Hsu, L.S. The 252A/G and 804C/A polymorphisms of Lymphotoxin-alpha is associated to onset of acute myocardial infarction in Taiwan. LabMedicine 2010, 41, 220–225. [Google Scholar]
- Govan, V.A.; Constant, D.; Hoffman, M.; Williamson, A.L. The allelic distribution of -308 tumor necrosis factor-alpha gene polymorphism in South African women with cervical cancer and control women. BMC Cancer 2006, 6, 24. [Google Scholar]
- Cambien, F.; Poirier, O.; Mallet, C.; Tiret, L. Coronary heart disease and genetics in epidemiologist’s view. Mol. Med. Today 1997, 3, 197–203. [Google Scholar]
- Incalcaterra, E.; Hoffmann, E.; Averna, M.R.; Caimi, G. Genetic risk factors in myocardial infarction at young age. Minerva Cardioangiol 2004, 52, 287–312. [Google Scholar]
- Ritchie, S.A.; Connell, J.M. The link between abdominal obesity, metabolic syndrome and cardiovascular disease. Nutr. Metab. Cardiovasc. Dis 2007, 17, 319–326. [Google Scholar]
- Elkind, M.S. Inflammation, atherosclerosis, and stroke. Neurologist 2006, 12, 140–148. [Google Scholar]
- Chen, D.; Assad-Kottner, C.; Orrego, C.; Torre-Amione, G. Cytokines and acute heart failure. Crit. Care Med 2008, 36, S9–S16. [Google Scholar]
- Abraham, L.J.; Kroeger, K.M. Impact of the -308 TNF promoter polymorphism on the transcriptional regulation of the TNF gene: relevance to disease. J. Leukoc. Biol 1999, 66, 562–566. [Google Scholar]
- Padovani, J.C.; Pazin-Filho, A.; Simoes, M.V.; Marin-Neto, J.A.; Zago, M.A.; Franco, R.F. Gene polymorphisms in the TNF locus and the risk of myocardial infarction. Thromb. Res 2000, 100, 263–269. [Google Scholar]
- Koch, W.; Kastrati, A.; Bottiger, C.; Mehilli, J.; von Beckerath, N.; Schomig, A. Interleukin-10 and tumor necrosis factor gene polymorphisms and risk of coronary artery disease and myocardial infarction. Atherosclerosis 2001, 159, 137–144. [Google Scholar]
- Antonicelli, R.; Olivieri, F.; Bonafe, M.; Cavallone, L.; Spazzafumo, L.; Marchegiani, F.; Cardelli, M.; Recanatini, A.; Testarmata, P.; Boemi, M.; et al. The interleukin-6 -174 G > C promoter polymorphism is associated with a higher risk of death after an acute coronary syndrome in male elderly patients. Int. J. Cardiol 2005, 103, 266–271. [Google Scholar]
- Sheu, W.H.; Lee, W.J.; Lin, L.Y.; Chang, R.L.; Chen, Y.T. Tumor necrosis factor alpha -238 and -308 polymorphisms do not associate with insulin resistance in hypertensive subjects. Metabolism 2001, 50, 1447–1451. [Google Scholar]
- Shiau, M.Y.; Wu, C.Y.; Huang, C.N.; Hu, S.W.; Lin, S.J.; Chang, Y.H. TNF-alpha polymorphisms and type 2 diabetes mellitus in Taiwanese patients. Tissue Antigens 2003, 61, 393–397. [Google Scholar]
- Benjafield, A.V.; Lin, R.C.; Dalziel, B.; Gosby, A.K.; Caterson, I.D.; Morris, B.J. G-protein beta3 subunit gene splice variant in obesity and overweight. Int. J. Obes. Relat. Metab. Disord 2001, 25, 777–780. [Google Scholar]
- Stefan, N.; Stumvoll, M.; Machicao, F.; Koch, M.; Haring, H.U.; Fritsche, A. C825T polymorphism of the G protein beta3 subunit is associated with obesity but not with insulin sensitivity. Obes. Res 2004, 12, 679–683. [Google Scholar]
- Yamamoto, M.; Abe, M.; Jin, J.J.; Wu, Z.; Tabara, Y.; Mogi, M.; Kohara, K.; Miki, T.; Nakura, J. Association of GNB3 gene with pulse pressure and clustering of risk factors for cardiovascular disease in Japanese. Biochem. Biophys. Res. Commun 2004, 316, 744–748. [Google Scholar]
- Naber, C.K.; Husing, J.; Wolfhard, U.; Erbel, R.; Siffert, W. Interaction of the ACE D allele and the GNB3 825T allele in myocardial infarction. Hypertension 2000, 36, 986–989. [Google Scholar]
- Hanon, O.; Luong, V.; Mourad, J.J.; Bortolotto, L.A.; Safar, M.; Girerd, X. Association between the G protein beta3 subunit 825t allele and radial artery hypertrophy. J. Vasc. Res 2002, 39, 497–503. [Google Scholar]
- Kiani, J.G.; Saeed, M.; Parvez, S.H.; Frossard, P.M. Association of G-protein beta-3 subunit gene (GNB3) T825 allele with Type II diabetes. Neuro. Endocrinol. Lett 2005, 26, 87–88. [Google Scholar]
- Hengstenberg, C.; Schunkert, H.; Mayer, B.; Doring, A.; Lowel, H.; Hense, H.W.; Fischer, M.; Riegger, G.A.; Holmer, S.R. Association between a polymorphism in the G protein beta3 subunit gene (GNB3) with arterial hypertension but not with myocardial infarction. Cardiovasc. Res 2001, 49, 820–827. [Google Scholar]
- Klintschar, M.; Stiller, D.; Schwaiger, P.; Kleiber, M. DNA polymorphisms in the tyrosine hydroxylase and GNB3 genes: Association with unexpected death from acute myocardial infarction and increased heart weight. Forensic. Sci. Int 2005, 153, 142–146. [Google Scholar]



{kind=link}
{kind=link}
| TGF -308 | G/A | p value | Spearman Colleration | O.R. (95%C.I.) |
|---|---|---|---|---|
| AA vs. GG+GA | ||||
| BMI >= 27 | AMI/IIA | |||
| GG 65%/54% | 0.063 | 0.323 (0.013) | ND | |
| GA 20%/46% | ||||
| AA 15%/0% | ||||
| BMI < 27 | GG 72%/69% | 1.0 | 0.013 (0.087) | 1.124 (0.260–4.855) |
| GA 23%/27% | ||||
| AA 5%/4% | ||||
| GNB C825T | ||||
| TT vs. CC+CT | ||||
| BMI >= 27 | AMI/IIA | |||
| CC 10%/5% | 0.009 ** | −0.381 (0.003) | 0.548 (0.331–0.910) | |
| CT 45%/13% | ||||
| TT 45%/82% | ||||
| BMI < 27 | ||||
| CC 13%/21% | 0.908 | 0.022 (0.783) | 1.048 (0.753–1.048) | |
| CT 37%/31% | ||||
| TT 50%/48% | ||||
| p value | Spearman Correlation | O.R. (95%C.I.) | ||
|---|---|---|---|---|
| BMI >= 27, female | ||||
| AMI/IIA | ||||
| TT | 0%/89% | 0.032 * | −0.667 (0.001) | ND |
| CC+CT | 100%/11% | |||
| BMI >= 27, male | ||||
| AMI/IIA | ||||
| TT | 50%/76% | 0.108 | −0.272 (0.094) | 0.658 (0.390–1.104) |
| CC+CT | 50%/24% | |||
| p value | Spearman Correlation | O.R. (95%C.I.) | ||
|---|---|---|---|---|
| BMI > 27/BMI < 27 | ||||
| TT | 70%/48% | 0.006 ** | 0.19 (0.006) ** | 1.437 (1.135–1.181) |
| CT+CC | 30%/52% |
| p value | Spearman Correlation | O.R. (95%C.I.) | ||
|---|---|---|---|---|
| AA vs. GG+GA | ||||
| Male | AMI/IIA+IIB | |||
| AA | 7%/100% | 0.026 * | 0.2 (0.008) | ND |
| GA+GG | 93%/0% | |||
| Female | ||||
| AA | 5%/6% | 1.00 | −0.016 (0.879) | 0.845 (0.100–7.159) |
| GA+GG | 95%/94% | |||
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chang, W.-T.; Wang, Y.-C.; Chen, C.-C.; Zhang, S.-K.; Liu, C.-H.; Chang, F.-H.; Hsu, L.-S. The -308G/A of Tumor Necrosis Factor (TNF)-α and 825C/T of Guanidine Nucleotide Binding Protein 3 (GNB3) are Associated with the Onset of Acute Myocardial Infarction and Obesity in Taiwan. Int. J. Mol. Sci. 2012, 13, 1846-1857. https://doi.org/10.3390/ijms13021846
Chang W-T, Wang Y-C, Chen C-C, Zhang S-K, Liu C-H, Chang F-H, Hsu L-S. The -308G/A of Tumor Necrosis Factor (TNF)-α and 825C/T of Guanidine Nucleotide Binding Protein 3 (GNB3) are Associated with the Onset of Acute Myocardial Infarction and Obesity in Taiwan. International Journal of Molecular Sciences. 2012; 13(2):1846-1857. https://doi.org/10.3390/ijms13021846
Chicago/Turabian StyleChang, Wei-To, Yi-Cheng Wang, Chun-Chang Chen, Shi-Kun Zhang, Chen-Hsun Liu, Fu-Hsin Chang, and Li-Sung Hsu. 2012. "The -308G/A of Tumor Necrosis Factor (TNF)-α and 825C/T of Guanidine Nucleotide Binding Protein 3 (GNB3) are Associated with the Onset of Acute Myocardial Infarction and Obesity in Taiwan" International Journal of Molecular Sciences 13, no. 2: 1846-1857. https://doi.org/10.3390/ijms13021846
APA StyleChang, W.-T., Wang, Y.-C., Chen, C.-C., Zhang, S.-K., Liu, C.-H., Chang, F.-H., & Hsu, L.-S. (2012). The -308G/A of Tumor Necrosis Factor (TNF)-α and 825C/T of Guanidine Nucleotide Binding Protein 3 (GNB3) are Associated with the Onset of Acute Myocardial Infarction and Obesity in Taiwan. International Journal of Molecular Sciences, 13(2), 1846-1857. https://doi.org/10.3390/ijms13021846